
Production and Molecular Identification of Interspecific Hybrids between Phaius mishmensis (Lindl. and Paxton) Rchb. f. and Phaius tankervilliae (Banks) Blume

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Self-Incompatibility and Crossability Test
2.3. Comparative Characteristics among Intra- and Interspecific Hybridizations
2.4. Morphological and Anatomical Analysis
2.5. Development and Validation of Sequence-Characterized Amplified Region (SCAR) Markers for Phaius Hybrid Identifications
3. Results
3.1. Self-Incompatibility and Cross Ability
3.2. Intra- and Interspecific crossing Potential
3.3. Comparative Characteristics of Pod and Embryo among Inter- and Intraspecific Hybridizations
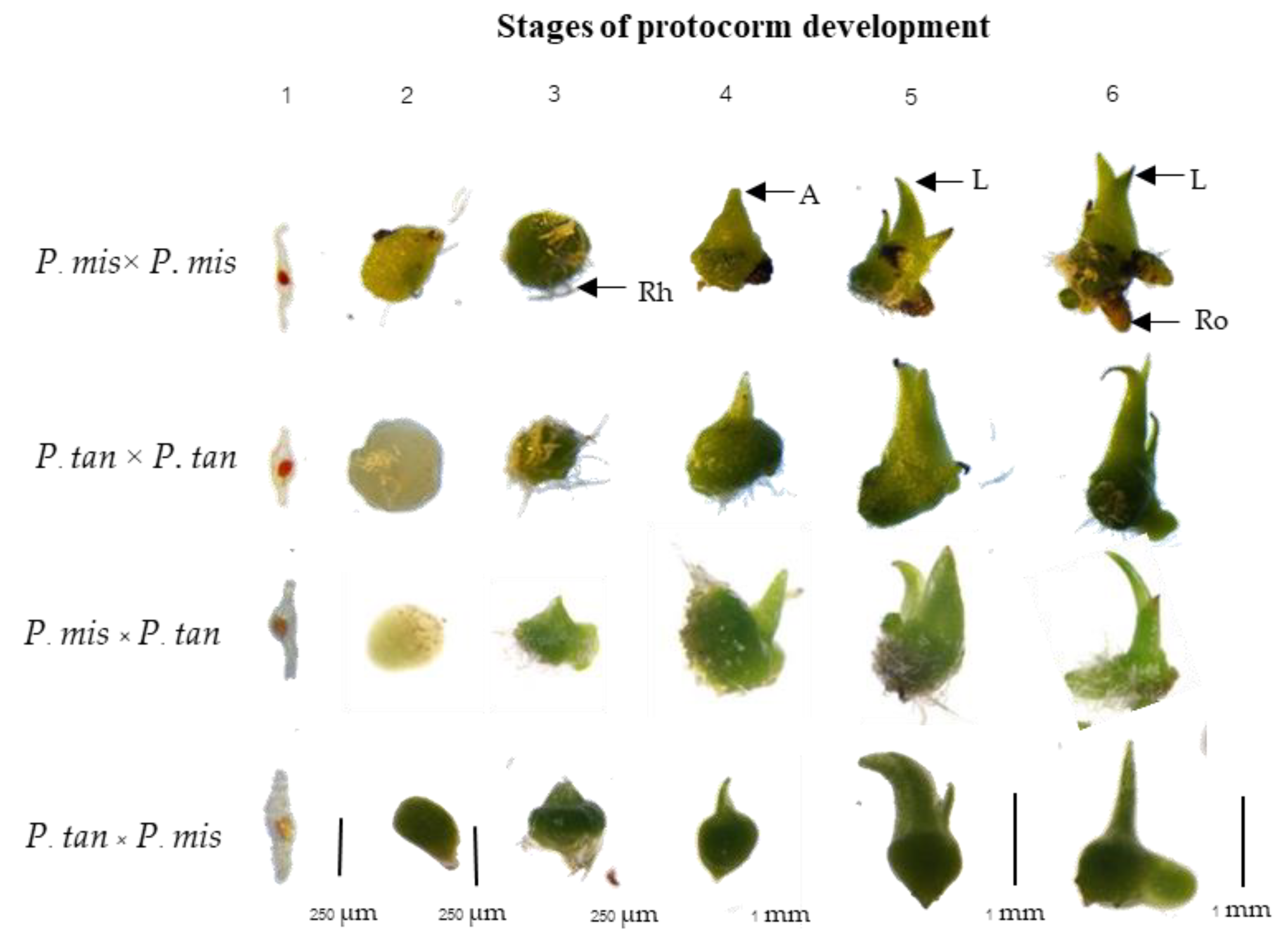
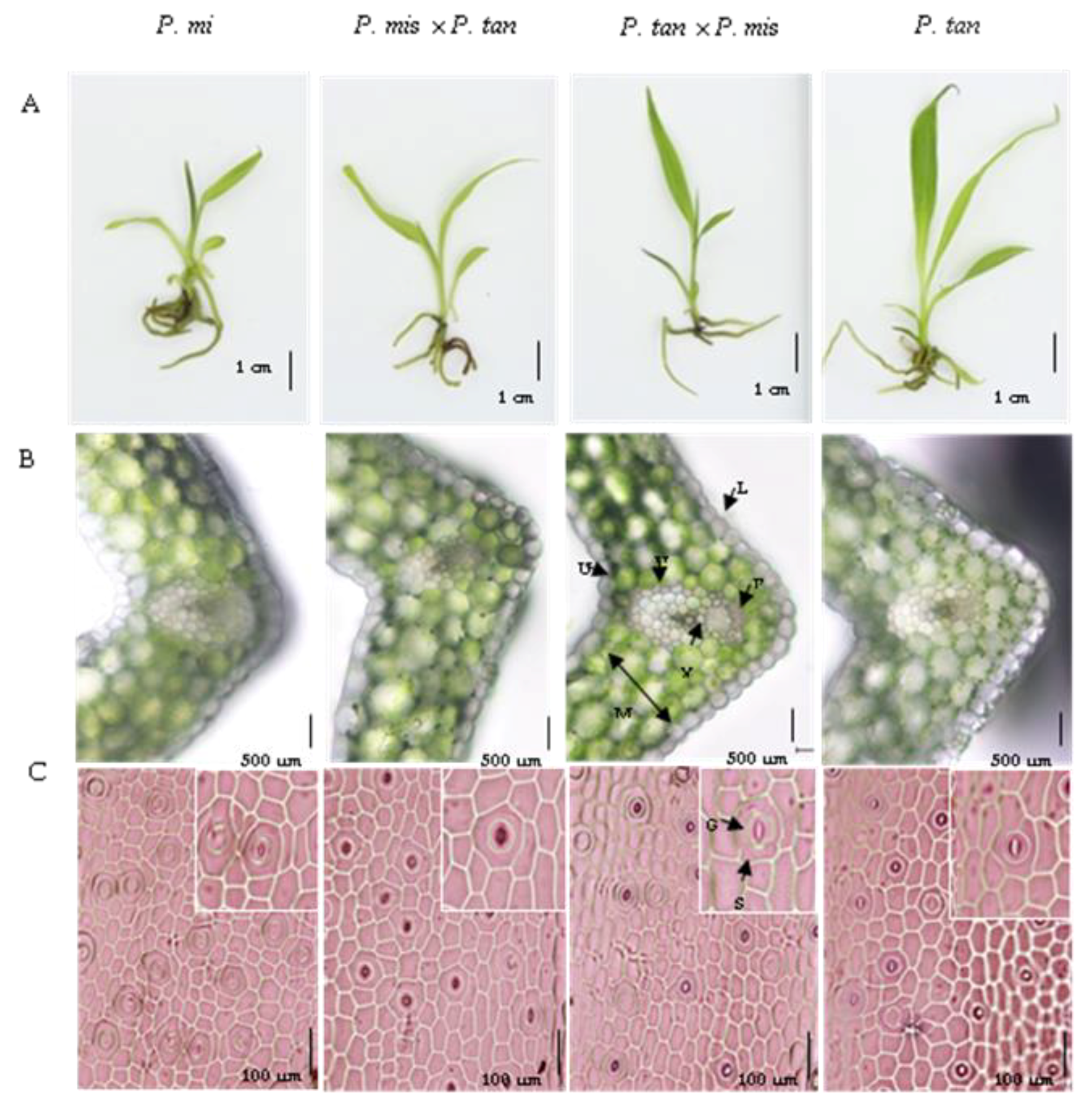
3.4. Morphology and Leaf Anatomy of Young Hybrid Seedlings
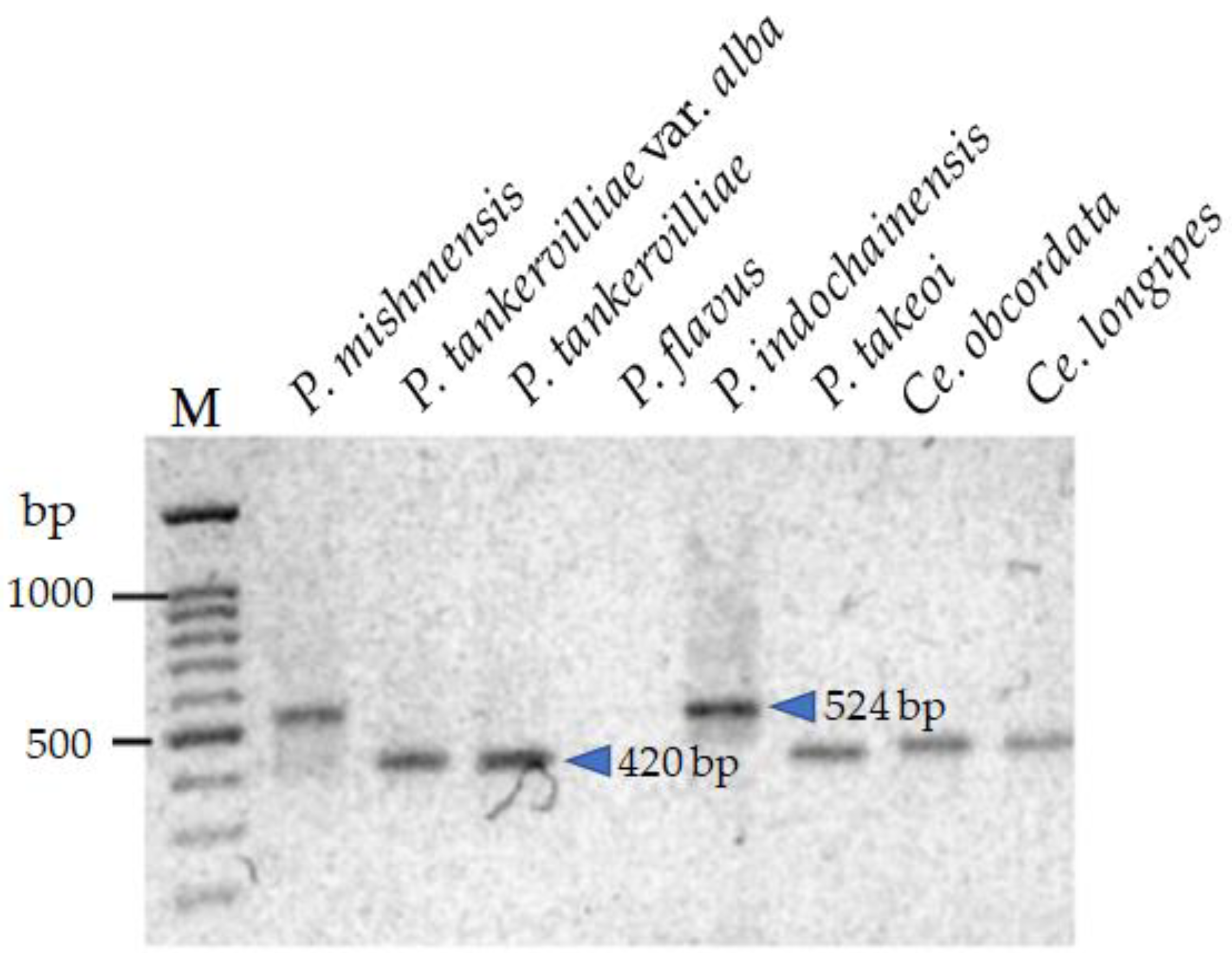
3.5. Development of Sequence-Characterized Amplified Region (SCAR) Markers for Identification of Phaius Hybrids
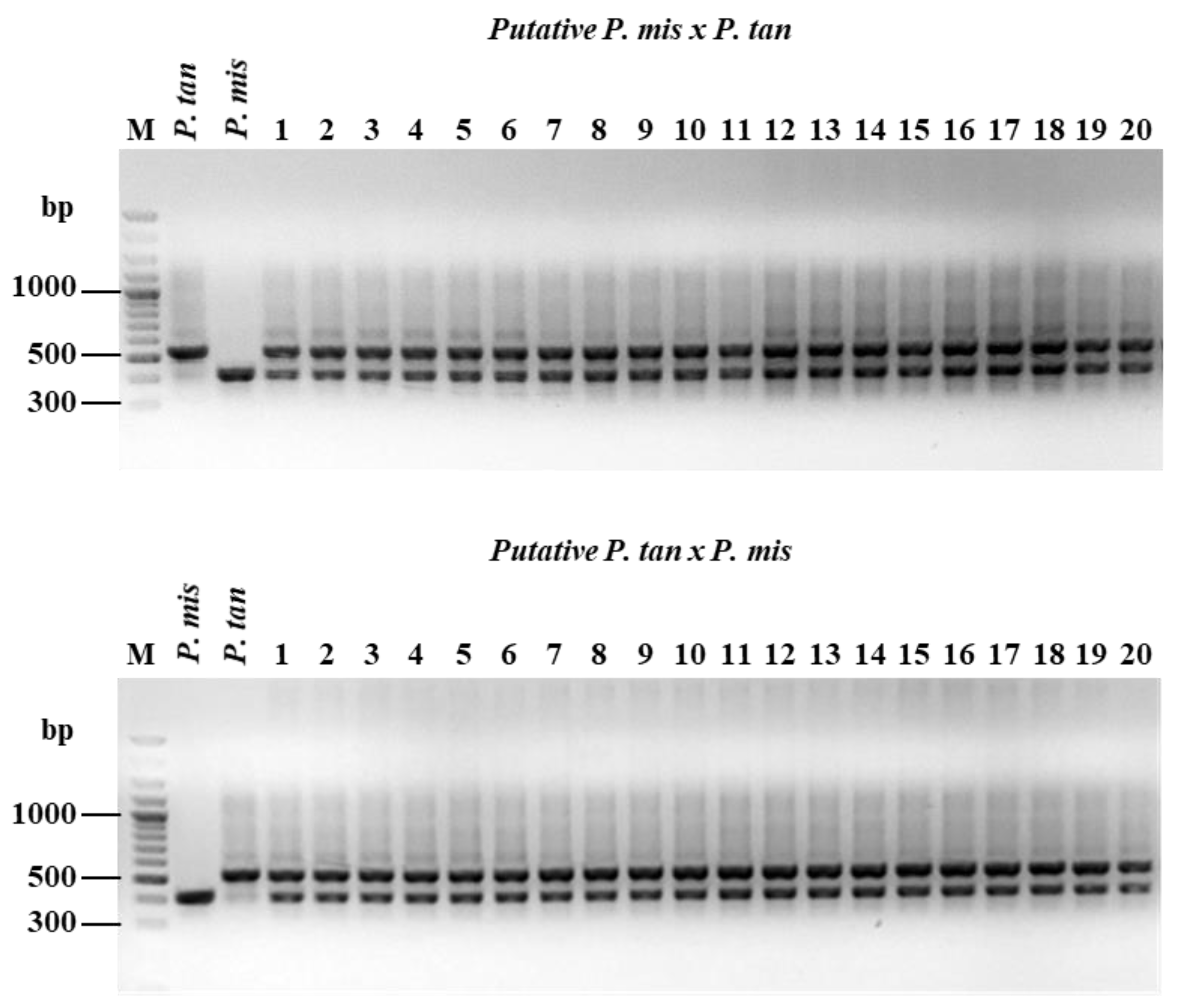
3.6. Confirmation of SCAR Primers with Hybrids Determination and Confirmation of Putative Hybrids were Conducted Using Pmis524 SCAR Primer Pairs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christenhusz, M.J.; Fay, M.F.; Chase, M.W. Plants of the World: An Illustrated Encyclopedia of Vascular Plants; University of Chicago Press: Chicago, IL, USA, 2017; p. 816. [Google Scholar]
- Kurzweil, H. A Precursory study of the Calanthe Group (Orchidaceae) in Thailand. Natl. Parks. Adansonia 2010, 32, 57–107. [Google Scholar] [CrossRef]
- Santisuk, T.; Larsen, K. Flora of Thailand; Orchidaceae 1 (Cypripedioideae, Orchidoideae, Vanilloideae); The Forest Herbarium, National Park, Wildlife and plant Conservation Department: Bangkok, Thailand, 2011; p. 12. [Google Scholar]
- Santisuk, T.; Chayamarit, K.; Pooma, R.; Sudee, S. Thailand Red Data: Plants; Integrated Promotion Technology: Bangkok, Thailand, 2006; p. 214. [Google Scholar]
- Tomonari, H.; Godo, T.; Miyoshi, K.; Ishikawa, K.; Ishikawa, M.; Mii, M. Cryopreservation and low-temperature storage of seeds of Phaius tankervilleae. Plant Biotechnol. Rep. 2009, 3, 103–109. [Google Scholar]
- Yam, T.W.; Arditti, J. History of orchid propagation: A mirror of the history of biotechnology. Plant Biotechnol. Rep. 2009, 3, 1–56. [Google Scholar] [CrossRef] [Green Version]
- Chugh, S.; Guha, S.; Rao, I.U. Micropropagation of orchids: A review on the potential of different explants. Sci. Hortic. 2009, 122, 507–520. [Google Scholar] [CrossRef]
- Lepick, N. NYBG’s orchid show Singapore. Mos Newslett. 2019, 1–15. [Google Scholar]
- Devadas, R.; Pamarthi, R.K.; Meitei, A.L.; Pattanayak, S.L.; Sherpa, R. Morphological description of novel Phaius primary hybrid (Orchidaceae). J. Exp. Biol. Agric. Sci. 2019, 7, 138–147. [Google Scholar] [CrossRef]
- Pérez, T.; Albornoz, J.; Domínguez, A. An evaluation of RAPD fragment reproducibility and nature. Mol. Ecol. 1998, 7, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Paran, I.; Michelmore, R.W. Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce. Theor. Appl. Genet. 1993, 85, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Sairkar, P.K.; Sharma, A.; Shukla, N.P. SCAR marker for identification and discrimination of Commiphora wightii and C. myrrha. Mol. Biol. Int. 2016, 1, 1–10. [Google Scholar] [CrossRef]
- Lee, M.Y.; Doh, E.J.; Park, C.H.; Kim, Y.H.; Kim, E.S.; Ko, B.S.; Ok, S.E. Development of SCAR marker for discrimination of Artemisia princeps and A. argyi from other Artemisia herbs. Biol. Pharm. Bull. 2006, 29, 629–633. [Google Scholar] [CrossRef] [Green Version]
- Devaiah, K.M.; Venkatasubramanian, P. Development of SCAR marker for authentication of Pueraria tuberosa (Roxb. Ex. Willd.) DC. Curr. Sci. 2008, 94, 1306–1309. [Google Scholar]
- Julio, E.; Verrier, J.; Dorlhac de Borne, F. Development of SCAR markers linked to three disease resistances based on AFLP within Nicotiana tabacum L. Theor. Appl. Genet. 2006, 112, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Yung, W.S.; Liaoa, Y.J.; Hunga, Y.S.; Changa, J.C.; Sung, J.M. Development of ITS sequence based SCAR markers for discrimination of Paphiopedilum armeniacum, P. micranthum, P. delenatii and their Hybrids. Sci. Hortic. 2011, 127, 405–410. [Google Scholar]
- Rodger, J.G.; Ellis, A.G. Distinct effects of pollinator dependence and self-incompatibility on pollen limitation in South African biodiversity hotspots. Biol. Lett. 2016, 12, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Takao, H.S.; Custódio, C.C.; Seaton, P.T.; Timothy, R.M.; Machado-Neto, N.B. Improved assessment of viability and germination of Cattleya (Orchidaceae) seeds following storage. In Vitro Cell. Dev. Biol. Plant 2012, 48, 27–36. [Google Scholar]
- Vacin, E.F.; Went, F.W. Some pH changes in nutrient solutions. Bot. Gazette. 1949, 110, 605–613. [Google Scholar] [CrossRef]
- Miyoshi, K.; Mii, M. Enhancement of seed germination and protocorm formation in Calanthe discolor (Orchidaceae) by NaOCl and polyphenol absorbent treatments. Plant Tiss. Cult. Lett. 1995, 12, 267–272. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.D. Isolation of plant DNA from fresh tissue. Electron. J. Biotechnol. 1990, 12, 13–15. [Google Scholar]
- Arditti, J. An history of orchid hybridization, seed germination and tissue culture. Bot. J. Linn. Soc. 1984, 89, 359–381. [Google Scholar] [CrossRef]
- Grantham, M.A.; Ford, B.A.; Worley, A.C. Pollination and fruit set in two rewardless slipper orchids and their hybrids (Cypripedium, Orchidaceae): Large yellow flowers outperform small white flowers in the northern tall grass prairie. Plant Biol. 2019, 21, 997–1007. [Google Scholar] [CrossRef]
- Udomdee, W.; Kongsawad, P.; Darak, R.; Somrit, P. Collection and evaluation on ex situ conservation of Phaius orchid. Khon Kaen Agr. J. 2014, 3, 529–534. [Google Scholar]
- Harrison, D.K.; Kwan, H.; Johnston, M.E.; Harris, W.K. Molecular taxonomy of the Australian swamp orchids (Phaius spp.). Acta Hortic. 2005, 694, 121–124. [Google Scholar] [CrossRef]
- Zhai, J.W.; Zhang, G.Q.; Li, L.; Wang, M.; Chen, L.J.; Chung, S.W.; Rodriguez, F.J.; Ortega, J.F.; Lan, S.R.; Xing, F.W.; et al. A new phylogenetic analysis sheds new light on the relationships in the Calanthe alliance (Orchidaceae) in China. Mol. Phylogenet. Evol. 2014, 77, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Teoh, S.B. Cytological studies in Malayan members of the Phaius tribe (Orchidaceae). Ii. Meiotic and b-Chromosomes. Caryologia 1980, 33, 483–493. [Google Scholar] [CrossRef]
- Jones, N. New species with B chromosomes discovered since 1980. Nucleus 2017, 60, 263–281. [Google Scholar] [CrossRef]
- Proctor, H.C. Effect of pollen age on fruit set, fruit weight and seed set in three orchid species. Can. J. Bot. 1998, 76, 420–427. [Google Scholar]
- Shiau, Y.J.; Sagare, A.P.; Chen, U.C.; Yang, S.R.; Tsay, H.S. Conservation of Anoectochilus formosanus Hayata by artificial cross pollination and in vitro culture of seeds. Bot. Bull. Acad. Sin. 2002, 43, 123–130. [Google Scholar]
- Edmands, S. Does parental divergence predict reproductive compatibility. Trends Ecol. Evol. 2002, 17, 520–527. [Google Scholar] [CrossRef]
- Devi, J.; Deka, P.C. Pollen viability, stigma receptivity and cross compatibility of some Indian orchids. J. Orchid Soc. India. 1992, 6, 79–84. [Google Scholar]
- Pershinaa, L.A.; Trubacheeva, N.V. Interspecific incompatibility in the wide hybridization of plants and ways to overcome it. Russ. J. Genet. Appl. Res. 2017, 7, 358–368. [Google Scholar] [CrossRef]
- Johansen, B. Incompatibility in Dendrodium (Orchidaceae). Bot. J. Linn. Soc. 1990, 103, 165–196. [Google Scholar] [CrossRef]
- Barbosa, A.R.; de Melo, M.C.; Borba, E.L. Self-incompatibility and myophily in Octomeria (Orchidaceae, Pleurothallidinae) species. Plant Syst. Evol. 2009, 283, 1–8. [Google Scholar] [CrossRef]
- Charlesworth, D.; Charlesworth, B. Inbreeding depression and its evolutionary consequences. Ann. Rev. Ecol. Evol. Syst. 1987, 18, 237–268. [Google Scholar] [CrossRef]
- Bellusci, F.; Pellegrino, G.; Musacchio, A. Different levels of inbreeding depression between outcrossing and selfing Serapias species. Biol. Plant 2009, 53, 175–178. [Google Scholar] [CrossRef]
- Petit, S.; Jusaitis, M.; Bickerton, D. Effect of pollen load, self-pollination and plant size on seeds and germination in the endangered pink-lipped spider orchid, Caladenia behrii. Aust. J. Bot. 2009, 57, 307–314. [Google Scholar] [CrossRef]
- Pakum, W.; Watthana, S.; Srimuang, K.; Kongbangkerd, A. Influence of medium component on in vitro propagation of Thai’s endangered orchid: Bulbophyllum nipondhii Seidenf. Plant Tiss. Cult. Biotechnol. 2016, 25, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Kunakhonnuruk, B.; Inthima, P.; Kongbangkerd, A. In vitro propagation of Epipactis flava Seidenf. an endangered rheophytic orchid: A first study on factors affecting asymbiotic seed germination, seedling development and greenhouse acclimatization. Plant Cell Tiss. Organ Cult. 2018, 135, 419–432. [Google Scholar] [CrossRef]
- Moccia, M.D.; Widmer, A.; Cozzolino, S. The strength of reproductive isolation in hybridizing food deceptive orchids. Mol. Ecol. 2007, 16, 2855–2866. [Google Scholar] [CrossRef]
- Busse, A.J.; Gola, E.M. Morphological variability of Helleborines. I. Diagnostic significance of morphological features in Epipactis helleborine (L.) Crantz, Epipactis atrorubens (Hoffm.) Besser and their hybrid, Epipactis × schmalhausenii Richt. (Orchidaceae, Neottieae). Acta Soc. Bot. Pol. 2010, 79, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Devadas, R.; Medhi, R.P.; Das, S.P. Interspecific hybrid developed in Epidendrum orchid from the cross E. radicans Pav. Ex. Lindl. × E. xanthinum Lindl. J. Hortic. Sci. 2010, 5, 144–147. [Google Scholar]
- Sinumporn, P.; Narumi-Kawasaki, T.; Fukai, S. Development of interspecific hybrids between Habenaria radiata and Habenaria rhodocheila complex. Adv. Hort. Sci. 2020, 34, 3–10. [Google Scholar]
- Punjansing, T.; Chaichanachap, N.; Sanitchon, J.; Pinta, W.; Nakkuntod, M. Genetic diversity of some species in the genus Calanthe analyzed by RAPD markers. Acta Hortic. 2017, 1167, 323–330. [Google Scholar] [CrossRef]
- Nutthapornnitchakul, S.; Peyachoknagul, S.; Sangin, P.; Kongbungkerd, A.; Punjansing, T.; Nakkuntod, M. Genetic relationship of orchids in the Calanthe group based on sequence-related amplified polymorphism markers and development of sequence-characterized amplified regions markers for some genus/species identification. Agric. Nat. Resour. 2019, 53, 340–347. [Google Scholar]
- Handa, T. Utilization of molecular markers for ornamental plants. J. Am. Soc. Hortic. Sci. 1998, 67, 1197–1199. [Google Scholar] [CrossRef]
- Malay, D.; Bhattacharya, S.; Pal, A. Generation and characterization of SCARs by cloning and sequencing of RAPD products: A strategy for species- specific marker development in bamboo. Ann. Bot. 2005, 95, 835–841. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | % Pod Setting (n) | % Embryo Formation | % Seed Viability | % In Vitro Seed Germination |
|---|---|---|---|---|
| M (♀) × M (♂) | 62.5 (40) | 34.0 ± 0.2 c | 29.9 ± 1.9 c | 72.2 ± 4.1 b |
| T (♀) × T (♂) | 60.0 (40) | 100.0 ± 0.0 a | 99.8 ± 0.2 a | 70.8 ± 3.1 b |
| M (♀) × T (♂) | 80.0 (20) | 48.3 ± 0.7 b | 44.6 ± 1.0 b | 94.8 ± 0.8 a |
| T (♀) × M (♂) | 25.0 (20) | 9.3 ± 0.8 d | 8.2 ± 0.6 d | 77.3 ± 6.8 b |
| Treatment | Average Days of Pod Dehiscence (Days after Pollination) | Pod Size (cm) | Pod Weight (g) | Embryo Size (µm) | ||
|---|---|---|---|---|---|---|
| Width | Length | Width | Length | |||
| M (♀) × M (♂) | 50 | 1.9 ± 0.5 ab | 4.7 ± 0.4 b | 4.1 ± 0.8 c | 55.4 ± 0.4 b | 94.8 ± 1.5 c |
| T (♀) × T (♂) | 147 | 2.4 ± 0.7 a | 5.3 ± 1.0 ab | 10.0 ± 3.8 a | 87.7 ± 1.6 a | 145.7 ± 1.5 a |
| M (♀) × T (♂) | 60 | 2.0 ± 0.2 ab | 5.1 ± 0.4 b | 6.8 ± 1.7 b | 46.5 ± 0.6 c | 67.3 ± 0.9 d |
| T (♀) × M (♂) | 105 | 1.6 ± 0.5 b | 5.4 ± 0.2 a | 6.2 ± 1.8 bc | 86.2 ± 0.9 a | 113.0 ± 6.9 b |
| Treatment | Stage of Protocorm Development (%) | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| M (♀) × M (♂) | 27.7 ± 4.1 a | 21.5 ± 1.5 c | 16.6 ± 1.7 a | 20.2 ± 2.6 ns | 10.6 ± 2.1 ns | 3.1 ± 0.5 b |
| T (♀) × T (♂) | 29.1 ± 3.1 a | 28.0 ± 3.1 b | 11.1 ± 1.7 c | 17.5 ± 2.7 | 10.4 ± 1.6 | 3.7 ± 0.7 b |
| M (♀) × T (♂) | 5.1 ± 0.8 b | 38.0 ± 1.4 a | 14.6 ± 1.7 ab | 22.0 ± 3.0 | 9.5 ± 0.2 | 10.6 ± 1.7 a |
| T (♀) × M (♂) | 22.6 ± 6.8 a | 23.1 ± 1.2 bc | 12.6 ± 1.3 ab | 21.1 ± 3.1 | 14.8 ± 3.5 | 5.5 ± 1.7 b |
| Plants Tested | Leaf Size (cm) | Leaf Area (cm2) | Thickness of Leaf (µm) | Stomata Density (Stomata/mm2) | Stomatal Size (µm) | No. of Subsidiary Cell/Stomata Complex | ||
|---|---|---|---|---|---|---|---|---|
| Length | Width | Length | Width | |||||
| M(♀) × M(♂) | 2.4 ± 0.1 c | 0.8 ± 0.1 a | 1.6 ± 0.1 b | 294.5 ± 6.2 a | 35.2 ± 0.4 a | 34.0 ± 0.4 c | 32.8 ± 0.6 c | 3.8 ± 0.1 b |
| T(♀) × T(♂) | 4.5 ± 0.2 a | 0.6 ± 0.1 b | 2.5 ± 0.2 a | 212.6 ± 7.3 b | 27.1 ± 0.5 b | 43.4 ± 1.2 a | 38.2 ± 1.0 a | 3.8 ± 0.1 b |
| M(♀) × T(♂) | 3.7 ± 0.3 b | 0.7 ± 0.4 b | 2.2 ± 0.2 b | 222.9 ± 3.3 b | 26.4 ± 0.9 bc | 44.2 ± 0.2 a | 37.7 ± 0.8 a | 4.1 ± 0.1 a |
| T(♀) × M(♂) | 4.9 ± 0.2 a | 0.7 ± 0.4 b | 2.3 ± 0.2 a | 202.8 ± 7.8 b | 24.8 ± 0.3 c | 40.8 ± 0.5 b | 33.1 ± 0.7 b | 3.4 ± 0.1 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Punjansing, T.; Nakkuntod, M.; Homchan, S.; Inthima, P.; Kongbangkerd, A. Production and Molecular Identification of Interspecific Hybrids between Phaius mishmensis (Lindl. and Paxton) Rchb. f. and Phaius tankervilliae (Banks) Blume. Agriculture 2021, 11, 306. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040306
Punjansing T, Nakkuntod M, Homchan S, Inthima P, Kongbangkerd A. Production and Molecular Identification of Interspecific Hybrids between Phaius mishmensis (Lindl. and Paxton) Rchb. f. and Phaius tankervilliae (Banks) Blume. Agriculture. 2021; 11(4):306. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040306
Chicago/Turabian StylePunjansing, Tasanai, Maliwan Nakkuntod, Somjit Homchan, Phithak Inthima, and Anupan Kongbangkerd. 2021. "Production and Molecular Identification of Interspecific Hybrids between Phaius mishmensis (Lindl. and Paxton) Rchb. f. and Phaius tankervilliae (Banks) Blume" Agriculture 11, no. 4: 306. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040306