
A Review of Potassium-Rich Crop Residues Used as Organic Matter Amendments in Tree Crop Agroecosystems


Abstract
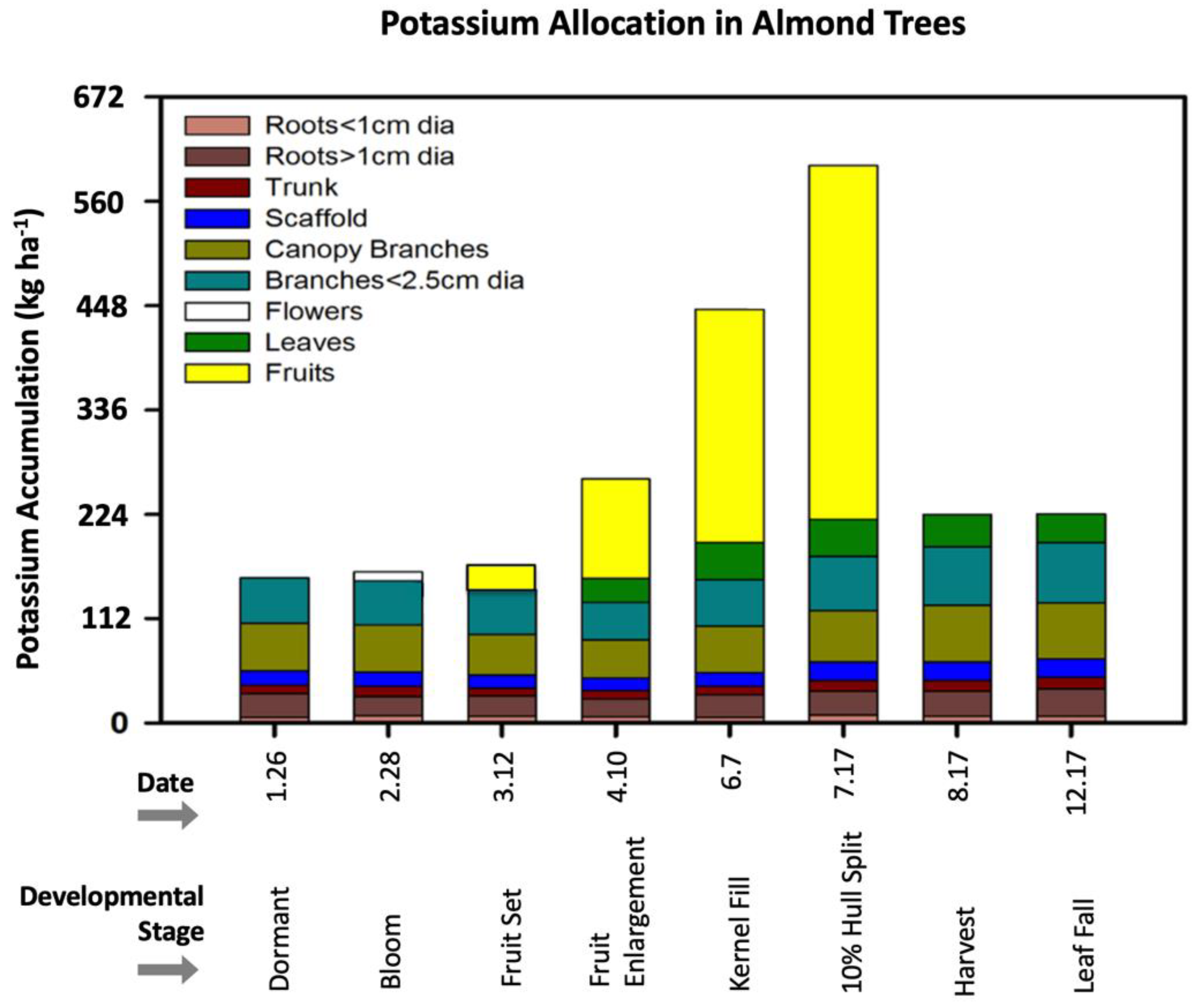
:1. Introduction
2. Residue Decomposition, Nitrogen Cycling, and Potassium Solubilization

{kind=link}
{kind=link}
| Source Crop | Material | Estimated %K | Region and Reference |
|---|---|---|---|
| Residues and biomass from tree crops and other permanent crops | |||
| Almond | hull | 3.2 | Murcia, Spain; Valverde et al., 2013 Table 2 |
| Almond | shell | 0.5 | Murcia, Spain; Valverde et al., 2013 Table 2 |
| Almond | hull | 3.3 | California, U.S.A.; Atkas et al., 2015 Table 6 |
| Almond | shell | 1.7 | California, U.S.A.; Atkas et al., 2015 Table 6 |
| Bhimal (Grewia) | leaf litter | 2.55 | Himachal Pradesh, India; Verma et al., 2012 Table 3 |
| Cacao | husk | 2.89 | Cote d’Ivoire and the Netherlands; Hougni et al., 2021 Table 1 |
| Cacao | husk | 3.18 | Review; Campos-Vega et al., 2018 Table 2 |
| Cacao | husk | 3.73 | Pingtung, Taiwan; Tsai and Huang et al., 2018 Table 2 |
| Cacao | husk | 2.8–3.8 | Review; Lu et al., 2018 Table 1 |
| Cacao | husk | 3.78 | Akure, Nigeria; Agele et al., 2008 Table 1 |
| Cacao | husk | 1.6 | Cote d’Ivoire; Kone et al., 2021 Discussion |
| Cacao | husk | 3.77–7.69 | Review; Hartemink et al., 2005 Table 3 |
| Coffee | husk | 4.57 | Brazil; Carmo et al., 2016 Table 2 |
| Coffee | pulp husk mixture | 2.49 | Kinshasha, Congo; Kasongo et al., 2011 Table 2 |
| Coffee | pulp | 3.89 | Sao Paulo, Brazil; Zoca et al., 2014 Table 2 |
| Coffee | husk | 3.47 | Sao Paulo, Brazil; Zoca et al., 2014 Table 2 |
| Gliricidia | leaves | 2.65 | Malaysia; Zahara et al., 1999 Results |
| Grape | stalk | 3.0 | Spain; Bustamante et al., 2008 Table 1 |
| Grape | pomace | 2.42 | Spain; Bustamante et al., 2008 Table 1 |
| Grape | wine lee | 7.28 | Spain; Bustamante et al., 2008 Table 1 |
| Hazelnut | husk | 4.29 | Turkey; Kizilkaya et al., 2008 Results |
| Olive | pruned material | 0.56 | Review; Zipori et al., 2020 Table 1 |
| Olive | leaf | 0.69–1.19 | Review; Zipori et al., 2020 Table 2 |
| Olive | olive mill waste | 2.10 | Italy; Altieri et al., 2008 Table 1 |
| Olive | olive mill waste | 2.6 | Spain; Cayuela et al., 2004 Table 1 |
| Pecan | husk | 3.47 | New Mexico, U.S.A.; Idowu et al., 2017 Table 2 |
| Pecan | shell | 0.15 | New Mexico, U.S.A.; Idowu et al., 2017 Table 2 |
| Poplar | leaf litter | 1.24 | Himachal Pradesh; Verma et al., 2012 Table 3 |
| Residues from row crops | |||
| Alfalfa | mulch | 2.2 | B.C., Canada; Neilsen et al., 2003 Table 1 |
| Black oat | green manure | 2.86 | Brazil; Franchini et al., 2003 Table 1 elongation stage |
| Black oat | residue | 4.22 | Brazil; Miyazawa et al., 2002 Table 1 |
| Maize | straw | 1.48 | Heilongjiang, China; Dong et al., 2019 Materials & Methods |
| Maize | residue | 1.53–1.69 | New Delhi, India; Madar et al., 2020 Table 1 |
| Radish | green manure | 3.85 | Brazil; Franchini et al., 2003 Table 1 vegetative stage |
| Rice | straw | 2.7 | Punjab, India; Yadav et al., 2019 Materials and Methods |
| Rice | residue | 2.1 | Punjab, India; Yadvinder-Singh et al., 2010 Methods and Materials |
| Rice | straw | 2.19 | Wuhan, China; Li et al., 2014 Materials and Methods |
| Rye | residue | 2.76 | Brazil; Miyazawa et al., 2002 Table 1 |
| Ryegrass cover crop | residue | 5.15 | Italy; Tagliavini et al., 2007 Table 1 |
| Soybean | straw | 1.05 | Heilongjiang, China; Dong et al., 2019 Materials & Methods |
| Sunflower | residue | 2.77 | Spain; Rodriguez-Lizana et al., 2010 Results |
| Wheat | straw | 1.22–1.91 | Jiangsu, China; Sui et al., 2014 Table 2 |
| Wheat | straw | 2.26–2.60 | New Delhi, India; Madar et al., 2020 Table 1 |
| Wheat | straw | 3.78 | Shaanxi, China; Wei et al., 2015 Table 1 |
3. Potassium Availability
4. Impacts of Increased Soil Health on Potassium Availability and Nutrient Cycling
5. Effects of Potassium-Rich Organic Matter Amendments on Crop Performance
6. Future Research Directions: Building the Organic Layer in Orchards and Implementation Considerations
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harter, T.; Lund, J.R. Addressing Nitrate in California’s Drinking Water: Executive Summary; University of California Center for Watershed Sciences: Davis, CA, USA, 2012. [Google Scholar]
- Drinkwater, L.E.; Snapp, S.S. Nutrients in Agroecosystems: Rethinking the Management Paradigm. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2007; Volume 92, pp. 163–186. [Google Scholar]
- Lal, R. Soils and sustainable agriculture. A review. Agron. Sustain. Dev. 2008, 28, 57–64. [Google Scholar] [CrossRef]
- Thomas, C.L.; Acquah, G.E.; Whitmore, A.P.; McGrath, S.P.; Haefele, S.M. The Effect of Different Organic Fertilizers on Yield and Soil and Crop Nutrient Concentrations. Agronomy 2019, 9, 776. [Google Scholar] [CrossRef] [Green Version]
- Thiyageshwari, S.; Gayathri, P.; Krishnamoorthy, R.; Anandham, R.; Paul, D. Exploration of Rice Husk Compost as an Alternate Organic Manure to Enhance the Productivity of Blackgram in Typic Haplustalf and Typic Rhodustalf. Int. J. Environ. Res. Public Health 2018, 15, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balentić, J.P.; Ačkar, Đ.; Jokić, S.; Jozinović, A.; Babić, J.; Miličević, B.; Šubarić, D.; Pavlović, N. Cocoa Shell: A By-Product with Great Potential for Wide Application. Molecules 2018, 23, 1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madar, R.; Singh, Y.V.; Meena, M.C.; Das, T.K.; Paramesh, V.; Al-Mana, F.A.; Mattar, M.A.; Elansary, H.O. Residue and Potassium Management Strategies to Improve Crop Productivity, Potassium Mobilization, and Assimilation under Zero-Till Maize-Wheat Cropping System. Agriculture 2020, 10, 401. [Google Scholar] [CrossRef]
- Esmaeili, A.; Khoram, M.R.; Gholami, M.; Eslami, H. Pistachio waste management using combined composting-vermicomposting technique: Physico-chemical changes and worm growth analysis. J. Clean. Prod. 2020, 242, 118523. [Google Scholar] [CrossRef]
- Karimi, H.R.; Sajjadinia, A.; Baghari, V.; Farahmand, H. Preliminary Evaluation of Composted Pistachio Hull and Rice Husk as Potting Medium on Scindapsus aureus Vegetative and Physiological Characteristics in Greenhouse Condition. J. Plant Nutr. 2013, 36, 2225–2235. [Google Scholar] [CrossRef]
- López, R.; Burgos, P.; Hermoso, J.M.; Hormaza, J.I.; González-Fernández, J.J. Long term changes in soil properties and enzyme activities after almond shell mulching in avocado organic production. Soil Tillage Res. 2014, 143, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Holtz, B.A.; Doll, D.; Browne, G. Whole almond orchard recycling and the effect on second generation tree growth, soil carbon, and fertility. Acta Hortic. 2015, 1112, 315–320. [Google Scholar] [CrossRef]
- Jahanzad, E.; Holtz, B.A.; Zuber, C.A.; Doll, D.; Brewer, K.M.; Hogan, S.; Gaudin, A.C.M. Orchard recycling improves climate change adaptation and mitigation potential of almond production systems. PLoS ONE 2020, 15, e0229588. [Google Scholar] [CrossRef]
- Hannam, K.D.; Neilsen, G.H.; Forge, T.A.; Neilsen, D.; Losso, I.; Jones, M.D.; Nichol, C.; Fentabil, M.M. Irrigation practices, nutrient applications, and mulches affect soil nutrient dynamics in a young Merlot (Vitis vinifera L.) vineyard. Can. J. Soil Sci. 2016, 96, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Kallenbach, C.M.; Wallenstein, M.D.; Schipanksi, M.E.; Grandy, A.S. Managing Agroecosystems for Soil Microbial Carbon Use Efficiency: Ecological Unknowns, Potential Outcomes, and a Path Forward. Front. Microbiol. 2019, 10, 1146. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.; Hu, C.; Oenema, O. Soil mulching significantly enhances yields and water and nitrogen use efficiencies of maize and wheat: A meta-analysis. Sci. Rep. 2015, 5, 16210. [Google Scholar] [CrossRef]
- Jafari, M.; Haghighi, J.A.P.; Zare, H. Mulching Impact on Plant Growth and Production of Rainfed Fig Orchards under Drought Conditions; Journal of Food Agriculture and Environment: Helsinki, Finland, 2012. [Google Scholar]
- Iqbal, M.; Ul-Hassan, A.; Van Es, H.M. Influence of Residue Management and Tillage Systems on Carbon Sequestration and Nitrogen, Phosphorus, and Potassium Dynamics of Soil and Plant and Wheat Production in Semi-arid Region. Commun. Soil Sci. Plant Anal. 2011, 42, 528–547. [Google Scholar] [CrossRef]
- Moore, K.J.; Anex, R.P.; Elobeid, A.E.; Fei, S.; Flora, C.B.; Goggi, A.S.; Jacobs, K.L.; Jha, P.; Kaleita, A.L.; Karlen, D.L.; et al. Regenerating Agricultural Landscapes with Perennial Groundcover for Intensive Crop Production. Agronomy 2019, 9, 458. [Google Scholar] [CrossRef] [Green Version]
- Lal, R. Soil organic matter content and crop yield. J. Soil Water Conserv. 2020, 75, 27A–32A. [Google Scholar] [CrossRef] [Green Version]
- Montanaro, G.; Xiloyannis, C.; Nuzzo, V.; Dichio, B. Orchard management, soil organic carbon and ecosystem services in Mediterranean fruit tree crops. Sci. Hortic. 2017, 217, 92–101. [Google Scholar] [CrossRef]
- Sardans, J.; Penuelas, J. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Tripler, C.E.; Kaushal, S.S.; Likens, G.E.; Walter, M.T. Patterns in potassium dynamics in forest ecosystems. Ecol. Lett. 2006, 9, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, S.; Sanden, B.L.; Saa, S.; Lampinen, B.D.; Smart, D.R.; Shackel, K.A.; DeJong, T.M.; Brown, P.H. Optimization of nitrogen and potassium nutrition to improve yield and yield parameters of irrigated almond (Prunus dulcis (Mill.) D.A. Webb). Sci. Hortic. 2018, 228, 204–212. [Google Scholar] [CrossRef]
- Alva, A.K.; Mattos, D., Jr.; Paramasivam, S.; Patil, B.; Dou, H.; Sajwan, K.S. Potassium Management for Optimizing Citrus Production and Quality. Int. J. Fruit Sci. 2006, 6, 3–43. [Google Scholar] [CrossRef]
- Zeng, Q.; Brown, P.H.; Holtz, B.A. Potassium Fertilization Affects Soil K, Leaf K Concentration, and Nut Yield and Quality of Mature Pistachio Trees. HortScience 2001, 36, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, S.; Sanden, B.L.; Lampinen, B.D.; Saa, S.; Siddiqui, M.I.; Smart, D.R.; Olivos, A.; Shackel, K.A.; de Jong, T.; Brown, P.H. Seasonal changes in nutrient content and concentrations in a mature deciduous tree species: Studies in almond (Prunus dulcis (Mill.) D.A. Webb). Eur. J. Agron. 2015, 65, 52–68. [Google Scholar] [CrossRef]
- Sui, N.; Zhou, Z.; Yu, C.; Liu, R.; Yang, C.; Zhang, F.; Song, G.; Meng, Y. Yield and potassium use efficiency of cotton with wheat straw incorporation and potassium fertilization on soils with various conditions in the wheat-cotton rotation system. Field Crop. Res. 2015, 172, 132–144. [Google Scholar] [CrossRef]
- Jiang, N.; Hu, B.; Wang, X.; Meng, Y.; Chen, B.; Zhou, Z. Effects of crop residue incorporation and inorganic potassium fertilization on soil potassium supply power. Arch. Agron. Soil Sci. 2019, 65, 1223–1236. [Google Scholar] [CrossRef]
- Kasongo, R.K.; Verdoodt, A.; Kanyankagote, P.; Baert, G.; van Ranst, E. Coffee waste as an alternative fertilizer with soil improving properties for sandy soils in humid tropical environments. Soil Use Manag. 2010, 27, 94–102. [Google Scholar] [CrossRef]
- Dong, S.; Ma, Z.; Wang, L.; Yan, C.; Liu, L.; Gong, Z.; Cui, G. Decomposition and Nutrient Release Characteristics of Incorporated Soybean and Maize Straw in Northeast China. Ekoloji 2019, 107, 2119–2129. [Google Scholar]
- Zipori, I.; Erel, R.; Yermiyahu, U.; Ben-Gal, A.; Dag, A. Sustainable Management of Olive Orchard Nutrition: A Review. Agriculture 2020, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Rafique, E.; Mahmood-Ul-Hassan, M.; Rashid, A.; Chaudhary, M.F. Nutrient Balances as Affected by Integrated Nutrient and Crop Residue Management in Cotton-Wheat System in Aridisols. I. Nitrogen. J. Plant Nutr. 2012, 35, 591–616. [Google Scholar] [CrossRef]
- Singh, V.K.; Dwivedi, B.S.; Singh, Y.; Singh, S.K.; Mishra, R.P.; Shukla, A.K.; Rathore, S.S.; Shekhawat, K.; Majumdar, K.; Jat, M.L. Effect of tillage and crop establishment, residue management and K fertilization on yield, K use efficiency and apparent K balance under rice-maize system in north-western India. Field Crop. Res. 2018, 224, 1–12. [Google Scholar] [CrossRef]
- Yadav, S.K.; Benbi, D.K.; Toor, A.S. Effect of long-term application of rice straw, farmyard manure and inorganic fertilizer on potassium dynamics in soil. Arch. Agron. Soil Sci. 2019, 65, 374–384. [Google Scholar] [CrossRef]
- Yan, S.; Song, J.; Fan, J.; Yan, C.; Dong, S.; Ma, C.; Gong, Z. Changes in soil organic carbon fractions and microbial community under rice straw return in Northeast China. Glob. Ecol. Conserv. 2020, 22, e00962. [Google Scholar] [CrossRef]
- Öborn, I.; Andrist-Rangel, Y.; Askekaard, M.; Grant, C.A.; Watson, C.A.; Edwards, A.C. Critical aspects of potassium management in agricultural systems. Soil Use Manag. 2008, 21, 102–112. [Google Scholar] [CrossRef]
- Nagao, M.A.; Hirae, H.H.; Stephenson, R.A. Macadamia: Cultivation and physiology. Crit. Rev. Plant Sci. 1992, 10, 441–470. [Google Scholar] [CrossRef]
- Tagliavini, M.; Tonon, G.; Scandellari, F.; Quinones, A.; Palmieri, S.; Menarbin, G.; Gioacchini, P.; Masia, A. Nutrient recycling during the decomposition of apple leaves (Malus domestica) and mowed grasses in an orchard. Agric. Ecosyst. Environ. 2007, 118, 191–200. [Google Scholar] [CrossRef]
- Wartelle, L.H.; Marshall, W.E. Nutshells as granular activated carbons: Physical, chemical and adsorptive properties. J. Chem. Technol. Biotechnol. 2001, 76, 451–455. [Google Scholar] [CrossRef]
- Neilsen, G.; Forge, T.; Angers, D.; Neilsen, D.; Hogue, E. Suitable orchard floor management strategies in organic apple orchards that augment soil organic matter and maintain tree performance. Plant Soil 2014, 378, 325–335. [Google Scholar] [CrossRef]
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Brito, O.R.; Telles, T.; Schnitzer, J.A.; Gaspar, G.G.; Guimarães, M.D.F. The influence of crop residues in vertical soil mobility of potassium. Semin. Ciênc. Agrár. 2014, 35, 3043. [Google Scholar] [CrossRef] [Green Version]
- Hougni, D.-G.J.M.; Schut, A.G.T.; Woittiez, L.S.; Vanlauwe, B.; Giller, K.E. How nutrient rich are decaying cocoa pod husks? The kinetics of nutrient leaching. Plant Soil 2021, 463, 155–170. [Google Scholar] [CrossRef]
- Kaushal, R.; Verma, K.; Chaturvedi, O.P.; Alam, N.M. Leaf Litter Decomposition and Nutrient Dynamics in Four Multipurpose Tree Species. Range Manag. Agrofor. 2012, 33, 20–27. [Google Scholar]
- Li, J.; Lu, J.; Li, X.; Ren, T.; Cong, R.; Zhou, L. Dynamics of Potassium Release and Adsorption on Rice Straw Residue. PLoS ONE 2014, 9, e90440. [Google Scholar] [CrossRef] [PubMed]
- Lizana, A.R.; Carbonell, R.; Gonzalez, P.; Ordoñez, R. N, P and K released by the field decomposition of residues of a pea-wheat-sunflower rotation. Nutr. Cycl. Agroecosyst. 2010, 87, 199–208. [Google Scholar] [CrossRef]
- Trofymow, J.A.; Moore, T.R.; Titus, B.; Prescott, C.; Morrison, I.; Siltanen, M.; Smith, S.M.; Fyles, J.; Wein, R.; Camiré, C.; et al. Rates of litter decomposition over 6 years in Canadian forests: Influence of litter quality and climate. Can. J. For. Res. 2002, 32, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.W.I.; Torn, M.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]
- Gross, C.D.; Harrison, R.B. The Case for Digging Deeper: Soil Organic Carbon Storage, Dynamics, and Controls in Our Changing World. Soil Syst. 2019, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Vida, C.; Ebonilla, N.; Vicente, A.E.; Cazorla, F.M. Microbial Profiling of a Suppressiveness-Induced Agricultural Soil Amended with Composted Almond Shells. Front. Microbiol. 2016, 7, 4. [Google Scholar] [CrossRef]
- Daly, A.B.; Jilling, A.; Bowles, T.M.; Buchkowski, R.W.; Frey, S.D.; Kallenbach, C.M.; Keiluweit, M.; Mooshammer, M.; Schimel, J.P.; Grandy, A.S. A holistic framework integrating plant-microbe-mineral regulation of soil bioavailable nitrogen. Biogeochemistry 2021, 154, 211–229. [Google Scholar] [CrossRef]
- Iremiren, G.O.; Ipinmoroti, R.R. Effect of Organic Fertilizer (Cocoa Pod Husk) on Okra and Maize Production under Okra/Maize Intercrop in Uhonmora, Edo State, Nigeria. Afr. J. Agric. Res. 2014, 9, 3789–3796. [Google Scholar]
- Molina-Murguía, J.F.F.; García-Calderón, N.E.; Martín, G.A. Cocoa Industrial Debris Composting in Soil and Earthworm Breeding. Compos. Sci. Util. 2009, 17, 197–204. [Google Scholar] [CrossRef]
- Bonanomi, G.; d’Ascoli, R.; Scotti, R.; Gaglione, S.; Caceres, M.G.; Sultana, S.; Scelza, R.; Rao, M.A.; Zoina, A. Soil quality recovery and crop yield enhancement by combined application of compost and wood to vegetables grown under plastic tunnels. Agric. Ecosyst. Environ. 2014, 192, 1–7. [Google Scholar] [CrossRef]
- Granatstein, D.; Andrews, P.; Groff, A. Productivity, economics, and fruit and soil quality of weed management systems in commercial organic orchards in Washington State, USA. Org. Agric. 2014, 4, 197–207. [Google Scholar] [CrossRef]
- TerAvest, D.; Smith, J.L.; Carpenter-Boggs, L.; Granatstein, D.; Hoagland, L.; Reganold, J.P. Soil Carbon Pools, Nitrogen Supply, and Tree Performance under Several Groundcovers and Compost Rates in a Newly Planted Apple Orchard. HortScience 2011, 46, 1687–1694. [Google Scholar] [CrossRef] [Green Version]
- Galanti, R.; Cho, A.; Ahmad, A.; Radovich, T. Soil Amendments and Soil Profiling Impact on Macadamia Growth and Yield Performance. HortScience 2019, 54, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Tahboub, M.B.; Lindemann, W.C.; Murray, L. Nutrient Availability in Soil Amended with Pecan Wood Chips. HortScience 2007, 42, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Nduka, B.A.; Adewale, D.B.; Akanbi, O.S.O.; Adejobi, K.B. Nursery Soil Amendments for Cashew Seedling Production: A Comparative Analysis of Coffee Husk and NPK. J. Agric. Sci. 2015, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, L.; Carpenter-Boggs, L.; Granatstein, D.; Mazzola, M.; Smith, J.; Peryea, F.; Reganold, J.P. Orchard floor management effects on nitrogen fertility and soil biological activity in a newly established organic apple orchard. Biol. Fertil. Soils 2008, 45, 11–18. [Google Scholar] [CrossRef]
- Idowu, O.J.; Sanogo, S.; Brewer, C.E. Short-Term Impacts of Pecan Waste By-Products on Soil Quality in Texturally Different Arid Soils. Commun. Soil Sci. Plant Anal. 2017, 48, 1781–1791. [Google Scholar] [CrossRef]
- Manzoni, S.; Jackson, R.B.; Trofymow, J.A.; Porporato, A. The Global Stoichiometry of Litter Nitrogen Mineralization. Science 2008, 321, 684–686. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 135–189. ISBN 9780123849052. [Google Scholar]
- Hartemink, A.E. Nutrient Stocks, Nutrient Cycling, and Soil Changes in Cocoa Ecosystems: A Review; Elsevier BV: Amsterdam, The Netherlands, 2005; Volume 86, pp. 227–253. [Google Scholar]
- Franchini, J.C.; Hoffmann-Campo, C.B.; Torres, E.; Miyazawa, M.; Pavan, M.A. Organic Composition of Green Manure During Growth and Its Effect on Cation Mobilization in an Acid Oxisol. Commun. Soil Sci. Plant Anal. 2003, 34, 2045–2058. [Google Scholar] [CrossRef]
- Singh, K.; Sanderson, T.; Field, D.; Fidelis, C.; Yinil, D. Soil security for developing and sustaining cocoa production in Papua New Guinea. Geoderma Reg. 2019, 17, e00212. [Google Scholar] [CrossRef]
- Zoca, S.M.; Penn, C.J.; Rosolem, C.A.; Alves, A.R.; Neto, L.O.; Martins, M.M. Coffee processing residues as a soil potassium amendment. Int. J. Recycl. Org. Waste Agric. 2014, 3, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Munongo, M.E.; Nkeng, G.E.; Njukeng, J.N. Production and Characterization of Compost Manure and Biochar from Cocoa Pod Husks. Int. J. Adv. Sci. Res. Manag. 2017, 2, 26–31. [Google Scholar]
- Adu-Dapaah, H.K.; Cobbina, J.; Asare, E.O.; Adu-Dapaah, H.K.; Cobbina, J.; Asare, E.O. Effect of cocoa pod ash on the growth of maize. J. Agric. Sci. 1994, 122, 31–33. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Calonego, J.C.; Foloni, J.S.S. Potassium Leaching from Millet Straw as Affected by Rainfall and Potassium Rates. Commun. Soil Sci. Plant Anal. 2005, 36, 1063–1074. [Google Scholar] [CrossRef]
- Cobo, J.G.; Barrios, E.; Kass, D.C.L.; Thomas, R.J. Decomposition and nutrient release by green manures in a tropical hillside agroecosystem. Plant Soil 2002, 240, 331–342. [Google Scholar] [CrossRef]
- Rahman, Z.A. Patterns of Decomposition and Nutrient Release by Fresh Gliricidia (Gliricidia sepium) Leaves in an Ultisol. Nutr. Cycl. Agroecosyst. 1999, 55, 269–277. [Google Scholar]
- Wu, J.; Guo, X.S.; Wang, Y.Q.; Xu, Z.Y.; Lu, J.W. Decomposition Characteristics of Rapeseed and Wheat Straw under Different Water Regimes and Straw Incorporating Models. J. Food Agric. Environ. 2011, 9, 572–577. [Google Scholar]
- Khalsa, S.D.S.; Hart, S.C.; Brown, P.H. Nutrient Dynamics of Surface-Applied Organic Matter Amendments on No-till Orchard Soil. Soil Use Manag. 2021. under review. [Google Scholar]
- Dhanya, B.; Viswanath, S.; Purushothaman, S. Decomposition and Nutrient Release Dynamics of Ficus benghalensis L. Litter in Traditional Agroforestry Systems of Karnataka, Southern India. ISRN For. 2013, 2013, 524679. [Google Scholar] [CrossRef] [Green Version]
- Mubarak, A.R.; Elbashir, A.A.; Elamin, L.A.; Daldoum, D.M.A.; Steffens, D.; Benckiser, G. Decomposition and Nutrient Release from Litter Fall in the Semi-arid Tropics of Sudan. Commun. Soil Sci. Plant Anal. 2008, 39, 2359–2377. [Google Scholar] [CrossRef]
- Bell, M.J.; Ransom, M.D.; Thompson, M.L.; Hinsinger, P.; Florence, A.M.; Moody, P.W.; Guppy, C.N. Considering Soil Potassium Pools with Dissimilar Plant Availability. In Improving Potassium Recommendations for Agricultural Crops; Murrell, T.S., Mikkelsen, R.L., Sulewski, G., Norton, R., Thompson, M.L., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 163–190. [Google Scholar]
- Havlin, J.L.; Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. Soil Fertility and Fertilizers, 8th ed.; Pearson: Uttar Pradesh, India, 2016. [Google Scholar]
- Pettygrove, S.; O’Geen, T.; Southard, R. Potassium Fixation and Its Significance for California Crop Production. Better Crop. Plant Food 2011, 95, 16–18. [Google Scholar]
- Murashkina, M.A.; Southard, R.J.; Pettygrove, G.S. Silt and fine sand fractions dominate K fixation in soils derived from granitic alluvium of the San Joaquin Valley, California. Geoderma 2007, 141, 283–293. [Google Scholar] [CrossRef]
- Olk, D.C.; Cassman, K.G. Reduction of Potassium Fixation by Two Humic Acid Fractions in Vermiculitic Soils. Soil Sci. Soc. Am. J. 1995, 59, 1250–1258. [Google Scholar] [CrossRef]
- Cassman, K.G.; Bryant, D.C.; Roberts, B.A. Comparison of soil test methods for predicting cotton response to soil and fertilizer potassium on potassium fixing soils. Commun. Soil Sci. Plant Anal. 1990, 21, 1727–1743. [Google Scholar] [CrossRef]
- Doungous, O.; Minyaka, E.; Longue, E.A.M.; Nkengafac, N.J. Potentials of cocoa pod husk-based compost on Phytophthora pod rot disease suppression, soil fertility, and Theobroma cacao L. growth. Environ. Sci. Pollut. Res. 2018, 25, 25327–25335. [Google Scholar] [CrossRef]
- Samuel, A.; Agbona, A. Effects of Cocoa Pod Husk Amendment on Soil and Leaf Chemical Composition and Growth of Cashew (Anacardium occidentale L.) Seedlings in the Nursery. Am. Eurasian J. Sustain. Agric. 2008, 2, 219–224. [Google Scholar]
- Carmo, D.L.D.; de Lima, L.B.; Silva, C.A. Soil Fertility and Electrical Conductivity Affected by Organic Waste Rates and Nutrient Inputs. Rev. Bras. Ciênc. Solo 2016, 40, 40. [Google Scholar] [CrossRef] [Green Version]
- Bittenbender, H.C.; Hue, N.V.; Fleming, K.; Brown, H. Sustainability of organic fertilization of macadamia with macadamia husk-manure compost. Commun. Soil Sci. Plant Anal. 1998, 29, 409–419. [Google Scholar] [CrossRef]
- Lobel, M.R.; Firth, D.J.; Johns, G.G. Effect of Mulch, Ca, and Mg on Growth, Yield, and Decline of Macadamia. Trop. Agric. 1994, 71, 170–175. [Google Scholar]
- Cayuela, M.L.; Bernal, M.P.; Roig, A. Composting Olive Mill Waste and Sheep Manure for Orchard Use. Compos. Sci. Util. 2004, 12, 130–136. [Google Scholar] [CrossRef]
- Altieri, R.; Esposito, A. Olive orchard amended with two experimental olive mill wastes mixtures: Effects on soil organic carbon, plant growth and yield. Bioresour. Technol. 2008, 99, 8390–8393. [Google Scholar] [CrossRef] [PubMed]
- Aranda, V.; Macci, C.; Peruzzi, E.; Masciandaro, G. Biochemical activity and chemical-structural properties of soil organic matter after 17 years of amendments with olive-mill pomace co-compost. J. Environ. Manag. 2015, 147, 278–285. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Psarras, G.; Moutsopoulou, M.; Stefanoudaki, E. Application of olive mill wastewater to a Cretan olive orchard: Effects on soil properties, plant performance and the environment. Agric. Ecosyst. Environ. 2010, 138, 293–298. [Google Scholar] [CrossRef]
- Guerrero, C.C.; de Brito, J.C.; Lapa, N.; Oliveira, J.F.S. Re-use of industrial orange wastes as organic fertilizers. Bioresour. Technol. 1995, 53, 43–51. [Google Scholar] [CrossRef]
- Meli, S.M.; Baglieri, A.; Porto, M.; Belligno, A.; Gennari, M. Chemical and Microbiological Aspects of Soil Amended with Citrus Pulp. J. Sustain. Agric. 2007, 30, 53–66. [Google Scholar] [CrossRef]
- Belligno, A.; di Leo, M.G.; Marchese, M.; Tuttobene, R. Effects of industrial orange wastes on soil characteristics and on growth and production of durum wheat. Agron. Sustain. Dev. 2005, 25, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, M.; Pavan, M.A.; Franchini, J.C. Evaluation of Plant Residues on the Mobility of Surface Applied Lime. Braz. Arch. Biol. Technol. 2002, 45, 251–256. [Google Scholar] [CrossRef]
- Amaral, A.S.; Anghinoni, I.; Deschamps, F.C. Residues of Cover Plants and Mobility of the Products of Lime Dissolution Applied to the Soil Surface. Braz. J. Soil Sci. 2004, 28, 115–123. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, S.; Al Mahmud, J.; Hossen; Masud, A.A.C.; Moumita; Fujita, M. Potassium: A Vital Regulator of Plant Responses and Tolerance to Abiotic Stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.L.; Rivero, M.C. Fertilization and Crop Residue Management in No-till Corn Production. In No-Till Farming; Nova Science Publishers, Inc.: New York, NY, USA, 2010; pp. 75–91. [Google Scholar]
- Ozenc, D.B.; Ozenc, N. Short-Term Effects of Hazelnut Husk Compost and Organic Amendment Applications on Clay Loam Soil. Compos. Sci. Util. 2008, 16, 192–199. [Google Scholar] [CrossRef]
- Moyin-Jesu, E.I. Use of plant residues for improving soil fertility, pod nutrients, root growth and pod weight of okra (Abelmoschus esculentum L). Bioresour. Technol. 2007, 98, 2057–2064. [Google Scholar] [CrossRef] [PubMed]
- Regni, L.; Nasini, L.; Ilarioni, L.; Brunori, A.; Massaccesi, L.; Agnelli, A.; Proietti, P. Long Term Amendment with Fresh and Composted Solid Olive Mill Waste on Olive Grove Affects Carbon Sequestration by Prunings, Fruits, and Soil. Front. Plant Sci. 2017, 7, 2042. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Xie, Y.; Wang, C.; Yue, J.; Yao, Y.; Li, X.; Liu, W.; Zhu, Y.; Guo, T. Effects of different soil conservation tillage approaches on soil nutrients, water use and wheat-maize yield in rainfed dry-land regions of North China. Eur. J. Agron. 2016, 81, 37–45. [Google Scholar] [CrossRef]
- Tahboub, M.B.; Lindemann, W.C.; Murray, L. Chemical and Physical Properties of Soil Amended with Pecan Wood Chips. HortScience 2008, 43, 891–896. [Google Scholar] [CrossRef]
- Moreno-Ramón, H.; Quizembe, S.J.; Ibanezasensio, S. Coffee husk mulch on soil erosion and runoff: Experiences under rainfall simulation experiment. Solid Earth 2014, 6, 851–862. [Google Scholar] [CrossRef] [Green Version]
- Laird, D.A.; Chang, C.-W. Long-term impacts of residue harvesting on soil quality. Soil Tillage Res. 2013, 134, 33–40. [Google Scholar] [CrossRef]
- Farzi, R.; Gholami, M.; Baninasab, B.; Gheysari, M. Evaluation of different mulch materials for reducing soil surface evaporation in semi-arid region. Soil Use Manag. 2017, 33, 120–128. [Google Scholar] [CrossRef]
- Karagoktas, M.; Uygur, V.; Aydoğdu, M.; Rastgeldi, I. The Effects of Pistachio Nut Hull Compost on Soil Fertility: A Comparative Study with Manure; Soil Science Society of Turkey & Federation of Eurasian Soil Science Societies: Side, Turkey, 2014. [Google Scholar]
- Morison, J.I.L.; Baker, N.R.; Mullineaux, P.M.; Davies, W.J. Improving water use in crop production. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 639–658. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; van Zwieten, L.; Ayres, M.; Morris, S. Macadamia Husk Compost Improves Soil Health in Sub-Tropical Horticulture; University of Sydney, Australia: Sydney, Australia, 2004. [Google Scholar]
- Zimmermann, S.; Braun, S.; Conedera, M.; Blaser, P. Macronutrient inputs by litterfall as opposed to atmospheric deposition into two contrasting chestnut forest stands in southern Switzerland. For. Ecol. Manag. 2002, 161, 289–302. [Google Scholar] [CrossRef]
- Hernández-Montoya, V.; Mendoza-Castillo, D.I.; Bonilla-Petriciolet, A.; Montes-Moran, M.A.; Pérez-Cruz, M.A. Role of the pericarp of Carya illinoinensis as biosorbent and as precursor of activated carbon for the removal of lead and acid blue 25 in aqueous solutions. J. Anal. Appl. Pyrolysis 2011, 92, 143–151. [Google Scholar] [CrossRef]
- Fernandez-Bayo, J.D.; Shea, E.A.; Parr, A.E.; Achmon, Y.; Stapleton, J.J.; van der Gheynst, J.S.; Hodson, A.K.; Simmons, C.W. Almond processing residues as a source of organic acid biopesticides during biosolarization. Waste Manag. 2020, 101, 74–82. [Google Scholar] [CrossRef]
- Diehl, R.C.; Miyazawa, M.; Takahashi, H.W. Water-Soluble Organic Compounds from Plant Residues and Their Effects on Soil Chemical Attributes. Braz. J. Soil Sci. 2008, 32, 2653–2659. [Google Scholar]
- Khalsa, S.D.S.; Almanza, C.A.; Brown, P.H.; Smart, D.R. Leaf litter C and N cycling from a deciduous permanent crop. Soil Sci. Plant Nutr. 2016, 62, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Forge, T.; Neilsen, G.; Neilsen, D.; O’Gorman, D.; Hogue, E.; Angers, D. Organic Orchard Soil Management Practices Affect Soil Biology and Organic Matter. Acta Hortic. 2015, 1076, 77–84. [Google Scholar] [CrossRef]
- Bonilla, N.; Cazorla, F.M.; Martínez-Alonso, M.; Hermoso, J.M.; González-Fernández, J.J.; Gaju, N.; Landa, B.B.; de Vicente, A. Organic amendments and land management affect bacterial community composition, diversity and biomass in avocado crop soils. Plant Soil 2012, 357, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Wall, D.H.; Nielsen, U.N.; Six, J. Soil biodiversity and human health. Nature 2015, 528, 69–76. [Google Scholar] [CrossRef]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2008, 105 (Suppl. 1), 11512–11519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haro, R.; Benito, B. The Role of Soil Fungi in K+ Plant Nutrition. Int. J. Mol. Sci. 2019, 20, 3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meena, V.S.; Maurya, B.R.; Verma, J.P. Does a rhizospheric microorganism enhance K+ availability in agricultural soils? Microbiol. Res. 2014, 169, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Benito, B.; Garciadeblás, B.; Fraile-Escanciano, A.; Rodriguez-Navarro, A. Potassium and sodium uptake systems in fungi. The transporter diversity of Magnaporthe oryzae. Fungal Genet. Biol. 2011, 48, 812–822. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.; Goss, E.M.; Havelaar, A.; van Diepeningen, A.D.; Finckh, M.; Morris, J.G. One Health—Cycling of diverse microbial communities as a connecting force for soil, plant, animal, human and ecosystem health. Sci. Total Environ. 2019, 664, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Diepeningen, A.D.; de Vos, O.J.; Korthals, G.W.; van Bruggen, A.H. Effects of organic versus conventional management on chemical and biological parameters in agricultural soils. Appl. Soil Ecol. 2006, 31, 120–135. [Google Scholar] [CrossRef]
- Urra, J.; Mijangos, I.; Lanzén, A.; Lloveras, J.; Garbisu, C. Effects of corn stover management on soil quality. Eur. J. Soil Biol. 2018, 88, 57–64. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. Soil microbiomes and climate change. Nat. Rev. Genet. 2020, 18, 35–46. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Wagg, C. Soil microbial diversity and agro-ecosystem functioning. Plant Soil 2013, 363, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Gámez-Virués, S.; Perovic, D.J.; Gossner, M.M.; Börschig, C.; Blüthgen, N.; de Jong, H.; Simons, N.K.; Klein, A.-M.; Krauss, J.; Maier, G.; et al. Landscape simplification filters species traits and drives biotic homogenization. Nat. Commun. 2015, 6, 8568. [Google Scholar] [CrossRef]
- Wood, S.A.; Karp, D.S.; De Clerck, F.; Kremen, C.; Naeem, S.; Palm, C.A. Functional traits in agriculture: Agrobiodiversity and ecosystem services. Trends Ecol. Evol. 2015, 30, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Kuzin, A.I.; Kashirskaya, N.Y.; Kochkina, A.M.; Kushner, A.V. Correction of Potassium Fertigation Rate of Apple Tree (Malus domestica Borkh.) in Central Russia during the Growing Season. Plants 2020, 9, 1366. [Google Scholar] [CrossRef]
- Zipori, I.; Yermiyahu, U.; Erel, R.; Presnov, E.; Faingold, I.; Ben-Gal, A.; Dag, A. The influence of irrigation level on olive tree nutritional status. Irrig. Sci. 2015, 33, 277–287. [Google Scholar] [CrossRef]
- Muhammad, S.; Sanden, B.L.; Lampinen, B.D.; Smart, D.R.; Saa, S.; Shackel, K.A.; Brown, P.H. Nutrient Storage in the Perennial Organs of Deciduous Trees and Remobilization in Spring—A Study in Almond (Prunus dulcis) (Mill.) D.A. Webb. Front. Plant Sci. 2020, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Grzebisz, W.; Gransee, A.; Szczepaniak, W.; Diatta, J. The effects of potassium fertilization on water-use efficiency in crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 355–374. [Google Scholar] [CrossRef]
- Martineau, E.; Domec, J.-C.; Bosc, A.; Denoroy, P.; Fandiño, V.A.; Lavres, J.; Jordan-Meille, L. The effects of potassium nutrition on water use in field-grown maize (Zea mays L.). Environ. Exp. Bot. 2017, 134, 62–71. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.S.; Berkowitz, G.; Pier, P. Maintenance of Photosynthesis at Low Leaf Water Potential in Wheat: Role of Potassium Status and Irrigation History. Plant Physiol. 1989, 89, 1358–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, P.H.; Weinbaum, S.A.; DeJong, T.M. Reassessment of Potassium Critical Values. In Years of Discovery (1972–2003); Almond Board of California: Modesto, CA, USA, 2000. [Google Scholar]
- Fidelis, C.; Rao, B.K.R. Enriched cocoa pod composts and their fertilizing effects on hybrid cocoa seedlings. Int. J. Recycl. Org. Waste Agric. 2017, 6, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Laclau, J.-P.; Bouillet, J.-P.; Ranger, J. Dynamics of biomass and nutrient accumulation in a clonal plantation of Eucalyptus in Congo. For. Ecol. Manag. 2000, 128, 181–196. [Google Scholar] [CrossRef]
- Kone, K.; Akueson, K.; Norval, G. On the Production of Potassium Carbonate from Cocoa Pod Husks. Recycling 2020, 5, 23. [Google Scholar] [CrossRef]
- Oyewole, O.; Omueti, J.; Rotimi, R. Growth of Cocoa (Theobroma cacao L.) Seedlings on Old Cocoa Soils Amended with Organic and Inorganic Fertilizers. Afr. J. Agric. Res. 2012, 7, 3604–3608. [Google Scholar] [CrossRef]
- Nasini, L.; Gigliotti, G.; Balduccini, M.A.; Federici, E.; Cenci, G.; Proietti, P. Effect of solid olive-mill waste amendment on soil fertility and olive (Olea europaea L.) tree activity. Agric. Ecosyst. Environ. 2013, 164, 292–297. [Google Scholar] [CrossRef]
- Granatstein, D.; Mullinix, K. Mulching Options for Northwest Organic and Conventional Orchards. HortScience 2008, 43, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Neilsen, G.H.; Hogue, E.J.; Forge, T.; Neilsen, D. Mulches and Biosolids Affect Vigor, Yield and Leaf Nutrition of Fertigated High Density Apple. HortScience 2003, 38, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Neilsen, G.H.; Hogue, E.J.; Forge, T.; Neilsen, D. Surface application of mulches and biosolids affect orchard soil properties after 7 years. Can. J. Soil Sci. 2003, 83, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Lu, X. A meta-analysis of the effects of crop residue return on crop yields and water use efficiency. PLoS ONE 2020, 15, e0231740. [Google Scholar] [CrossRef]
- Khalsa, S.D.S.; Smart, D.R.; Muhammad, S.; Armstrong, C.M.; Sanden, B.L.; Houlton, B.Z.; Brown, P.H. Intensive fertilizer use increases orchard N cycling and lowers net global warming potential. Sci. Total Environ. 2020, 722, 137889. [Google Scholar] [CrossRef]
- Valenzuela-Solano, C.; Crohn, D.M.; Downer, J.A. Nitrogen mineralization from eucalyptus yardwaste mulch applied to young avocado trees. Biol. Fertil. Soils 2004, 41, 38–45. [Google Scholar] [CrossRef]
- Van der Vossen, H.A.M. A Critical Analysis of the Agronomic and Economic Sustainability of Organic Coffee Production. Exp. Agric. 2005, 41, 449–473. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.T.; Nguyen, N.H.; Yen, P.N.D.; Lam, T.D.; Le, N.T.T. Proposed Techniques to Supplement the Loss in Nutrient Cycling for Replanted Coffee Plantations in Vietnam. Agronomy 2020, 10, 905. [Google Scholar] [CrossRef]
- Chen, C.; Khir, R.; Shen, Y.; Wu, X.; Zhang, R.; Cao, X.; Niederholzer, F.; Pan, Z. Energy consumption and product quality of off-ground harvested almonds under hot air column drying. LWT 2021, 138, 110768. [Google Scholar] [CrossRef]
- Janzen, H.H. The soil carbon dilemma: Shall we hoard it or use it? Soil Biol. Biochem. 2006, 38, 419–424. [Google Scholar] [CrossRef]
- Glover, J.D.; Reganold, J.P.; Andrews, P.K. Systematic method for rating soil quality of conventional, organic, and integrated apple orchards in Washington State. Agric. Ecosyst. Environ. 2000, 80, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.E.; Edson, C.E.; Bird, G.W.; Whalon, M.E.; Willson, T.C.; Harwood, R.R.; Kizilkaya, K.; Nugent, J.E.; Klein, W.; Middleton, A.; et al. Orchard Floor and Nitrogen Management Influences Soil and Water Quality and Tart Cherry Yields. J. Am. Soc. Hortic. Sci. 2003, 128, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Khalsa, S.D.S.; Brown, P.H. Grower Analysis of Organic Matter Amendments in California Orchards. J. Environ. Qual. 2017, 46, 649–658. [Google Scholar] [CrossRef]
- Lubell, M.; Niles, M.; Hoffman, M. Extension 3.0: Managing Agricultural Knowledge Systems in the Network Age. Soc. Nat. Resour. 2014, 27, 1089–1103. [Google Scholar] [CrossRef]
- Walton, M.; Hall, J.; Guest, D.; Butubu, J.; Vinning, G.; Black, K.; Beardsley, J. Applying one health methods to improve cocoa production in Bougainville: A case study. One Health 2020, 10, 100143. [Google Scholar] [CrossRef]
- Agbeniyi, S.O.; Oluyole, K.A.; Ogunlade, M.O. Impact of Cocoa Pod Husk Fertilizer on Cocoa Production in Nigeria. World J. Agric. Sci. 2011, 7, 113–116. [Google Scholar]
- Lu, F.; Garcia, J.R.; van Damme, I.V.; Westwood, N.J.; Shaw, L.; Robinson, J.S.; Warren, G.; Chatzifragkou, A.; Mason, S.M.; Gomez, L.; et al. Valorisation strategies for cocoa pod husk and its fractions. Curr. Opin. Green Sustain. Chem. 2018, 14, 80–88. [Google Scholar] [CrossRef]
- Shemekite, F.; Gómez-Brandón, M.; Franke-Whittle, I.H.; Praehauser, B.; Insam, H.; Assefa, F. Coffee husk composting: An investigation of the process using molecular and non-molecular tools. Waste Manag. 2014, 34, 642–652. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, W.A.; Nogueira, F.N.; Devens, D.C. Temperature and pH control in composting of coffee and agricultural wastes. Water Sci. Technol. 1999, 40, 113–119. [Google Scholar] [CrossRef]
- Sathianarayanan, A.; Khan, A. An Eco-Biological Approach for Resource Recycling and Pathogen (Rhizoctoniae solani Kuhn.). Suppression 2008, 2, 36–39. [Google Scholar]
- Winter, E.; Marton, S.M.R.R.; Baumgart, L.; Curran, M.; Stolze, M.; Schader, C. Evaluating the Sustainability Performance of Typical Conventional and Certified Coffee Production Systems in Brazil and Ethiopia Based on Expert Judgements. Front. Sustain. Food Syst. 2020, 4, 49. [Google Scholar] [CrossRef]
- Le, Q.V.; Jovanovic, G.; Le, D.-T.; Cowal, S. Understanding the Perceptions of Sustainable Coffee Production: A Case Study of the K’Ho Ethnic Minority in a Small Village in Lâm Đồng Province of Vietnam. Sustainability 2020, 12, 1010. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrews, E.M.; Kassama, S.; Smith, E.E.; Brown, P.H.; Khalsa, S.D.S. A Review of Potassium-Rich Crop Residues Used as Organic Matter Amendments in Tree Crop Agroecosystems. Agriculture 2021, 11, 580. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070580
Andrews EM, Kassama S, Smith EE, Brown PH, Khalsa SDS. A Review of Potassium-Rich Crop Residues Used as Organic Matter Amendments in Tree Crop Agroecosystems. Agriculture. 2021; 11(7):580. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070580
Chicago/Turabian StyleAndrews, Ellie M., Sire Kassama, Evie E. Smith, Patrick H. Brown, and Sat Darshan S. Khalsa. 2021. "A Review of Potassium-Rich Crop Residues Used as Organic Matter Amendments in Tree Crop Agroecosystems" Agriculture 11, no. 7: 580. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070580