
Combining Ability and Gene Action for Yield Characteristics in Novel Aromatic Cytoplasmic Male Sterile Hybrid Rice under Water-Stress Conditions

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genotypes Materials and Experimental Designation
2.2. Data Analysis
3. Results
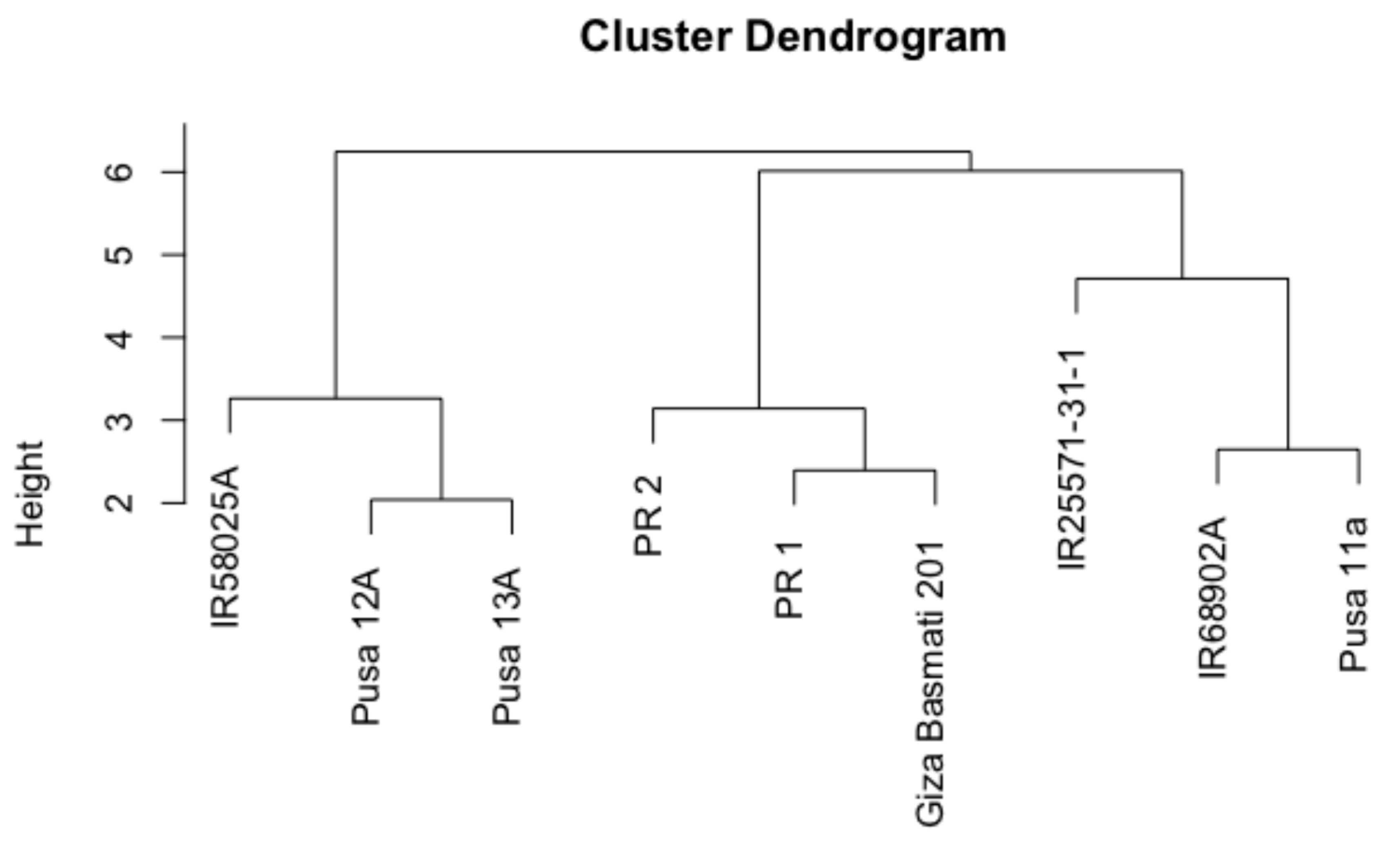
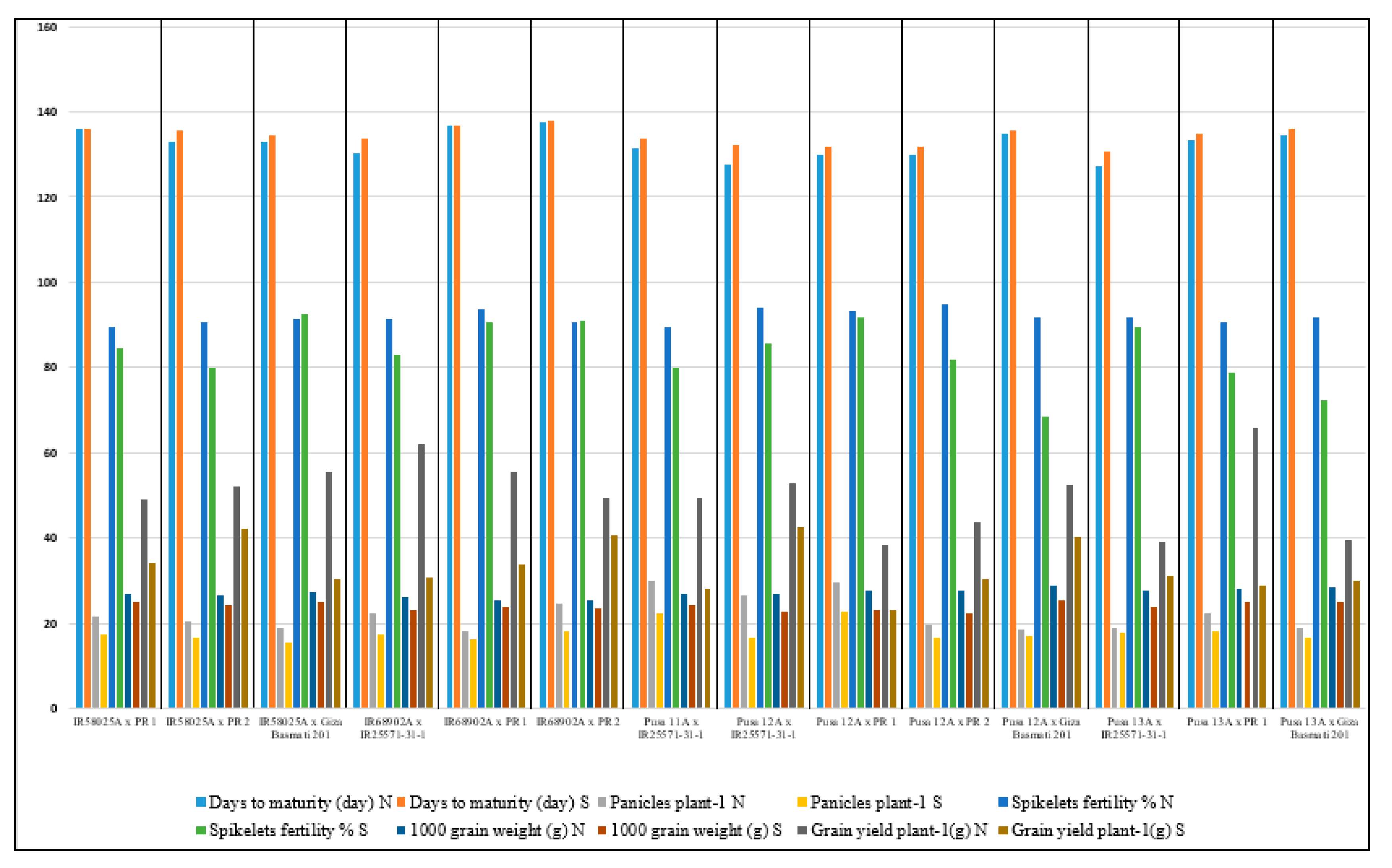
3.1. Genotypes Performance and Variation
3.2. Analysis of Variance (ANOVA)
3.3. Estimation of the Genetic Parameters and Gene Action
3.4. General Combining Ability Estimation
3.5. Specific Combining Ability Estimation
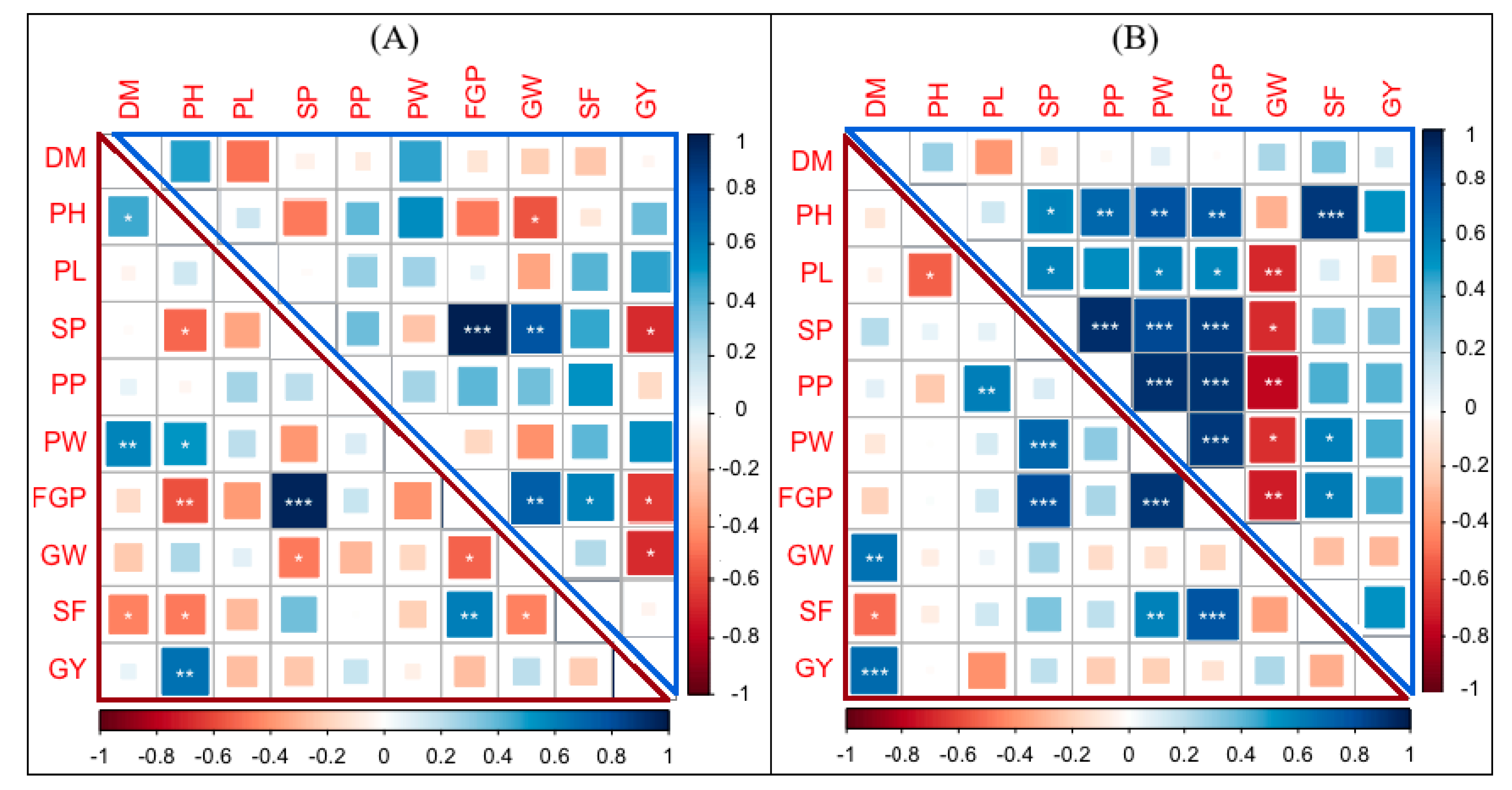
3.6. Correlation and Distribution of Combining Ability of Yield Traits
4. Discussion
4.1. Variation Analysis
4.2. Genetic Components of Variance
4.3. Combining Ability and Gene Action
4.4. Correlation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khush, G.S. What it will take to Feed 5.0 Billion Rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.H.; Zhuang, J.I.Y.; Fan, Y.Y.; Du, J.I.H.; Cao, L.I.Y. Progress in research and development on hybrid rice: A super-domesticate in China. Ann. Bot. 2007, 100, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Leonilo, V.G.; Joanne, D.C.; Enriquez, J.O.S. Heterosis and combining ability analysis in CMS hybrid rice. Euphytica 2020, 216, 14. [Google Scholar]
- Khush, G. Strategies for increasing the yield potential of cereals: Case of rice as an example. Plant. Breed. 2013, 132, 433–436. [Google Scholar] [CrossRef]
- El-Mowafi, H.F.; Bastawisi, A.O.; Abo-Youssef, M.I.; Zaman, F.U. Exploitation of rice heterosis under Egyptian conditions. Egypt. J. Agric. 2005, 83, 143–166. [Google Scholar]
- Shull, G.H. Duplicate genes for capsule form in Bursa bursa-pastoris. Zeitschrift Induktive Abstammungs Vererbungslehre 1914, 12, 97–149. [Google Scholar] [CrossRef]
- Virmani, S.S. Hybrid rice. Adv. Agron. 1996, 57, 378–462. [Google Scholar]
- Virmani, S.S.; Mao, C.X.; Hardy, B. Hybrid Rice for Food Security, Poverty Alleviation, and Environmental Protection. In Proceedings of the 4th International Symposium on Hybrid Rice, Hanoi, Vietnam, 14–17 May 2002; p. 407. [Google Scholar]
- Virmani, S.S.; Sun, Z.X.; Mou, T.M.; Ali, A.J.; Mao, C.X. Two Lines Hybrid Rice Breeding Manual; International Rice Research Institute: Los Banos, Philippines, 1997. [Google Scholar]
- Chang, Z.; Wang, N.; Xie, G.; Lu, W.; Zhou, J.; Tang, X.; Deng, X.W. Construction of a male sterility system for hybrid rice breeding and seed production using a nuclear male sterility gene. Proc. Natl. Acad. Sci. USA 2016, 113, 14145–14150. [Google Scholar] [CrossRef] [Green Version]
- Sprague, G.A.; Tatum, L.A. General vs specific combining ability in single crosses of corn. J. Am. Soc. Agron. 1942, 34, 923–932. [Google Scholar] [CrossRef]
- Griffing, B. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Aly, R.S.H. Relationship between combining ability of grain yield and yield components for some newly yellow maize inbred lines via line 9 tester analysis. Alex. J. Agric. Res. 2013, 58, 115–124. [Google Scholar]
- Latha, S.; Sharma, D.; Sanghera, G.S. Combining ability and heterosis for grain yield and its components in rice (Oryza sativa L.). Not. Sci. Biol. 2013, 5, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Zhang, F.; Yang, X.; Feng, Y.; Yang, X.; Wu, Y.; Guan, Z.; Fang, W.; Chen, F. Combining ability, heterosis, genetic distance and their intercorrelations for waterlogging tolerance traits in chrysanthemum. Euphytica 2017, 213, 42. [Google Scholar] [CrossRef] [Green Version]
- Kempthorne, O. An Introduction to Genetic Studies; John Willey Sons Inc.: New York, NY, USA, 1957; pp. 458–471. [Google Scholar]
- Dey, S.S.; Bhatia, R.; Bhardwaj, I.; Mishra, V.; Sharma, K.; Parkash, C. Molecular-agronomic characterization and genetic study reveals usefulness of refined Ogura cytoplasm based CMS lines in hybrid breeding of cauliflower (Brassica oleracea var. botrytis L.). Sci. Hortic. 2017, 224, 27–36. [Google Scholar] [CrossRef]
- Wang, K.; Qiu, F.; Larazo, W.; Dela Paz, M.A.; Xie, F. Heterotic groups in tropical indica rice germplasm. Appl. Genet. 2015, 128, 421–430. [Google Scholar] [CrossRef]
- Wang, Y.; Cai, Q.; Xie, H.; Wu, F.; Lian, L.; He, W.; Chen, L.; Xie, H.; Zhang, J. Determination of heterotic groups and heterosis analysis of yield performance in indica rice. Rice Sci. 2018, 25, 261–269. [Google Scholar]
- Xie, F.; He, Z.; Esguerra, M.Q.; Qiu, F.; Ramanathan, V. Determination of heterotic groups for tropical Indica hybrid rice germplasm. Appl. Genet. 2014, 127, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.K.; Kumar, A. Combining ability analysis to identify suitable parents for heterotic rice hybrid breeding. IRRN 2004, 29, 21–22. [Google Scholar]
- El-Mowafi, H.F.; Abdallah, R.M.; Abdelkhalek, S.M.; Mubarak, M.H. Combining ability and molecular evaluation for developing new basmati rice hybrids in Egypt. Egypt J. Plant Breed. 2018, 22, 1467–1485. [Google Scholar]
- International Rice Research Institute (IRRI). Standard Evaluation System (SES) for Rice, 5th ed.; International Rice Research Institute (IRRI): Los Banos, Philippines, 2013.
- Esteves, F.R. Script for UPGMA Analysis. Available online: https://www.researchgate.net/publication/303944638_R_script_for_Principal_Component_Analysis_PCA (accessed on 15 July 2018).
- Team, C.R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008. [Google Scholar]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Corrplot: Visualization of a Correlation Matrix. Available online: http://cran.r-project.org/package=corrplot (accessed on 2 August 2018).
- Peterson, B.G.; Carl, P.; Boudt, K.; Bennett, R.; Ulrich, J.; Zivot, E.; Cornilly, D.; Hung, E.; Lestel, M.; Balkissoon, K. Performance Analytics: Econometric Tools for Performance and Risk Analysis. Available online: https://github.com/braverock/PerformanceAnalytics (accessed on 2 August 2018).
- Griffiths, A.J.F.; Miller, J.H.; Suzuki, D.T.; Lewontin, R.C.; Gelbart, W.M. An Introduction to Genetic Analysis, 7th ed.; WH Freeman and Company: New York, NY, USA, 2000; p. 860. [Google Scholar]
- Verma, O.P.; Srivastava, H.K. Genetic components and combining ability analysis in relation to heterosis for yield and associated traits using three divers rice growing ecosystem. Proc. Natl. Acad. Sci. USA 2004, 88, 91–102. [Google Scholar]
- Manickavelu, A.; Nadarajan, S.K.; Ganesh, R.P.; Gnanamalar, R.C. Drought tolerance in rice: Morphological and molecular genetic consideration. Plant. Growth Regul. 2006, 50, 121–138. [Google Scholar] [CrossRef]
- El-Hity, M.A.; El-Degwy, E.S.; Reda, A.M.; Abdallah, R.M.; Hadefa, E.A. Combining ability and genetic parameters analysis for three and two line system in hybrid rice under normal and water-stress conditions. J. Agric. Res. 2015, 41, 1118–1131. [Google Scholar]
- Herwibawa, B.; Sakhidin; Haryanto, T.A. Agronomic performances of aromatic and nonaromatic M1 rice under drought stress. Open Agric. 2019, 4, 575–584. [Google Scholar] [CrossRef]
- Kropff, M.J.; Cassman, K.G.; Peng, S.; Matthews, R.B.; Setter, T.L. Quantitative understanding of yield potential. Break. Yield Barrier 1994, 1, 21–38. [Google Scholar]
- Ying, J.; Peng, S.; He, Q.; Yang, H.; Yang, C.; Visperas, R.M.; Cassman, K.G. Comparison of high yield rice in tropical and subtropical environments I: Determinants of grain and dry matter yields. Field Crop. Res. 1998, 57, 71–84. [Google Scholar] [CrossRef]
- Singh, R.K.; Chaudhary, B.D. Biometrical methods in quantitative genetic analysis. Biom. Methods Quant. Genet. Anal. 1979, 3, 318. [Google Scholar]
- Sharma, M.K.; Sharma, A.K.; Agarwal, R.K.; Richharia, A.K. Combining ability and gene action for yield and quality characteristics in Ahu rices of Assam. Indian J. Genet. 2007, 67, 278–280. [Google Scholar]
- Tyagi, J.P.; Tejbir, S.; Singh, V.P. Genetic analysis of combining ability for quality characteristics in Basmati rice. Int. J. Rice 2010, 47, 96–99. [Google Scholar]
- Qu, Z.; Li, L.; Luo, J.; Wang, P.; Yu, S.; Mou, T.; Zheng, X.; Hu, Z. QTL mapping of combining ability and heterosis of agronomic traits in rice backcross recombinant inbred lines and hybrid crosses. PLoS ONE 2012, 7, 1–10. [Google Scholar] [CrossRef]
- Ganapati, R.K.; Rasul, M.G.; Sarker, U.; Singha, A.; Faruquee, M. Gene action of yield and yield contributing traits of submergence tolerant rice (Oryza sativa L.) in Bangladesh. Bull. Natl. Res. Cent. 2020, 44, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A. Combining ability for yield and its related traits in upland rice. Int. J. Rice 1993, 30, 275–279. [Google Scholar]
- Sreeramachandra, B.M.; Satyanarana, P.V.; Madhuri, J.; Kumar, R.V. Combining ability analysis for identifying elite parents for hybrid rice. Acad. J. Agric. Res. 2000, 37, 19–22. [Google Scholar]
- Singh, R.K. Heterosis breeding in aromatic rice for yield and quality characteristics. Indian J. Genet. Plant Breed. 2005, 65, 176–179. [Google Scholar]
- Kumar, S.; Singh, H.B.; Sharma, J.K. Combining ability analysis for grain yield and other associated traits in rice. Oryza 2007, 44, 108–114. [Google Scholar]
- El-Mowafi, H.F.; Reda, A.M.; Abdallah, R.M.; Arafat, E.F.A. Combining ability analysis for agronomic and yield attributing traits in hybrid rice. Egypt J. Plant Breed. 2015, 19, 2195–2219. [Google Scholar] [CrossRef]
- Huang, M.; Chen, L.Y.; Chen, Z.Q. Diallel analysis of combining ability and heterosis for yield and yield components in rice by using positive loci. Euphytica 2015, 205, 37–50. [Google Scholar] [CrossRef]
- Tiwari, K.; Pandey, P.; Giri, S.P.; Dwivedi, J.L. Heterosis studies for yield and its components in rice hybrids using CMS system. Asian J. Plant. Sci. 2011, 10, 29–42. [Google Scholar] [CrossRef]
- Yuga, M.E.; Kimani, P.M.; Olubayo, M.F.; Muthomi, J.W.; Nzuve, F.M. Combining ability of heterosis for agronomic and yield traits in indica and japonica rice crosses. J. Agric. Sci. 2018, 10, 92–103. [Google Scholar] [CrossRef]
- Attia, K.A.; Zhong, X.Q.; Bastawisi, A.O. Combining ability and standard heterosis analysis of two-line system hybrid rice. Pak. J. Biol. Sci. 2001, 4, 346–350. [Google Scholar]
- Singh, S.; Bhatia, R.; Kumar, R.; Sharma, K.; Dash, S.; Dey, S.S. Cytoplasmic male sterile and doubled haploid lines with desirable combining ability enhances the concentration of important antioxidant attributes in Brassica oleracea. Euphytica 2018, 214, 207. [Google Scholar] [CrossRef]
- Fasahat, P.; Rajabi, A.; Rad, M.J.; Derera, J. Principles and utilization of combining ability in plant breeding. Biom. Biostat. Int. J. 2016, 4, 1–24. [Google Scholar] [CrossRef]
- Singh, S.; Dey, S.S.; Bhatia, R.; Kumar, R.; Sharma, K.; Behera, T.K. Heterosis and combining ability in cytoplasmic male sterile and doubled haploid based Brassica oleracea progenies and prediction of heterosis using microsatellites. PLoS ONE 2019, 14, 1–26. [Google Scholar] [CrossRef]
- Zaid, I.U.; Tang, W.; Liu, E.; Khan, S.U.; Wang, H.; Mawuli, E.W.; Hong, D. Genome-Wide single-nucleotide polymorphisms in cms and restorer lines discovered by genotyping using sequencing and association with marker-combining ability for 12 yield-related traits in Oryza sativa L. subsp. Japonica. Front. Plant Sci. 2019, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhou, H.; Xie, W.; Xia, D.; Gao, G.; Zhang, Q.; Wang, G.; Lian, X.; Xiao, J.; He, Y.; et al. Genome-wide association analyses reveal the genetic basis of combining ability in rice. Plant Biot. J. 2019, 17, 2211–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, V.; Dhillon, S.K.; Kaushik, P.; Kaur, G. Characterization for Drought tolerance and physiological efficiency in novel cytoplasmic male sterile sources of sunflower (Helianthus annuus L.). Agronomy 2018, 8, 232. [Google Scholar] [CrossRef] [Green Version]
- Sanghera, G.S.; Hussain, W. Heterosis and combining ability estimates using line × tester analysis to develop rice hybrids for temprate conditions. Not. Sci. Biol. 2012, 4, 131–142. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.O.V. | df | Days to Maturity (Day) | Plant Height (cm) | Panicle Length (cm) | Spikelets/Panicle | Tillers Plant | |||||
| N | S | N | S | N | S | N | S | N | S | ||
| Replications | 2 | 0.39 | 0.55 | 6.02 | 5.38 | 0.04 | 0.01 | 13.35 | 111.86 | 2.69 | 0.01 |
| Treatments | 28 | 34.91 ** | 22.79 ** | 170.58 ** | 178.57 ** | 8.77 ** | 9.28 ** | 1855.11 ** | 2504.73 ** | 37.21 ** | 18.02 ** |
| Parents | 8 | 44.62 ** | 52.02 ** | 135.76 ** | 95.75 ** | 14.73 ** | 8.06 ** | 458.96 ** | 713.41 ** | 24.22 ** | 26.65 ** |
| Crosses | 19 | 26.93 ** | 10.68 ** | 63.19 ** | 152.45 ** | 4.60 ** | 8.03 ** | 697.40 ** | 1707.76 ** | 39.71 ** | 13.82 ** |
| Par.vs.crosses | 1 | 108.70 ** | 18.94 ** | 2489.36 ** | 1337.44 ** | 40.26 ** | 42.99 ** | 35,020.86 ** | 31,977.68 ** | 93.47 ** | 28.92 ** |
| Lines | 4 | 35.19 ** | 18.17 ** | 127.74 ** | 383.48 ** | 2.65 ** | 26.84 ** | 1900.03 ** | 3098.91 ** | 28.72 ** | 13.82 ** |
| Testers | 3 | 87.54 ** | 25.84 ** | 27.15 * | 57.65 ** | 3.29 * | 3.15 ** | 644.52 ** | 1151.53 ** | 56.25 ** | 12.40 ** |
| Lines × testers | 12 | 9.03 ** | 4.40 * | 50.69 ** | 99.14 ** | 5.58 ** | 2.97 ** | 309.75 ** | 1383.10 ** | 39.24 ** | 14.18 ** |
| Residual | 56 | 0.52 | 1.79 | 11.23 | 2.12 | 0.94 | 0.01 | 28.50 | 122.01 | 0.96 | 0.02 |
| CV% | 0.54 | 0.99 | 2.95 | 1.43 | 3.48 | 0.47 | 2.34 | 6.03 | 4.61 | 0.7 | |
| S.O.V. | df | Panicle Weight | Filled Grains Panicle | Grain Yield Plant | Spikelet Fertility% | 1000 Grain Weight | |||||
| N | S | N | S | N | S | N | S | N | S | ||
| Replications | 2 | 0.03 | 0.01 | 7.74 | 0.86 | 2.45 | 0.05 | 0.22 | 4.56 | 0.05 | 0.002 |
| Treatments | 28 | 2.13 ** | 2.41 ** | 1689.51 ** | 2482.01 ** | 212.35 ** | 93.58 ** | 4.79 ** | 123.23 ** | 8.47 ** | 13.70 ** |
| Parents | 8 | 1.30 ** | 0.59 ** | 373.21 ** | 570.26 ** | 53.36 ** | 57.87 ** | 1.57 | 177.96 ** | 20.87 ** | 32.76 ** |
| Crosses | 19 | 0.26 ** | 1.96 ** | 743.52 ** | 2005.10 ** | 209.98 ** | 84.40 ** | 6.36 ** | 101.85 ** | 2.96 ** | 2.66 ** |
| Par.vs.crosses | 1 | 44.29 ** | 25.46 ** | 301,93.81 ** | 26,837.20 ** | 1529.30 ** | 553.66 ** | 0.55 | 91.64 ** | 13.89 ** | 70.79 ** |
| Lines | 4 | 0.37 ** | 5.21 ** | 2001.32 ** | 2737.07 ** | 103.81 ** | 74.50 ** | 13.16 ** | 143.07 ** | 9.62 ** | 5.50 ** |
| Testers | 3 | 0.21 | 0.04 | 599.43 ** | 1431.12 ** | 243.62 ** | 69.14 ** | 0.80 | 23.04 ** | 3.76 ** | 4.19 ** |
| Lines × testers | 12 | 0.23 ** | 1.35 ** | 360.27 ** | 1904.61 ** | 236.96 ** | 91.52 | 5.49 ** | 107.81 ** | 0.54 ** | 1.33 ** |
| Residual | 56 | 0.08 | 0.02 | 36.97 | 0.67 | 6.37 | 0.53 | 1.71 | 1.90 | 0.07 | 0.04 |
| CV% | 5.30 | 3.49 | 2.91 | 0.54 | 5.49 | 2.37 | 1.42 | 1.66 | 0.99 | 0.80 | |
| S.O.V. | Days to Maturity (Day) | Plant Height (cm) | Panicle Length (cm) | Spikelets Panicle−1 | Panicles Plant−1 | |||||
| N | S | N | S | N | S | N | S | N | S | |
| Parameter | ||||||||||
| Additive variance (σ2 A) | 7.414 | 2.577 | 4.852 | 20.928 | −0.397 | 2.001 | 154.842 | 127.546 | 0.257 | −0.149 |
| Dominant variance (σ2 D) | 2.835 | 0.869 | 13.153 | 32.341 | 1.545 | 0.986 | 93.747 | 420.363 | 12.763 | 4.720 |
| Environmental variance (σ2 E) | 0.522 | 1.794 | 11.227 | 2.120 | 0.942 | 0.014 | 28.504 | 122.013 | 0.955 | 0.016 |
| Genotypic variance (σ2 G) | 10.249 | 3.445 | 18.006 | 53.269 | 1.148 | 2.987 | 248.589 | 547.909 | 13.020 | 4.571 |
| Phenotypic variance (σ2 P) | 10.771 | 5.239 | 29.233 | 55.389 | 2.090 | 3.001 | 277.093 | 669.922 | 13.975 | 4.587 |
| Broad sense heritability (h2b)% | 95.154 | 65.758 | 61.595 | 96.173 | 54.930 | 99.533 | 89.713 | 81.787 | 93.166 | 99.651 |
| Narrow sense heritability (h2n)% | 68.833 | 49.178 | 16.599 | 37.784 | −18.975 | 66.689 | 55.881 | 19.039 | 1.839 | −3.242 |
| Relative importance of GCA% * | 72.339 | 74.786 | 26.950 | 39.288 | −34.545 | 67.002 | 62.288 | 23.279 | 1.973 | −3.254 |
| Relative importance of SCA% ** | 27.661 | 25.214 | 73.050 | 60.712 | 134.545 | 32.998 | 37.712 | 76.721 | 98.027 | 103.254 |
| S.O.V. | Panicle Weight (g) | Filled Grains Panicle−1 | 1000 Grain Weight (g) | Spikelet Fertility% | Grain Yield Plant−1 | |||||
| N | S | N | S | N | S | N | S | N | S | |
| Parameter | ||||||||||
| Additive variance (σ2 A) | 0.010 | 0.235 | 152.698 | 37.805 | 0.971 | 0.537 | 0.326 | −2.713 | −10.652 | −2.909 |
| Dominant variance (σ2 D) | 0.050 | 0.444 | 107.768 | 634.649 | 0.156 | 0.433 | 1.260 | 35.302 | 76.866 | 30.328 |
| Environmental variance (σ2 E) | 0.084 | 0.019 | 36.965 | 0.666 | 0.070 | 0.035 | 1.708 | 1.902 | 6.366 | 0.530 |
| Genotypic variance (σ2 G) | 0.059 | 0.679 | 260.467 | 672.454 | 1.128 | 0.970 | 1.586 | 32.589 | 66.214 | 27.419 |
| Phenotypic variance (σ2 P) | 0.143 | 0.698 | 297.432 | 673.120 | 1.198 | 1.005 | 3.294 | 34.491 | 72.580 | 27.949 |
| Broad sense heritability (h2b)% | 41.429 | 97.276 | 87.572 | 99.901 | 94.155 | 96.519 | 48.155 | 94.485 | 91.229 | 98.104 |
| Narrow sense heritability (h2n)% | 6.798 | 33.677 | 51.339 | 5.616 | 81.101 | 53.454 | 9.910 | −7.866 | −14.677 | −10.410 |
| Relative importance of GCA% * | 16.410 | 34.620 | 58.625 | 5.622 | 86.136 | 55.382 | 20.578 | −8.325 | −16.088 | −10.611 |
| Relative importance of SCA% ** | 83.590 | 65.380 | 41.375 | 94.378 | 13.864 | 44.618 | 79.422 | 108.325 | 116.088 | 110.611 |
| Genotypes | Days to Maturity (day) | Plant Height (cm) | Panicle Length (cm) | Spikelets Panicle−1 | Panicles Plant−1 | |||||
| N | S | N | S | N | S | N | S | N | S | |
| Lines | ||||||||||
| IR58025A | 0.35 * | 0.21 | 2.79 ** | 2.44 ** | −0.96 ** | −0.33 ** | −15.70 ** | −1.29 | −0.08 | −0.28 ** |
| IR68902A | 2.03 ** | 1.83 ** | 2.71 ** | 4.69 ** | 0.51 * | −0.47 ** | −11.04 ** | −4.02 | −0.27 | 0.24 ** |
| Pusa 11a | 0.93 ** | 0.03 | 1.29 | −2.31 ** | 0.98 ** | 1.69 ** | 8.54 ** | 9.29 ** | 0.14 | 0.79 ** |
| Pusa 12A | −2.42 ** | −1.55 ** | −2.21 * | 4.03 ** | 1.64 ** | 0.26 ** | 12.75 ** | 19.47 ** | 0.72 ** | 1.61 ** |
| Pusa 13A | −0.89 ** | −0.52 | −4.57 ** | −8.85 ** | −2.18 ** | −1.16 ** | 5.45 ** | −23.45 ** | −0.51 * | −2.36 ** |
| LSD 5% | 0.35 | 0.65 | 1.62 | 0.70 | 0.47 | 0.06 | 2.58 | 5.33 | 0.47 | 0.06 |
| 1% | 0.50 | 0.92 | 2.31 | 1.00 | 0.67 | 0.09 | 3.68 | 7.62 | 0.67 | 0.08 |
| Testers | ||||||||||
| IR25571-31-1 | −3.48 ** | −1.86 ** | 0.22 | −1.14 ** | 2.35 ** | 0.95 ** | −9.44 ** | −4.59 | 0.09 | −0.33 ** |
| PR 1 | 0.92 ** | 0.12 | −1.71 * | 2.13 ** | 0.65 ** | 0.57 ** | 0.82 | 8.68 ** | −0.46 * | 0.31 ** |
| PR 2 | 0.48 ** | 0.58 * | −0.07 | −2.11 ** | −0.87 ** | −0.53 ** | 5.30 ** | 5.88 ** | −0.24 | 0.47 ** |
| Giza Basmati 201 | 2.08 ** | 1.16 ** | 1.57 * | 1.13 ** | −2.13 ** | −0.98 ** | 3.32 ** | −9.97 ** | 0.61 ** | −0.45 ** |
| LSD 5% | 0.31 | 0.58 | 1.45 | 0.63 | 0.42 | 0.05 | 2.30 | 4.77 | 0.42 | 0.05 |
| 1% | 0.45 | 0.83 | 2.07 | 0.90 | 0.60 | 0.08 | 3.29 | 6.82 | 0.60 | 0.07 |
| Genotypes | Panicle Weight (g) | Filled Grains Panicle−1 | 1000 Grain Weight (g) | Spikelet Fertility% | Grain Yield Plant−1 (g) | |||||
| N | S | N | S | N | S | N | S | N | S | |
| Lines | ||||||||||
| IR58025A | −0.02 | 0.16 ** | −16.57 ** | 3.63 ** | −0.54 ** | 0.58 ** | −0.95 ** | 1.58 ** | 3.89 ** | 2.29 ** |
| IR68902A | 0.30 ** | 0.18 ** | −9.44 ** | 5.92 ** | −1.21 ** | −0.32 ** | 0.34 | 4.53 ** | 2.38 ** | 1.55 ** |
| Pusa 11a | −0.09 | 0.41 ** | 5.86 ** | 4.75 ** | 0.07 | −0.60 ** | −0.79 * | −2.01 ** | −1.51 * | −2.50 ** |
| Pusa 12A | −0.03 | 0.41 ** | 15.82 ** | 12.08 ** | 0.72 ** | −0.52 ** | 1.64 ** | 0.42 | −2.01 ** | 1.57 ** |
| Pusa 13A | −0.15 * | −1.16 ** | 4.33 ** | −26.38 ** | 0.96 ** | 0.87 ** | −0.24 | −4.52 ** | −2.74 ** | −2.91 ** |
| LSD 5% | 0.14 | 0.07 | 2.93 | 0.39 | 0.13 | 0.09 | 0.63 | 0.67 | 1.22 | 0.35 |
| 1% | 0.20 | 0.10 | 4.19 | 0.56 | 0.18 | 0.13 | 0.90 | 0.95 | 1.74 | 0.50 |
| Testers | ||||||||||
| IR25571-31-1 | −0.06 | 0.01 | −9.02 ** | −1.34 ** | −0.43 ** | −0.38 ** | −0.12 | 1.06 ** | 2.73 ** | 0.42 * |
| PR 1 | 0.15 * | 0.07 * | 1.05 | 10.99 ** | −0.13 * | 0.06 | 0.09 | 0.88 ** | 4.13 ** | −2.34 ** |
| PR 2 | −0.13 * | −0.01 | 5.68 ** | 2.83 ** | −0.16 ** | −0.40 ** | 0.28 | −1.62 ** | −2.83 ** | 2.74 ** |
| Giza Basmati 201 | 0.04 | −0.07 * | 2.29 | −12.48 ** | 0.72 ** | 0.73 ** | −0.24 | −0.32 | −4.03 ** | −0.82 ** |
| LSD 5% | 0.13 | 0.06 | 2.62 | 0.35 | 0.11 | 0.08 | 0.56 | 0.60 | 1.09 | 0.31 |
| 1% | 0.18 | 0.09 | 3.75 | 0.50 | 0.16 | 0.12 | 0.81 | 0.85 | 1.56 | 0.45 |
| Hybrid | Days to Maturity (day) | Plant Height (cm) | Panicle Length (cm) | Spikelets Panicle−1 | Panicles Plant−1 | |||||
| N | S | N | S | N | S | N | S | N | S | |
| IR58025A × IR25571-31-1 | 1.57 ** | −0.03 | −0.38 | −4.44 ** | −0.04 | 2.80 ** | 19.34 ** | −14.25 * | 0.96 * | 1.15 ** |
| IR58025A × PR 1 | 1.67 ** | 1.02 | −1.79 | −3.38 ** | 0.00 | −0.87 ** | −6.68 * | −9.51 | −0.15 | 0.48 ** |
| IR58025A × PR 2 | −0.78 * | 0.40 | 1.91 | −0.14 | 0.18 | −0.55 ** | −5.71 * | 2.95 | −0.17 | −1.70 ** |
| IR58025A × Giza Basmati 201 | −2.46 ** | −1.38 * | 0.27 | 7.96 ** | −0.13 | −1.38 ** | −6.95 * | 20.81 ** | −0.65 | 0.08 |
| IR68902A × IR25571-31-1 | −1.41 ** | −0.67 | 1.37 | 7.98 ** | −2.44 ** | −1.36 ** | 0.12 | −11.85 * | −0.51 | −0.63 ** |
| IR68902A × PR 1 | 0.96 * | 0.17 | 1.63 | −2.29 ** | −4.77 ** | −2.00 ** | 1.27 | 20.23 ** | −0.62 | −0.69 ** |
| IR68902A × PR 2 | 1.81 ** | 1.06 | 1.32 | −1.06 | 2.96 ** | 1.06 ** | 1.48 | −6.65 | 1.16 * | 0.72 ** |
| IR68902A × Giza Basmati 201 | −1.37 ** | −0.57 | −4.32 * | −4.63 ** | 4.26 ** | 2.30 ** | −2.87 | −1.73 | −0.03 | 0.60 ** |
| Pusa 11A × IR25571-31-1 | 0.79 * | 0.95 | 5.12 ** | −2.03 ** | 4.81 ** | 1.44 ** | −14.12 ** | 9.84 | −1.26 * | 0.03 |
| Pusa 11A × PR 1 | −1.25 ** | −0.59 | −1.29 | 1.71 * | −2.62 ** | −1.72 ** | 4.52 | −32.45 ** | −0.70 | −0.93 ** |
| Pusa 11A × PR 2 | −0.83 * | 0.04 | −0.59 | −2.39 ** | 0.26 | 1.46 ** | 1.18 | 30.38 ** | 2.08 ** | 1.46 ** |
| Pusa 11A × Giza Basmati 201 | 1.29 ** | −0.41 | −3.23 * | 2.71 ** | −2.45 ** | −1.18 ** | 8.42 ** | −7.77 | −0.11 | −0.56 ** |
| Pusa 12A × IR25571-31-1 | 0.47 | 1.21 | 0.95 | 3.98 ** | 0.76 | −2.63 ** | −13.38 ** | 13.34 * | 1.17 * | 0.14 * |
| Pusa 12A × PR 1 | −1.46 ** | −1.21 | −2.12 | −5.29 ** | 5.47 ** | 3.93 ** | 4.61 | 6.74 | −0.01 | 0.60 ** |
| Pusa 12A × PR 2 | −1.18 ** | −1.61 * | −4.76 ** | 2.94 ** | −3.14 ** | −1.08 ** | 10.00 ** | −31.13 ** | −0.57 | 0.01 |
| Pusa 12A × Giza Basmati 201 | 2.18 ** | 1.61 * | 5.93 ** | −1.63 * | −3.09 ** | −0.23 ** | −1.22 | 11.06 * | −0.59 | −0.75 ** |
| Pusa 13A × IR25571-31-1 | −1.42 ** | −1.46 * | −7.05 ** | −5.48 ** | −3.08 ** | −0.24 ** | 8.04 ** | 2.92 | −0.37 | −0.69 ** |
| Pusa 13A × PR 1 | 0.08 | 0.60 | 3.57 * | 9.25 ** | 1.92 ** | 0.65 ** | −3.72 | 14.99 * | 1.49 ** | 0.54 ** |
| Pusa 13A × PR 2 | 0.99 * | 0.11 | 2.13 | 0.65 | -0.25 | −0.90 ** | −6.95 * | 4.45 | −2.50 ** | −0.50 ** |
| Pusa 13A × Giza Basmati 201 | 0.35 | 0.75 | 1.35 | −4.42 ** | 1.42 ** | 0.49 ** | 2.62 | −22.36 ** | 1.38 ** | 0.64 ** |
| LSD 5% | 0.70 | 1.29 | 3.23 | 1.40 | 0.94 | 0.12 | 5.15 | 10.66 | 0.94 | 0.11 |
| 1% | 1.00 | 1.85 | 4.62 | 2.01 | 1.35 | 0.17 | 7.37 | 15.24 | 1.34 | 0.16 |
| Hybrid | Panicle Weight (g) | Filled Grains Panicle−1 | 1000 Grain Weight (g) | Spikelet Fertility% | Grain Yield Plant−1 | |||||
| N | S | N | S | N | S | N | S | N | S | |
| IR58025A × IR25571-31-1 | −0.03 | −0.32 ** | 20.63 ** | −14.16 ** | −0.54 ** | −0.21 * | 1.25 | −0.23 | −1.33 | −3.09 ** |
| IR58025A × PR 1 | 0.17 | −0.42 ** | −9.46 ** | −14.16 ** | 0.38 ** | 0.46 ** | −1.50 * | −2.18 ** | −7.71 ** | 1.89 ** |
| IR58025A × PR 2 | 0.20 | −0.26 ** | −6.77 * | −6.99 ** | 0.17 | 0.01 | −0.56 | −4.98 ** | 2.12 | 4.80 ** |
| IR58025A × Giza Basmati 201 | −0.34 * | 1.00 ** | −4.41 | 35.31 ** | −0.01 | −0.26 ** | 0.81 | 7.39 ** | 6.92 ** | −3.61 ** |
| IR68902A × IR25571-31-1 | −0.15 | −0.57 ** | −1.73 | −22.28 ** | 0.60 ** | −0.19 * | −0.85 | −5.73 ** | 8.24 ** | −3.74 ** |
| IR68902A × PR 1 | 0.21 | 0.82 ** | 4.54 | 19.88 ** | −0.45 ** | 0.05 | 1.49 * | 2.02 ** | 0.44 | 2.12 ** |
| IR68902A × PR 2 | 0.07 | 0.03 | −2.72 | 2.05 ** | −0.44 ** | 0.37 ** | −1.67 * | 4.61 ** | 1.23 | 3.71 ** |
| IR68902A × Giza Basmati 201 | −0.13 | −0.28 ** | −0.09 | 0.35 | 0.29 * | −0.23 * | 1.03 | −0.89 | −9.91 ** | −2.09 ** |
| Pusa 11A × IR25571-31-1 | 0.23 | 0.46 ** | −15.95 ** | 1.72 ** | 0.29 * | 1.19 ** | −1.37 * | −3.14 ** | −0.50 | −2.50 ** |
| Pusa 11A × PR 1 | −0.09 | −1.06 ** | 7.22 * | −37.62 ** | −0.12 | −0.36 ** | 1.28 * | −4.97 ** | 4.09 ** | 3.33 ** |
| Pusa 11A × PR 2 | −0.06 | 0.59 ** | 2.69 | 27.38 ** | 0.23 | −0.43 ** | 0.71 | 1.83 * | 0.75 | 1.78 ** |
| Pusa 11A × Giza Basmati 201 | −0.09 | 0.01 | 6.04 * | 8.52 ** | −0.40 ** | −0.40 ** | −0.62 | 6.28 ** | −4.35 ** | −2.61 ** |
| Pusa 12A × IR25571-31-1 | 0.29 * | 0.43 ** | −11.30 ** | 17.55 ** | -0.32 * | -0.40 ** | 0.58 | −0.08 | 3.40 * | 8.14 ** |
| Pusa 12A × PR 1 | −0.07 | 0.67 ** | 3.83 | 25.72 ** | −0.06 | −0.38 ** | −0.17 | 6.45 ** | −12.64 ** | −8.80 ** |
| Pusa 12A × PR 2 | −0.31 * | −0.36 ** | 12.56 ** | −22.45 ** | −0.10 | −0.50 ** | 1.20 | −0.32 | −0.41 | −6.28 ** |
| Pusa 12A × Giza Basmati 201 | 0.09 | −0.74 ** | −5.09 | −20.82 ** | 0.48 ** | 1.29 ** | −1.61 * | −6.06 ** | 9.65 ** | 6.95 ** |
| Pusa 13A × IR25571-31-1 | −0.34 * | 0.00 | 8.35 * | 17.18 ** | −0.02 | −0.40 ** | 0.39 | 9.17 ** | −9.81 ** | 1.18 ** |
| Pusa 13A × PR 1 | −0.22 | −0.01 | −6.14 * | 6.18 ** | 0.24 | 0.24 * | −1.10 | −1.32 | 15.82 ** | 1.46 ** |
| Pusa 13A × PR 2 | 0.09 | 0.00 | −5.76 | 0.01 | 0.14 | 0.55 ** | 0.31 | −1.13 | −3.69 ** | −4.01 ** |
| Pusa 13A × Giza Basmati 201 | 0.47 ** | 0.02 | 3.55 | −23.36 ** | −0.36 * | −0.40 ** | 0.40 | −6.72 ** | −2.32 | 1.37 ** |
| LSD 5% | 0.28 | 0.13 | 5.87 | 0.79 | 0.26 | 0.18 | 1.26 | 1.33 | 2.43 | 0.70 |
| 1% | 0.40 | 0.19 | 8.39 | 1.13 | 0.37 | 0.26 | 1.80 | 1.90 | 3.48 | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Mowafi, H.F.; AlKahtani, M.D.F.; Abdallah, R.M.; Reda, A.M.; Attia, K.A.; El-Hity, M.A.; El-Dabaawy, H.E.; Husnain, L.A.; Al-Ateeq, T.K.; EL-Esawi, M.A. Combining Ability and Gene Action for Yield Characteristics in Novel Aromatic Cytoplasmic Male Sterile Hybrid Rice under Water-Stress Conditions. Agriculture 2021, 11, 226. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030226
El-Mowafi HF, AlKahtani MDF, Abdallah RM, Reda AM, Attia KA, El-Hity MA, El-Dabaawy HE, Husnain LA, Al-Ateeq TK, EL-Esawi MA. Combining Ability and Gene Action for Yield Characteristics in Novel Aromatic Cytoplasmic Male Sterile Hybrid Rice under Water-Stress Conditions. Agriculture. 2021; 11(3):226. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030226
Chicago/Turabian StyleEl-Mowafi, Hamdi F., Muneera D. F. AlKahtani, Rizk M. Abdallah, Amr M. Reda, Kotb A. Attia, Mahmoud A. El-Hity, Hend E. El-Dabaawy, Latifa Al Husnain, Talal K. Al-Ateeq, and Mohamed A. EL-Esawi. 2021. "Combining Ability and Gene Action for Yield Characteristics in Novel Aromatic Cytoplasmic Male Sterile Hybrid Rice under Water-Stress Conditions" Agriculture 11, no. 3: 226. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030226