
Breeding for Nutritional and Organoleptic Quality in Vegetable Crops: The Case of Tomato and Cauliflower


1
Council for Agricultural Research and Economics (CREA), Research Centre for Vegetable and Ornamental Crops (OF), 63077 Monsampolo del Tronto, Italy
2
Council for Agricultural Research and Economics (CREA), Research Centre for Vegetable and Ornamental Crops (OF), 84098 Pontecagnano Faiano, Italy
*
Author to whom correspondence should be addressed.
†
Retired.
Agriculture 2021, 11(7), 606; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070606
Submission received: 29 May 2021
/
Revised: 22 June 2021
/
Accepted: 25 June 2021
/
Published: 29 June 2021
(This article belongs to the Special Issue Breeding and Genetics of Horticultural Crops)
Abstract
:Due to novel and more demanding consumers’ requirements, breeding of vegetable crops confronts new challenges to improve the nutritional level and overall appearance of produce. Such objectives are not easy to achieve considering the complex genetic and physiological bases. Overtime, plant breeders relied on a number of technologies and methods to achieve ever changing targets. F1 hybrid seed production allowed the exploitation of heterosis and facilitated the combination of resistance and other useful genes in a uniform outperforming variety. Mutagenesis and tissue culture techniques permitted to induce novel variation, overcome crossing barriers, and speed up the achievement of true-breeding lines. Marker-assisted selection was one of the milestones in fastening selection, starting from the early ’90s in almost all seed companies. Only recently, however, are novel omics tools and genome editing being used as cutting-edge techniques to face old and new challenges in vegetable crops, with the potential to increase the qualitative value of crop cultivation and solve malnutrition in 10 billion people over the next 30 years. In this manuscript, the evolution of breeding approaches in vegetable crops for quality is reviewed, reporting case studies in tomato (Solanum lycopersicum L.) and cauliflower (Brassica oleracea var. botrytis L.) as model systems for fleshy fruit and floral edible parts, respectively.
1. Introduction
Vegetable quality improvement is a global requirement due to current evolution of consumers’ demands. In particular, producing vegetables with enhanced nutritional and organoleptic quality is one of the most challenging targets for breeding, facing with climatic changes and the needs for a more efficient production system [1]. High throughput metabolomic, transcriptomic and genomic advances represent useful tools to identify genetic architecture and biochemical pathways and also to predict breeding values for selection and deployment [2].
Among vegetables, tomato and cauliflower, cultivated throughout the world, are important dietary sources of phytochemical and bioactive compounds and represent model systems for fleshy fruit and floral edible parts, respectively [3,4]. While advanced genomic and genetic transformation resources are available in tomato for next-generation precise breeding, the availability of such resources in cauliflower is lower and hence breeding methods are essentially conventional, resulting in slow progress, despite recent interesting novel products [5].
In this manuscript, the evolution of breeding in tomato and cauliflower is reviewed, focusing on nutritional and organoleptic quality, flavour, and taste.
2. Breeding for Old and New Challenges in Vegetable Crops
Modern breeding needs to address global challenges including climate change, increasing yield and quality to secure a global food supply for a growing world population and participate in maximising the resources of farming systems [1,6]. Current tools in multiomic approaches are a fundamental support for plant breeding to match crop metabolism genetics and further improve organoleptic and nutraceutical quality in new varieties [1].
As reported in Figure 1, starting from the discovery of the laws of genetics by Gregor Mendel in 1865, plant breeders improved breeding methods that exploit genetic diversity in recombining and selecting the most performing plants. At the beginning of last century, Wilhelm Johansen in 1903 elaborated the pure line theory of heredity through the distinction between genotype (heritable variation in plants) and phenotype (non-heritable variation in plants) [7]. The first hybrid breeding schemes were introduced during 1920s to develop varieties, and among them, the investigations of Nazareno Strampelli have been a milestone in food production, mainly for cereal crops [8]. In a pioneering publication in 1907, Strampelli evidenced the laws of inheritance applied to 27 traits of interest in wheat, and that investigation led to the development of elite lines according to “innovative” hybridization schemes for the constitution of new high yield and resistant varieties [9]. In order to increase genetic diversity, starting from the ‘30s, radiation has been largely used to induce random mutations in plant genomes followed by selection for new desirable traits. Tissue culture techniques were initially introduced since 1960 to produce clones and fasten wide crosses through embryo rescue technologies [6]. Furthermore, the use of microspore cultures allowed the productions of double haploid plants, saving several steps in backcross breeding schemes to achieve homozygosity [6,10]. In the 1940s, in corn, Barbara Mc Clintock, Nobel prize winner, identified the transposons, or transposable elements, as regulators of gene expression and defined the basic concepts of epigenetics [11]. In 1977, van Montagu and Schell used for the first time Agrobacterium tumefaciens as a vector to create transgenic plants and in 1983 during a challenging symposium held in Miami on “Genetic manipulation of Plants”, Mary-Dell Chilton, Jeff Schell, and Bob Horsch reported successful regeneration of plant cells with an antibiotic resistance gene [12]. As another main step, in the 1980s, Botstein and co-authors introduced restriction fragment length polymorphism (RFLP) for linkage mapping in humans. RFLP was adopted as the first DNA-based molecular marker system, and it was used to develop marker linkage maps and map several traits of interest in many crops [13]. Subsequently, other markers have been developed, such as random amplified polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP), simple sequence repeat (SSR), etc. The high availability and reproducibility and the low cost of new markers represented a big advantage in plant breeding. Nowadays, molecular markers are widely used for tomato breeding that is not only an important crop but also represents a model in vegetables. More than 40 genes that confer resistance to the main tomato pathogens have been mapped, cloned, and/or sequenced [14]. These maps allowed to “pyramid” resistance genes through MAS by which multiple genes for desirable traits can be introduced into one genotype. Furthermore, in the last two decades, new technologies for genome sequencing have been developed and integrated into traditional breeding pipelines [2]. Genomics rapid cycling has fundamental importance for the prediction of breeding value with the potential of reducing cycle time (rapid cycling) and increasing accuracy [2]. Genomic information prompted also the development of novel tools for genome editing, such as oligonucleotide mutagenesis (ODM) or nuclease-mediated site-specific genome modifications, which provide the possibility to increase genetic variation as it was done in the past for some crops by radiation or chemical mutagenesis for knockout function studies but also to precisely introduce mutations in genes to silence or improve their function [6]. Further, in comparison with established transgenic technology, gene transfer by genome editing methodologies allows the precise integration of the transgene/cisgene and a more predictable expression [15]. Above all the site-specific genome modifications, CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats/CRISPR) associated endonuclease 9) is the most representative in genome editing, and it was reported for the first time in 2013 for plants as reviewed by Bortesi and Fischer (2015) [16].
Nonetheless, in spite of the availability of new molecular tools to improve specific traits, plant breeding still maintains also a phenotypic approach testing in several environments by considering all the relevant agronomic characteristics: yield, resistance/tolerance to biotic and abiotic stress, and quality. Due to modern consumers’ demands, plant breeders are now more focused on improving crop quality traits, such as flavour, or nutritional value in some important commercial crops, such as tomato [17]. Most crop agronomic and quality characteristics (mainly primary and secondary metabolites) are controlled by multiple quantitative trait loci (QTL), and their expression is also affected by environmental conditions. In order to identify the position of relevant loci, linkage mapping and genome-wide association (GWAS) are frequently applied. After having identified the association between genetic and phenotypic variation for the traits of interest, marker-assisted selection is frequently used, when possible, in breeding programs, in order to develop high-quality crops [1].
In particular, the extensive use of next generation sequencing platforms (NGSP) has allowed high-density genotyping with SNPs and the application of GWAS to fine-map loci involved in plant metabolic complexity. In particular, GWAS combined with metabolomic platforms (mGWAS) has allowed a consistent screening of several accessions for primary metabolic content in Arabidopsis and, subsequently, also in tomato [18,19]. The identified QTLs controlling metabolites are not randomly distributed on the chromosomes, but QTL ‘hotspots’ are frequently observed, suggesting pleiotropic effects and also heterogeneous genetic control based on the groups of sub-populations [1]. Furthermore, Chen and co-authors, demonstrated a tissue-specific metabolic variation for metabolite production and accumulation in cereals that could be referred as differential allelic expression [20]. Due to the complex genetic control underlying metabolite production in plant tissues, integrative approaches, combining metabolomics and functional genomics, could improve the constitution of new improved varieties.
Notwithstanding the availability of NGSP, conventional breeding based on crosses, phenotypic selection and breeding schemes (e.g., pedigree method, hybridization, mass selection, and backcrossing) is still used in public as well as in private breeding programs to generate improved varieties with desirable traits [21]. In particular for not genome-sequenced crops and specific markets (e.g., organic agriculture), the exploitation of genetic variations among cultivars, landraces, wild species, and the variability generated by inter-intraspecific cross is the basis to generate prebreeding lines to be developed as parent lines.
Recently, in order to limit derogations to the use of conventionally produced seeds in organic farming, attention has been directed to breed varieties adapted to such systems and characterized by appreciable yield and quality. Two EU H2020 projects (Liveseed, https://www.liveseed.eu/ (accessed on 22 June 2021), and Bresov, https://bresov.eu/ (accessed on 22 June 2021)) have been funded to select highly resilient varieties and identify genes/QTL underpinning relevant traits linked to resilience and low input farming in tomato, Brassicas, and other vegetables. In the framework of those projects, traditional tomato germoplasm resources characterized by high resilience (e.g., Spanish and Italian ‘da serbo’ Landraces) as well as accessions of underexploited genetic reservoirs and derived populations (the wild species Solanum pimpinellifolium and Solanum cheesmaniae and the weedy S. lycopersicum var. cerasiforme) have been deeply genotyped for further use in organic breeding programs [22,23,24,25,26].
3. Botanical Aspects of Tomato and Cauliflower
Notably, tomato (Solanum lycopersicum L.) is one of the most consumed vegetables worldwide and the area under cultivation is increasing daily, as it is a relatively short duration crop, gives a high yield, and is economically attractive. Total production is around 180 million tons globally and the main production regions are located in temperate zones, close to the 40th parallels North and South (http://www.fao.org/faostat (accessed on 28 April 2021)). S. lycopersicum L. can be further divided into two botanical types: large-fruited tomatoes S. lycopersicum var. lycopersicum (SLL) and cherry-sized early domesticates S. lycopersicum var. cerasiforme (SLC) (Figure 2) [27]. From a vegetative point of view, tomatoes are distinguished into three different types of plants: (i) tall or indeterminate type, (ii) semi-bush or semi-indeterminate type, (iii) bush or determinate type. Due to its economic importance and short growing cycle, a reference tomato genome (processing tomato variety Heinz 1706) has been sequenced and released [28].
Thanks to the superior performance of hybrids, due to heterosis, commercial varieties are mainly F1 hybrids that can be produced by: (i) hand emasculation and hand pollination, (ii) using male sterility and hand pollination, (iii) using male sterility and natural pollination [29].
Cauliflower (Brassica oleracea var. botrytis L.) is an important vegetable grown on approximatively 9 million Ha worldwide with a production of around 27 million tons in 2020 combined with broccoli (www.fao.org) (accessed on 28 April 2021). Cauliflower belongs to the Brassica genus which contains 37 species of which 6 interrelated ones gained worldwide economic importance. The various forms of B. oleracea include vegetable and forage crops: cabbage, broccoli, kale, brussels sprouts, cauliflower, and others. The Cauliflower group is divided into subgroups based on thermic and photoperiodic requirements necessary to produce the curd (e.g., tropical, subtropical, and winter types) or, more commonly, on the colour of the edible organ (namely head or flesh): white, green (e.g., Romanesco type), light yellow, purple, and more recently also orange cauliflowers (Figure 3) [30,31]. Nevertheless, as anticipated by Schulz in 1919, many B. oleracea forms come from Mediterranean wild ecotypes/landraces [32]; in particular, cauliflower domestication derived from a heading Calabrese broccoli through an intermediate Sicilian type in southern Italy [33,34].
Cauliflower hybrids also show commercial advantages thanks to heterosis, mainly in terms of uniform maturity, total yield, better curd quality for compactness and colour, and resistance to pests and diseases. There are two pollination mechanisms for hybrid seed production in cauliflower: self-incompatibility (SI) and cytoplasmic male sterility, largely used in the commercial hybrid seed production (improved Ogura based CMS lines) [35,36].
4. Quality in Tomato and Cauliflower
The quality of vegetables is defined as a “dynamic composite of the physicochemical properties pertaining to horticultural commodities and consumer perception” [37]. Therefore, the concept of quality is strictly related to the specific vegetable and its consumption, and it depends on genetic, environmental conditions (open field, greenhouse, glasshouse conditions), and crop management (low input system production, pesticide residue free, etc.).
Dorais and co-authors (2001) properly defined tomato fruit quality for fresh consumption as the sum of “appearance (colour, size, shape, absence of physiological disorders, and decay), firmness, texture, dry matter content and organoleptic (flavour) and nutraceutical (health benefit) properties [38]. Regarding firmness, breeders used in their programs tomato ripening mutants in order to increase shelf-life. Among them, the most important are ripening inhibitor (rin) and non-ripening (nor) that are linked to ripening alteration related to ethylene insensitivity [39]. Due to pleiotropic effects on colour and flavour, rin and nor genes are introduced only in heterozygous forms in the candidate new hybrid, resulting in extended shelf-life [40].
The organoleptic quality of tomato is mainly attributed to aroma volatiles, sugar and acid content, while nutraceutical quality is defined by mineral, vitamin, carotenoid, and flavonoid content [38]. Regarding tomato flavour, this attribute is a complex trait influenced by sugars, acids, and volatiles. Ripe tomato fruits produce hundreds of volatile compounds that are derived from essential nutrients: essential fatty acids, amino acids, and coloured carotenoids [41]. Phenotypic selection for volatile compounds is almost impossible because their concentrations range from micrograms per gram fresh weight for hexanal to nanograms per gram for β-damascenone [41]. Each volatile compound contributes to the global fruit flavour based on interactions among components and not according to additive effect [42]. Furthermore, some volatiles (esters) are negatively related to consumer acceptance and other ones can improve also sweetness perception [41].
In cauliflower, the definition of quality of the edible part is related to its biology. Indeed, while tomato fruit is a berry, cauliflower curds are undeveloped pre-floral fleshy apical meristem (flower buds) including also a shoot system with short internodes, branch apices, and bracts [34]. Cauliflower is the only variety of B. oleracea presenting a hypertrophic structure of the immature inflorescence termed curd [43]. Quality in cauliflower is related to attributes related to the external inflorescence (colour, firmness, tissue turgidity, absence of undesirable disorders) and chemical attributes such as glucosinolates, vitamin C, polyphenols, and carotenoids [44]. Nowadays, consumers are becoming extremely health-conscious regarding the system adopted to produce vegetables and the quality of the edible part, focusing on phytochemicals and bioactive compounds. In cauliflower, the development of new candidate hybrids has to include yield, adaptability to environmental conditions, quality, and nutritional value [45]. Heterosis has been studied in cauliflower in relation to commercial and vegetative traits [4,46] and, more recently, also to antioxidant pigments [47].
The main pigments are made up of several phytochemicals such as β-carotene (orange), lutein (yellowish-green), chlorophyll (green), and anthocyanin (purple) [47]. In the curd of the purple cauliflower variety ‘Graffiti’, it has been reported that the biosynthesis of anthocyanin is regulated by a single semi-dominant gene Pr [48,49]. Carotenoids are secondary plant compounds, and they constitute lipid soluble yellow, orange, and red pigments, with evidence on their positive action to prevent cancer and other chronic diseases [50]. Singh and co-authors reported a lack in the knowledge of heterosis and gene action for these molecules in cauliflower [47].
The first orange curd was reported by Crisp in a commercial white curded autumn crop variety named Extra Early Snowball commercially grown in Bradford, Ontario, Canada [51]. The orange colour is controlled by the Or semidominant single-locus gene mutation, while “or” is the recessive allelic form responsible for white colour [52]. The Or gene is responsible for high β-carotene accumulation in plastids of the edible curd and shoot meristems [53].
Lycopene is a hydrocarbon carotenoid, responsible for red colour pigmentation due to conjugated double bonds. In vegetables, lycopene is abundant in all the red-coloured fruits and vegetables, such as tomato and watermelon, and interestingly, it has been found also in white cauliflower [4]. Nowadays, there are no further details on the genetics of lycopene accumulation in cauliflower (inheritance and heritability), but new insights will be useful for breeding. Anthocyanins are responsible for blue, purple, and black colours in vegetables. These pigments have been revealed as an excellent source of antioxidants [54]. In cauliflower, anthocyanins have been reported in several studies, such as the investigations of Volden and co-authors on white and purple cauliflowers and those of Lo Scalzo and co-workers on violet cauliflower [55,56]. Ascorbic acid (vitamin C) is compartmentalised into chloroplasts and other cell compartments, including the cell wall [4]. Ascorbate is commonly present in white and purple cauliflower, but freezer storage can negatively affect the content of this pigment with a significative reduction of 20% compared to the initial composition in fresh curds at harvest [55].
For breeding purposes, the development of hybrids with high content in phytochemicals and bioactive compounds (vitamins and other antioxidants pigments) will be of great interest for the evolution of consumers’ demand oriented to nutraceutical vegetables. For example, in tomato, monogenic recessive high pigment (hp-1 and hp-2) and dark green (dg) mutations are used to develop new varieties with high phytochemical content as investigated by Tommonaro and co-authors (2012) on dark tomatoes [57]. “Dark skin tomatoes” is a general term including black as well as purple tomatoes obtained with mutations in carotenoid and/or flavonoid biosynthesis or chlorophill catabolism [58]. An example of black tomato is the purple line called “Sun Black” (Aft Aft/atv atv) derived from S. chilense and S. cheesmaniae that have been used to introduce the alleles Anthocyanin fruit (Aft) and atroviolacea (atv) responsible for anthocyanin biosynthesis [59]. This genotype has been reported to present anthocyanins in the peel and other bioactive compounds in the whole fruit, such as chlorogenic acid (the most abundant phenolic acid), the flavonol rutin other than carotenoids, polyphenols, and vitamin C [58].
In the San Marzano (SM) landrace, single and combined mutations for colour have been introduced in several introgression lines [60]. SM mutants for yellow flesh (r), green flesh (gf), and colourless fruit epidermis (y) generated yellow, brown and pink fruit variants were investigated for volatile compounds, non-polar and polar metabolites [60]. The authors found a decrease in carotenoids and flavonoids for r and y mutants, but xanthophyll, tocopherol, and amino acids increased, whereas in gf mutants, the vitamin E and sugar content were enhanced [60].
Nevertheless, it is also necessary to deeply investigate the genetic combining ability and extent of heterosis for these traits.
5. Evolution of Breeding for Nutritional and Organoleptic Quality
Investigations on the genes involved in the metabolism and detailed studies of the metabolic pathways are fundamental for plant breeders to develop varieties with improved quality. Since the publication of the Arabidopsis thaliana genome in 2000, next-generation sequencing (NGS) has been applied on several crops, other than model plants. NGS technologies evolved in their sequencing approach from BAC-by-BAC-based sequencing methodologies to whole-genome shotgun with longer read lengths and higher quality of genome sequences [61].
After the release of the inbred tomato cultivar Heinz 1706 as reference genome, several cultivated and wild tomato accessions (e.g., S. pimpinellifolium, S. cheesmaniae, S. galapagense) have been resequenced and compared to the reference [28,62,63,64,65,66]. The aim was to create a pan-genome comprising genomes of cultivated tomatoes and their wild ancestor and progenitors [66], in order to have a more complete characterization of tomato gene function and potential. In these studies, genomic variation of the tested accessions compared to the reference was estimated through mapping of short reads. Several (unknown) loci and highly divergent alleles have been identified, including 4873 additional genes that were absent in the reference genome. Among the (unknown) genes, for example, the tomato lipoxygenase C TomLOXC promoter was identified by Gao and co-workers. TomLoxC has been previously selected against during domestication and conventional breeding. TomLoxC contributes to tomato quality in terms of flavour by catalysing the synthesis of lipid-derived C5 and C6 volatiles and also in terms of nutraceutical value being involved in apocarotenoid (carotenoid derived compounds) production [66]. In particular, genotypes heterozygous for the TomLoxC promoter showed the highest expression in orange-stage fruit and QTL mapping identified TomLoxC as a key gene controlling the content of flavour-associated lipid- and carotenoid-derived volatiles. Furthermore, transgenic tomato fruit, with reduced TomLoxC expression, showed unknown non-enzymatic apocarotenoid pathway [66].
In detail, two flavour molecules are geranylacetone and 6-methyl-5-hepten-2-one (MHO) belonging to the family of apocarotenoids. In GWAS study, Tieman and co-workers [64] identified one associated locus for MHO, four loci for geranylacetone and two loci for both compounds. Comparing the frequency of alleles in wild species and modern accessions, the authors found that allelic combinations for geranylacetone were lost along domestication while two alleles for MHO content were positively selected. In other terms, over cycles of selection, breeders increased MHO levels by enhancing lycopene content and the MHO precursor as well [1,64].
The combination of metabolomics, transcriptomics, and mGWAS is a comprehensive approach to understand the fruit metabolic, and quality changes occurred during domestication and plant breeding [1,67,68]. The QTL approach in biparental populations revealed the genetic architecture of traits related to fruit quality. Nonetheless, QTL mapping does not consider the entire genetic diversity present in germplasm collections; therefore, the GWAS strategy has been used in several instances. Indeed, GWAS allows the screening of a wide range of accessions in order to understand the inheritance of important fruit metabolic traits [1,19,64,68]. GWAS has been performed in hundreds of tomato genotypes to identify genes involved in primary metabolism which includes sugars, acids, and other compounds of tomato flavour. For example, Sauvage and co-authors (2014) identified 44 loci for 19 primary metabolic traits [19].
One of the detected loci linked to soluble solid content is lin5 gene, which encodes for a cell wall invertase that was previously identified in a QTL study for fruit sugar content in silenced tomato plants [69]. Moreover, Tieman and co-authors identified, by means of GWAS, two loci on chromosomes 9 and 11, respectively, that control glucose and fructose contents, respectively. The locus on chromosome 9 corresponded to the lin5 gene linked to an extracellular invertase as emerged in QTL mapping of an F2 population, derived from a cross between a big sized flavourful variety and a small sized flavourless inbred line [64]. Interestingly, the same authors also found a SNP for lin5 associated to an amino acid change in the protein sequence at the position 366 as revealed after molecular investigations and confirmed with transgenic tomato plants overexpressing lin5 variants [64]. As suggested by Pott and co-workers (2021) [1], breeders, selecting for fruit size, have selected the combination of alleles on chromosome 9 and 11 associated with the lowest sugar content.
As previously mentioned, breeding activities have been oriented also to improve colour and increase antioxidant content, introducing, for example, the alleles responsible for anthocyanin biosynthesis in Sun Black tomato. This genotype is a double mutant line for anthocyanins (Aft Aft/atv atv) obtained after 20 years of breeding activities in which wild tomatoes species have been used in interspecific breeding program as source of Aft and atv genes (accessions LA 1996 for Aft and LA0797 for atv) [59]. In detail, Aft is a dominant gene on chromosome 10 encoding an R2R3-MYB transcription factor responsible for the activation of anthocyanin biosynthesis (absent in WT tomatoes) in presence of light. On the other hand, atv, located on chromosome 7, is a recessive mutation of Atv, which encodes an R3-MYB transcription factor acting as repressor of anthocyanin biosynthesis [58,70].
In tomato, other important qualitative traits are fruit shape and size, being positively selected for larger fruited types since domestication and variably shaped fruit according to the consumers’ requirements. A milestone review is the publication of Tanksley for reporting genetic and molecular bases of fruit size and shape variation in tomato [71]. Linked to domestication, the key loci controlling fruit size are: fw1.1, fw2.2, fw3.1, and fw4.1 which positively affected the selection for size from wild berries to the modern tomatoes. The above-mentioned loci can indirectly affect fruit shape under the direct genetic control of three other major loci with a minimal effect on fruit size. The loci for shape are located on chromosome 8 (sun and fs8.1) and on chromosome 2 (ovate) [71]. More recently, based on segregating populations obtained by crossing a plum tomato breeding line (NC 30P) and a grape tomato breeding line (NC-22L-1), QTL analysis and linkage mapping revealed the existence of a QTL on chromosome 10 regulating triangle fruit shape and two possible QTLs on chromosome 12 controlling obovoid shape [72].
Parthenocarpy is the fruit set and growth without fertilization and is a common phenomenon in Angiosperms in sub-optimal growing conditions (e.g., low temperature, low solar radiation, high relative humidity). In tomato, parthenocarpy leads to the absence or low number of seeds, which is a desirable trait for sauce production, and it has been demonstrated to affect fruit quality in terms of flavour (pH, total solid content) [73]. A parthenocarpic tomato mutant line is pat2 that has been introduced in the Severianin variety derived from the interspecific cross S. lycopersicum × S. peruvianum with contrasting results. Indeed, the expression of pat2 gene is affected by the introgression genetic background and by environmental conditions. Extreme daily and night temperatures and adverse radiation and relative humidity increase the parthenocarpy regulated by pat2 gene [74,75,76]. More recently, researchers demonstrated that Aucsia genes (mainly the DefH9-iaaM gene) regulate auxin synthesis and are responsible for parthenocarpic fruits. Two tomato lines (UC 82), transformed with DefH9-iaaM, were (almost) seedless and showed a higher β-carotene level compared to the control [77].
In Brassica genus, omics approaches have been adopted, not only to investigate pathogen resistance and abiotic stress tolerance (high temperature and high salinity) but also to phenotype and characterize biochemical pathways [33,78,79]. Metabolomic approaches have been used to target primary and secondary metabolites, mainly glucosinolates. These compounds are constituted by a sulphur-linked β-D glucopyranose and an amino-acid derived chain [78]. Park and co-workers correlated the amount of carotenoid, anthocyanin and phenolic acids to phenotypic variation and metabolic networks of several accessions for purple, white and green coloured cauliflowers [80]. Cauliflower breeding activities were focused on providing hybrids with high carotenoid content [81]. Interestingly, a crossbred between a cytoplasmic male sterile line (marketable white cauliflower) and an inbred line (orange mutant line with small curds) resulted in a marketable orange F1 with high nutritional quality. Indeed, the hybrid showed a 10.8 times higher β-carotene content than the parental white cauliflower [81].
RAPD, ISSR, and AFLP molecular markers have been applied to identify genetic diversity among several cultivars of cauliflower, cabbage, and broccoli [82]. Other useful applications for breeding purposes were the possibility to identify gene introgression among varieties and genetic relationships based on maturity group [82]. Another important milestone in cauliflower is the work of Zhao and co-authors [43]. A specific locus amplified fragments (SLAF) sequencing was used to identify large scale nucleotide polymorphism (SNP) and a high-density genetic map using a segregating population obtained by microspore culture. In this experiment, to design markers, the reference genome of the cabbage B. oleracea var. capitata (http://www.ocri-genomics.org/bolbase/ (accessed on 28 April 2021)) has been used. This research identified 81,311 SLAFs, 6815 of which showed polymorphisms between the parents of the segregating population. Interestingly, the distribution of the SLAFs markers was quite homogenous, throughout the reference genome, and therefore, the markers could be usefully applied for breeding purposes [37].
As reported by Zhao and co-authors (2016), genomic studies on B. oleracea used, in the past, parental material belonging to different subspecies to develop an intra-specific segregant population in order to identify QTLs, developing genetic maps and markers [43]. As a consequence, the application in terms of breeding was quite limited because the advances did not cover diverse genetic background referred to specific plant organs (e.g., curd in white cauliflower). Indeed, using intra-group parental lines, it has been possible to identify, for example, 13 QTLs associated to the curd-specific traits in white cauliflower [43,83].
As previously reported, the most important system for hybrid seed production is based on Ogura cytoplasmic male sterility, derived from radish and transferred to cauliflower in 1968 by back cross [84]. Dey and co-authors (2017) investigated the influence of Ogura cytoplasm on several qualitative traits related to antioxidant properties [85]. Since the radish chloroplasts of the original Ogura-based cytoplasm showed a negative interaction in the nuclear background of B. oleracea, the authors investigated if the substituted chloroplast (through somatic hybridization followed by backcross substitution) Ogura cybrid cytoplasm could still have adverse interaction with nuclear genes in cauliflower. The improved Ogura cybrid cytoplasm was obtained by protoplast fusion and regeneration in several Brassica species [86]. Interestingly, Dey and colleagues evidenced that Ogura cytoplasm is responsible of a reduction in anthocyanin, total chlorophylls, and ascorbic acid content in CMS lines compared to corresponding cauliflower inbred lines. On the other hand, CMS lines showed higher total carotenoid and β-carotene contents compared to inbred lines. This work evidenced that the effect of Ogura cybrid cytoplasm in different nuclear backgrounds is genotype-specific and the possibility to explore in detail the relevant metabolic pathways could fasten breeding to develop improved varieties for quality.
More recently, Sun and co-authors released the first high-quality genome sequence of B. oleracea [87] (Table 1). The cauliflower genome has been reported to be one of the biggest genomes in the Brassica genus, being 584.60 Mb with over 47,772 genes. Chromosome n. 3 emerged to be the most ancient in cauliflower, being inherited from the common ancestor of Brassicas. The authors performed a comparative genomic analysis and found thousands of cauliflower specific genes, which (presumably) differentially regulate curd formation and development. Indeed, the curd is the main phenotypic difference between cauliflower and similar species of the genus Brassica used in the comparative study [87].
6. Biotechnological Approaches: From Transgenesis to Genome Editing
The combined use of recombinant DNA technology, gene transfer methods, and tissue culture techniques substantially prompted the use of genetic engineering and transformation technologies in vegetables, both in public and private breeding programs, in accordance with local legislative restrictions. Transgenesis has the big advantage to transfer a single-gene without co-transfer undesirable genes unlike conventional breeding based on crossing two parental plants and subsequent back-cross cycles [95].
Among gene transfer methods, that based on Agrobacterium has been widely used on tomato and, to a lesser extent, on cauliflower. Tomato is one of the most used crops for genetic transformation because, due to its features, is often used as a model plant: 24 somatic chromosomes, a relatively small genome size (950 Mb per haploid nucleus), short generation time, easily reproduced by seed and by vegetative propagation, and relatively easy transformation protocols [96].
The first commercial release of recombinant DNA technology was the Flavr Savr tomato, also known as CGN-89564, approved by the Food and Drug Administration (Silver Spring, MD, USA) in 1994. That variety had a remarkable shelf-life due to the introduction of an antisense polygalacturonase gene, but it had also a bland taste, and it was dropped and removed from the market in 1996 [97,98]. In the second half of 1990, tomato Huafan No 1 was released in China; the variety 351N was constituted in the US by Agritope Inc. (Portland, OR, USA) while Monsanto (St. Louis, MO, USA) released 8338 and 5345 and DNA Plant Technology Corp. (Cinnaminson, NJ, USA) commercialized 1345-4 in USA, Japan, Mexico, Canada [98]. The above-mentioned varieties were transformed to increase shelf-life using different gene targets: antisense for polygalacturonase or antisense and gene suppression for enzymes belonging to the ethylene biosynthetic pathway [99]. Subsequently, transgenic tomatoes for improved starch biosynthesis, pectin, and sugar metabolism have been produced, as reported by Barg and co-authors [99].
In around 30 years of tomato genetic transformation, carotenoid and phenolic pathways have repeatedly been the subject of single and multi-gene engineering, as reviewed, for example, in Rosati and co-workers (2000) [100], Long and co-authors (2006) [101], and Giuliano (2014) [102]. Increase in β-carotene synthesis could be achieved by nuclear as well as chloroplast transformation [100,103]. The latter has remarkable advantages: no epigenetic effects leading to gene inactivation, high expression level, possibility of multiple transgene expression in synthetic operons and, being the plastids maternally inherited, reduced risk of transgene dispersal by outcrossing. Wurbs ad co-authors (2007) developed and applied a chloroplast transformation system that introduced in a panel of transgenic and mutant tomato linesa bacterial lycopene β-cyclase gene and led to a fourfold increase in pro-vitamin A content by converting the lycopene to β-carotene [103]. Subsequent investigations on plastid transformation, using both bacterial lycopene β-cyclase and a plant (Narcissus pseudonarcissus) lycopene β-cyclase, demonstrated a different action in tomato plants [104]. Indeed, the bacterial enzyme increased the amount of pro-vitamin A, but did not change carotenoid composition compared to WT. Furthermore, plant lycopene β-cyclase induced (similarly to the bacterial gene) a conversion of lycopene to β-carotene up to 1 mg/g dry weight of fruit and increased the total amount of carotenoids accumulated in the fruit, suggesting a role as regulatory enzyme [104].
Anthocyanin and flavonoid have also been investigated with transgenic techniques. Orzaez and co-authors (2009) reported that a transgenic tomato line (Del-Ros1) expressing Antirrhinum majus Delila (Del) and Rosea1 (Ros1) transcription factors showed a purple colour and an anthocyanin-rich phenotype [105]. Further investigations demonstrated that Del and Ros1 transformed tomatoes under the control of the fruit-specific E8 promoter produced higher anthocyanins contents than WT but equal to the content in blackberries and blueberries levels. The above-mentioned transgenes enhanced the hydrophilic antioxidant capacity of transformed tomato and resulted in purple colouration of both peel and flesh [106]. Furthermore, both Aft/Aft atv/atv and Ros1-Del transformed purple tomatoes showed, besides an improvement in their nutraceutical value, extended shelf life and lower susceptibility to the postharvest fungus Botrytis cinerea compared to normal tomato [107]. MicroTom plants were transformed using the promoter of the PLI gene because it is a light induced gene, and it is active mainly at skin level. The promoter was able to lead the expression of MYB transcriptional factor Rosea1 together with 35S Delila. The authors suggested that Aft/Aft atv/atv tomatoes and the transgenic tomato lines possess longer storage capability compared to WT due to delay in over-ripening. The higher content in anthocyanins at skin level increases the antioxidant activity and indirectly reduces pathogen spread [107].
Scientific attention has been addressed to stilbene family and mainly to resveratrol, a typical bioactive compound naturally presented in grape (for a review see Giovinazzo et al., 2013) [108]. Giovinazzo and co-authors (2005) produced transformed tomato for a stilbene synthase (StSy) transcriptionally regulated by the cauliflower mosaic virus (CaMV) 35 S promoter [109]. Trans-resveratrol and trans resveratrol-glucopyranoside were produced in tomato tissues and resulting in an increase in ascorbate and glutathione. As a consequence, the total antioxidant level increased compared to WT, but the basal level of tocopherole and lycopene did not change [109].
Recent genome editing tools provided novel functional insights in genes controlling, directly or indirectly, nutritional and organoleptic quality traits in various crops and hinted, at least in some cases, at the possibility to readily translate new knowledges and genotypes in applied breeding. Based on the fact that the tomato fruit colour depends on the accumulation of specific compounds directly related to organoleptic (flavour, aroma) and nutritional quality, Li Xindi et al. (2018) selected five genes (SGR1, LCY-E, LCY-B1, LCY-B2, and Blc) involved in lycopene biosynthesis and its conversion to carotene [110] (Table 2).
Using a multiplex approach for metabolic engineering, single, double, triple, and quadruple mutants were obtained, which showed, in comparison with wild type, a higher level of both lycopene and β-carotene. Mutants with homozygous and biallelic mutations could be readily used in breeding. The pink colour of fruits, a trait particularly appreciated in Asian countries, depends on the absence of yellow-coloured flavonoid naringenin chalcone (NarCh) in the fruit peel. Deng and colleagues (2018) reproduced the well-known y mutation, disrupting the SlMYB12 gene by CRISPR/Cas, changing the fruit colour of four elite inbred lines from red to pink [112]. The obtention of homozygous and biallelic mutations already in an elite background made the mutants readily usable. In a proof-of-concept study, Cermák et al. (2015) [113] showed the possibility to accumulate anthocyanin in different tissues of the tomato plant by substituting, through genome editing-driven homologous recombination, the promoter of the MYB transcription factor ANT1, being MYB (MYB–bHLH–WDR protein) complexes responsible for the transcriptional control of the flavonoid biosynthesis pathway [129,130,131].
Increasing the level of the non-proteinogenic amino acid GABA (γ-aminobutyric acid) in tomato fruits could help control blood pressure in hypertensive individuals. The actual level of GABA depends on the function of biosynthetic GAD genes, which have a C-terminal autoinhibitory domain, and on that of genes involved in catabolism (GABA-T and SSADH). In two independent studies, the GABA content in fruits was increased up to 19-fold that in controls by deleting, by CRISPR/Cas9 technology, the autoinhibitory domain of SlGAD2 and SlGAD3, regularly expressed during fruit development [108], or by a multiplex genome editing approach on five genes (GABA-TP1, GABA-TP2, GABA-TP3, CAT9, and SSADH), involved in GABA metabolism [115]. The “Sicilian Rouge” High GABA variety, edited using the former approach, will be the first genome editing product to be released in Japan (http://www.tomatonews.com/en/japan-breeders-launch-genome-edited-tomato_2_1236.html) (accessed on 28 April 2021).
A range of fruit metabolic phenotypes were obtained by Gago et al. (2017), who adopted the ZFN technology to disrupt the L1L4 gene, encoding one subunit of the heterotrimeric transcription factor Y [116]. In comparison with wild type, fruits of different edited lines showed an increase of some “positive” metabolites (e.g., SSC, fiber, fructose, ascorbic acid, total phenol, β-carotene) as well as a reduction in “negative” ones (e.g., oxalic acid). A “visionary” and futuristic objective was proposed by Rezende Naves and colleagues (2019), who suggested to activate pungency biosynthesis in tomato fruits in order to produce capsaicinoid-accumulating ‘Hot’ Tomatoes [132]. According to them, that objective could be achieved by the use of transcriptional activator-like effectors (TALEs) and the targeted replacement of promoters of relevant genes.
Organoleptic and nutritional traits in tomato fruits vary dramatically during ripening, which is a multidimensional process involving hundreds of genes with expression regulated by the hormone ethylene, ripening transcription factors, degree of DNA methylation, and several post-transcriptional processes (long non-coding RNAs, RNA-editing of mitochondrial transcripts). Genome-editing approaches in tomato, mostly based on CRISPR/Cas technology, allowed to generate new mutants and pinpoint the role of factors cited above [66,120,121,122,123,124]. The production of novel CRISPR/Cas mutants in genes coding for master regulators, such as RIN, NOR, and CNR, suggested new models of the ripening process, due to the different phenotypes obtained by genome editing compared to the original ones. Thanks to the novel mutants induced by CRISPR/Cas, a clearer picture is now available for rin, once thought to be a loss-of-function mutation, but now considered a gain-of-function one [117,118,119,133]. Those results might have an impact on breeding, since the use in commercial hybrids of the classical rin mutation in heterozygous state (Rin/rin) often resulted in poor flavour and reduced nutritional value [133]. CRISPR/Cas-based genome editing was also used to study the effect of genes involved in fruit softening during ripening. With the aim to slow down the softening process without altering colour, soluble solid content and other aspects of ripening, those studies highlighted the role of PL, encoding a pectate lyase, and the possibility to use such a mutant in tomato breeding [121,125,133]. A result with great potential for tomato breeding was obtained by Yu and colleagues (2017), who, using CRISPR/Cas, introduced viaHDR the mutation alcobaca (alc), responsible for long shelf-life, in a normal background. Fruit colour and other agronomic traits were unaffected [127].
Finally, in a recent paper, S. pimpinellifolium, the wild closest ancestor of cultivated tomato, was de novo domesticated by editing simultaneously few major genes. Among them, editing of Lycopene Beta Cyclase (CycB) resulted in 500% improvement of fruit lycopene content compared to S. lycopersicum [128].
In cauliflower, Nugent and co-authors (2006) reported nuclear and plastid transformation using PEG-mediated uptake of DNA into protoplasts. Cauliflower has been also transformed using A. tumefaciens with isopentenyl transferase (ipt) gene under the control of a senescence-associated gene promoter, pSAG12, isolated from A. thaliana [134]. The pSAG12:ipt gene was evidenced to be responsible for delay in leaf senescence, altered synchronous curd initiation, smaller curd size, and higher susceptibility to fungal infection [135]. More recently, Kowalczyk et al. (2018) developed a high efficiency cauliflower transformation method using Rhizobium (=Agrobacterium) rhizogenes in hypocotyl sections of 10-day-old de-rooted seedling [136]. Zhou and co-workers (2008) isolated the Or gene from orange cauliflower and introduced the transgene in potato revealing that the gene controls carotenoid accumulation by inducing the formation of chromoplasts [137].
7. Conclusions and Perspectives
Domestication and plant breeding increased yield and productivity but led to genetic erosion and reduction in nutritional value and taste in tomato crop [64,128]. Based on the previous assumption, the current commercial varieties should show a detrimental in quality compared to old landraces. Nevertheless, Schouten and co-authors evidenced an increase in diversity for aroma compounds in the cultivars grown in The Netherlands using a combined approach of genomics and metabolomics [142]. Interestingly, this consideration demonstrates the need to develop further investigations on the complex genetic mechanism controlling quality in vegetables.
In this review, we highlighted the importance of quality traits in tomato and cauliflower, focusing on phytochemical and bio-active compounds linked to flavour and taste and how plant breeding can respond to novel consumers’ requirements. The two crops can be considered as model systems for fleshy fruit and inflorescence edible portion, respectively. Comparing plant breeding activities and results achieved in the two species, a relevant gap could be denoted, depending not only on the higher economic value of tomato but also on a reduced availability of genomic and biotechnological resources in cauliflower [5,136].
Advances in -omics technologies make it possible to elucidate the genetic and molecular bases of the multiple traits composing quality using genetic, genomic, and metabolomics approaches, particularly in case of pleiotropy and unfavourable trait association [143]. The application of CRISPR/Cas in tomato breeding triggered the so-called de novo domestication of tomato varieties by introducing elite traits evolved during domestication and selective breeding into its wild relative/ancestor [144]. A similar technology, applied to cauliflower, could fasten breeding, reducing the gap with other model system crops. Furthermore, CRISPR/Cas could be used also in mutagenesis, being more precise than random mutagenesis technologies using chemicals or radiations. These tools are continuously improving to become more efficient and precise for an era of fast breeding for improved nutritional and organoleptic quality in vegetables.
Plant breeding programs are more and more increasing into complexity in terms of traits and underlying genetic and physiological factors. Nevertheless, the new genomic tools and genome editing approaches will help face such complexity increasing the efficiency of selection and the precision of new variability induction. As a result, positive implications related to the bioavailability and exploitation of functional compounds in improved vegetables can be expected [145].
Consumer acceptance of gene-edited vegetables is also an important issue to be considered to reach a commercial success, depending on several variables. Indeed, Kato-Nitta and co-authors (2019) reported the results of a survey conducted in Japan on expert and public perceptions on gene-edited crops. The study revealed the uncertainty for public acceptance but also the need of knowledge toward emerging breeding technologies in order to understand the value and risk perceptions [146].
The excessive regulatory load and the scale of investment required for approval of new genotypes has limited the application of transgenic technology to a few crops for extensive farming (thus not including vegetables) and a few characters (almost exclusively resistance to insects and herbicides). The novelty and value of genomic editing in applied breeding has largely recognized. Nevertheless, before such technology could be fully implemented and largely produce practical results, it will be important that a science-based regulatory frame be applied consistently worldwide [147,148,149,150,151].
Author Contributions
All the authors contributed equally. All authors have read and agreed to the published version of the manuscript.
Funding
Tomato breeding activities at CREA Research Centre for Vegetable and Ornamental Crops are presently carried out in the framework of BRESOV and LIVESEED projects, funded by the European Union Horizon 2020 research and innovation program under grant agreements Nos. 774244 and 727230, respectively.
Data Availability Statement
Not applicable.
Acknowledgments
The authors acknowledge the BRESOV project for supporting the publication costs of this review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pott, D.M.; Durán-Soria, S.; Osorio, S.; Vallarino, J.G. Combining metabolomic and transcriptomic approaches to assess and improve crop quality traits. CABI Agric. Biosci. 2021, 2, 1–20. [Google Scholar] [CrossRef]
- Crossa, J.; Fritsche-Neto, R.; Montesinos-Lopez, O.A.; Costa-Neto, G.; Dreisigacker, S.; Montesinos-Lopez, A.; Bentley, A.R. The Modern Plant Breeding Triangle: Optimizing the Use of Genomics, Phenomics, and Enviromics Data. Front. Plant Sci. 2021, 12, 651480. [Google Scholar] [CrossRef]
- Aharoni, A.; O’Connell, A.P. Gene expression analysis of strawberry achene and receptacle maturation using DNA microarrays. J. Exp. Bot. 2002, 53, 2073–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, S.S.; Singh, N.; Bhatia, R.; Parkash, C.; Chandel, C. Genetic combining ability and heterosis for important vitamins and antioxidant pigments in cauliflower (Brassica oleracea var. botrytis L.). Euphytica 2014, 195, 169–181. [Google Scholar] [CrossRef]
- Cardi, T.; D’Agostino, N.; Tripodi, P. Genetic Transformation and Genomic Resources for Next-Generation Precise Genome Engineering in Vegetable Crops. Front. Plant Sci. 2017, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Jorasch, P. The global need for plant breeding innovation. Transgenic Res. 2019, 28, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Churchill, F.B. William Johannsen and the genotype concept. J. Hist. Biol. 1974, 7, 5–30. [Google Scholar] [CrossRef]
- D’Amato, F. Introduction to Plant Genetic. In Italian Contribution to Plant Genetic and Breeding; Scarascia Mugnozza, G.T., Pagnotta, M.A., Eds.; Tipolitografia Quatrini A&F: Viterbo, Italy, 1998; pp. 21–53. [Google Scholar]
- Strampelli, N. Alla Ricerca e Creazione di Nuove Varietà di Frumenti a Mezzo di Ibridazione; Ti-Pografia Unione Coop. Editrice: Rome, Italy, 1907. [Google Scholar]
- Borojevic, K. Historic role of the wheat variety Akakomugi in Southern and Central European Wheat Breeding Programs. Breed. Sci. 2005, 55, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Pray, L.; Zhaurova, K. Barbara McClintock and the discovery of jumping genes (transposons). Nat. Sci. Educ. 2008, 1, 169. [Google Scholar]
- Somssich, M. A short history of plant transformation. PeerJ 2019. [Google Scholar] [CrossRef] [Green Version]
- Swaminathan, M.S. Preface. In Marker-Assisted Plant Breeding, Principles and Practices; Singh, B.D., Singh, A.K., Eds.; Springer: New Delhi, India, 2015; pp. vii–viii. [Google Scholar]
- Grube, R.C.; Radwanski, E.R.; Jahn, M. Comparative Genetics of Disease Resistance within the Solanaceae. Genetics 2000, 155, 873–887. [Google Scholar] [CrossRef]
- Cardi, T.; Stewart, C.N. Progress of targeted genome modification approaches in higher plants. Plant. Cell Rep. 2016, 35, 1401–1416. [Google Scholar] [CrossRef] [PubMed]
- Bortesi, L.; Fischer, R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Klee, H.J.; Tieman, D.M. The genetics of fruit flavour preferences. Nat. Rev. Genet. 2018, 19, 347–356. [Google Scholar] [CrossRef]
- Chan, E.K.F.; Rowe, H.C.; Hansen, B.G.; Kliebenstein, D.J. The Complex Genetic Architecture of the Metabolome. PLoS Genet. 2010, 6, e1001198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauvage, C.; Segura, V.; Bauchet, G.; Stevens, R.; Do, P.T.; Nikoloski, Z.; Fernie, A.R.; Causse, M. Genome-Wide Association in Tomato Reveals 44 Candidate Loci for Fruit Metabolic Traits. Plant Physiol. 2014, 165, 1120–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Wang, W.; Peng, M.; Gong, L.; Gao, Y.; Wan, J.; Wang, S.; Shi, L.; Zhou, B.; Li, Z.; et al. Comparative and parallel genome-wide association studies for metabolic and agronomic traits in cereals. Nat. Commun. 2016, 7, 12767. [Google Scholar] [CrossRef]
- Iqbal, R.K.; Saeed, K.; Khan, A.; Noreen, I.; Bashir, M. Tomato (Lycopersicon esculetum) fruit improvement through breeding. Sch. J. Appl. Sci. Res. 2019, 2, 21–25. [Google Scholar]
- Campanelli, G.; Sestili, S.; Acciarri, N.; Montemurro, F.; Palma, D.; Leteo, F.; Beretta, M. Multi-Parental Advances Generation Inter-Cross Population, to Develop Organic Tomato Genotypes by Participatory Plant Breeding. Agronomy 2019, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Gramazio, P.; Pereira-Dias, L.; Vilanova, S.; Prohens, J.; Soler, S.; Esteras, J.; Garmendia, A.; Díez, M.J. Morphoagronomic characterization and whole-genome resequencing of eight highly diverse wild and weedy S. pimpinellifolium and S. lycopersicum var. cerasiforme accessions used for the first interspecific tomato MAGIC population. Hortic. Res. 2020, 7, 1–16. [Google Scholar] [CrossRef]
- Esposito, S.; Cardi, T.; Campanelli, G.; Sestili, S.; Díez, M.J.; Soler, S.; Prohens, J.; Tripodi, P. ddRAD sequencing-based genotyping for population structure analysis in cultivated tomato provides new insights into the genomic diversity of Mediterranean ‘da serbo’ type long shelf-life germplasm. Hortic. Res. 2020, 7, 1–14. [Google Scholar] [CrossRef]
- Martínez-Cuenca, M.-R.; Pereira-Dias, L.; Soler, S.; López-Serrano, L.; Alonso, D.; Calatayud, Á.; Díez, M.J. Adaptation to Water and Salt Stresses of Solanum pimpinellifolium and Solanum lycopersicum var. cerasiforme. Agronomy 2020, 10, 1169. [Google Scholar] [CrossRef]
- Rosa-Martínez, E.; Adalid, A.M.; Alvarado, L.E.; Burguet, R.; García-Martínez, M.D.; Pereira-Dias, L.; Casanova, C.; Soler, E.; Figàs, M.R.; Plazas, M.; et al. Variation for Composition and Quality in a Collection of the Resilient Mediterranean ‘de penjar’ Long Shelf-Life Tomato Under High and Low N Fertilization Levels. Front. Plant Sci. 2021, 12, 633957. [Google Scholar] [CrossRef]
- Gao, Y.; Zhu, N.; Zhu, X.; Wu, M.; Jiang, C.-Z.; Grierson, D.; Luo, Y.; Shen, W.; Zhong, S.; Fu, D.-Q.; et al. Diversity and redundancy of the ripening regulatory networks revealed by the fruit ENCODE and the new CRISPR/Cas9 CNR and NOR mutants. Hortic. Res. 2019, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- The Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Cheema, D.S.; Dhaliwal, M.S. Hybrid Tomato Breeding. J. New Seeds 2005, 6, 1–14. [Google Scholar] [CrossRef]
- Schiavi, M.; Vitelli, G.; Maestrelli, A.; Forni, E.; Giovannessi, L. Breeding of green curded cauliflower (Brassica oleracea L., botrytis group) for fresh market and freezing. Acta Hortic. 1998, 459, 403–410. [Google Scholar] [CrossRef]
- Acciarri, N.; Branca, F.; Sabatini, E.; Argento, S.; Magnifico, V. Miglioramento genetico dei cavolfiori a corimbo bianco e colourato. L’Inf. Agrar. 2004, 25, 33–36. [Google Scholar]
- Schulz, O.E. Cruciferae-Brassicaceae part I, Brassicinae and raphaninae. In Das Pflanzenreich; Wilhelm Engelmann: Leipzig, Germany, 1919; pp. 1–290. [Google Scholar]
- Smith, L.B.; King, G.J. The distribution of BoCAL-a alleles in Brassica oleracea is consistent with a genetic model for curd development and domestication of the cauliflower. Mol. Breed. 2000, 6, 603–613. [Google Scholar] [CrossRef]
- Singh, B.K.; Singh, B.; Singh, P.M. Breeding Cauliflower: A Review. Int. J. Veg. Sci. 2018, 24, 58–84. [Google Scholar] [CrossRef]
- Pelletier, G.; Primard, C.; Vedel, F.; Chétrit, P.; Rémy, R.; Roussell, P.; Renard, M. Intergeneric cytoplasmic hybridization in Cruciferae by protoplast fusion. Mol. Gen. Genet. 1983, 191, 244–250. [Google Scholar] [CrossRef]
- Kucera, V.; Chytilov, V.; Vyvadilov, M.; Klima, M. Hybrid breeding of cauliflower using self-incompatibility and cytoplasmic male sterility. Hortic. Sci. 2006, 33, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Kyriacou, M.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Dorais, A.; Gosselin, A.; Papadopolus, M. Greenhouse Tomato Fruit Quality. Hortic. Rev. 2001, 26, 238–262. [Google Scholar]
- Barry, C.; Giovannoni, J.J. Ethylene and Fruit Ripening. J. Plant. Growth Regul. 2007, 26, 143–159. [Google Scholar] [CrossRef]
- Conesa, M.À.; Fullana-Pericàs, M.; Granell, A.; Galmés, J. Mediterranean Long Shelf-Life Landraces: An Untapped Genetic Resource for Tomato Improvement. Front. Plant Sci. 2020, 10, 1651. [Google Scholar] [CrossRef]
- Wang, D.; Seymour, G.B. Tomato Flavor: Lost and Found? Mol. Plant 2017, 10, 782–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambla, J.L.; Tikunov, Y.M.; Monforte, A.J.; Bovy, A.G.; Granell, A. The expanded tomato fruit volatile landscape. J. Exp. Bot. 2013, 65, 4613–4623. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Gu, H.; Sheng, X.; Yu, H.; Wang, J.; Huang, L.; Wang, D. Genome-Wide Single-Nucleotide Polymorphisms Discovery and High-Density Genetic Map Construction in Cauliflower Using Specific-Locus Amplified Fragment Sequencing. Front. Plant Sci. 2016, 7, 334. [Google Scholar] [CrossRef] [Green Version]
- Raja, M.M.; Raja, M.; Imran, M.; Rahmsn, H.A. Quality aspects of cauliflower during storage. Int. Food Res. J. 2011, 18, 427–431. [Google Scholar]
- Fernie, A.R.; Tadmor, Y.; Zamir, D. Natural genetic variation for improving crop quality. Curr. Opin. Plant Biol. 2006, 9, 196–202. [Google Scholar] [CrossRef]
- Varalakshmi, B. Heterosis and combining ability for yield and its components in early cauliflower. Indian J. Hortic. 2009, 66, 198–203. [Google Scholar]
- Singh, S.; Kalia, P.; Meena, R.K.; Mangal, M.; Islam, S.; Saha, S.; Tomar, B.S. Genetics and Expression Analysis of Anthocyanin Accumulation in Curd Portion of Sicilian Purple to Facilitate Biofortification of Indian Cauliflower. Front. Plant Sci. 2020, 10, 1766. [Google Scholar] [CrossRef] [Green Version]
- Chiu, L.-W.; Zhou, X.; Burke, S.; Wu, X.; Prior, R.L.; Li, L. The Purple Cauliflower Arises from Activation of a MYB Transcription Factor. Plant Physiol. 2010, 154, 1470–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, L.W. Transposon Insertion at the Promoter of a MYB Transcription Factor Results in Ectopic Anthocyanins Accumulation in Purple Cauliflower (Brassica oleracea var. botrytis). Ph.D. Thesis, Cornell University, Ithaca, NY, USA, 2010. [Google Scholar]
- Johnson, E.J.; Hammond, B.; Yeum, K.-J.; Qin, J.; Wang, X.D.; Castaneda, C.; Snodderly, D.; Russell, R.M. Relation among serum and tissue concentrations of lutein and zeaxanthin and macular pigment density. Am. J. Clin. Nutr. 2000, 71, 1555–1562. [Google Scholar] [CrossRef] [Green Version]
- Crisp, P.; Walkey, D.G.A.; Bellman, E.; Roberts, E. A mutation affecting curd colour in cauliflower (Brassica oleracea L. var. Botrytis DC). Euphytica 1975, 24, 173–176. [Google Scholar] [CrossRef]
- Dickson, M.H.; Lee, C.Y.; Blamble, A.E. Orange-curd high carotene cauliflower inbreds, NY 156, NY 163, and NY Hort165. Science 1998, 23, 778–779. [Google Scholar]
- Li, L.; Paolillo, D.J.; Parthasarathy, M.V.; DiMuzio, E.M.; Garvin, D.F. A novel gene mutation that confers abnormal patterns of β-carotene accumulation in cauliflower (Brassica oleracea var. botrytis). Plant J. 2001, 26, 59–67. [Google Scholar] [CrossRef]
- Wang, L.-S.; Stoner, G.D. Anthocyanins and their role in cancer prevention. Cancer Lett. 2008, 269, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Volden, J.; Bengtsson, G.B.; Wicklund, T. Glucosinolates, l-ascorbic acid, total phenols, anthocyanins, antioxidant capacities and colour in cauliflower (Brassica oleracea L. ssp. botrytis); effects of long-term freezer storage. Food Chem. 2009, 112, 967–976. [Google Scholar] [CrossRef]
- Scalzo, R.L.; Genna, A.; Branca, F.; Chedin, M.; Chassaigne, H. Anthocyanin composition of cauliflower (Brassica oleracea L. var. botrytis) and cabbage (B. oleracea L. var. capitata) and its stability in relation to thermal treatments. Food Chem. 2008, 107, 136–144. [Google Scholar] [CrossRef]
- Tommonaro, G.; De Prisco, R.; Abbamondi, G.R.; Marzocco, S.; Saturnino, C.; Poli, A.; Nicolaus, B. Evaluation of Antioxidant Properties, Total Phenolic Content, and Biological Activities of New Tomato Hybrids of Industrial Interest. J. Med. Food 2012, 15, 483–489. [Google Scholar] [CrossRef]
- Blando, F.; Berland, H.; Maiorano, G.; Durante, M.; Mazzucato, A.; Picarella, M.E.; Nicoletti, I.; Gerardi, C.; Mita, G.; Andersen, Ø.M. Nutraceutical Characterization of Anthocyanin-Rich Fruits Produced by “Sun Black” Tomato Line. Front. Nutr. 2019, 6, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzucato, A.; Willems, D.; Bernini, R.; Picarella, M.E.; Santangelo, E.; Ruiu, F.; Tilesi, F.; Soressi, G.P. Novel phenotypes related to the breeding of purple-fruited tomatoes and effect of peel extracts on human cancer cell proliferation. Plant Physiol. Biochem. 2013, 72, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Dono, G.; Rambla, J.L.; Frusciante, S.; Granell, A.; Diretto, G.; Mazzucato, A. Color Mutations Alter the Biochemical Composition in the San Marzano Tomato Fruit. Metabolites 2020, 10, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gapper, N.; Giovannoni, J.J.; Watkins, C. Understanding development and ripening of fruit crops in an ‘omics’ era. Hortic. Res. 2014, 1, 14034. [Google Scholar] [CrossRef] [Green Version]
- Carli, P.; Barone, A.; Fogliano, V.; Frusciante, L.; Ercolano, M.R. Dissection of genetic and environmental factors involved in tomato organoleptic quality. BMC Plant Biol. 2011, 11, 58. [Google Scholar] [CrossRef] [Green Version]
- Ercolano, M.R.; Sacco, A.; Ferriello, F.; D’Alessandro, R.; Tononi, P.; Traini, A.; Barone, A.; Zago, E.; Chiusano, M.L.; Buson, G.; et al. Patchwork sequencing of tomato San Marzano and Vesuviano varieties highlights genome-wide variations. BMC Genom. 2014, 15, 138. [Google Scholar] [CrossRef] [Green Version]
- Tieman, D.; Zhu, G., Jr.; Lin, T.; Nguyen, C.; Bies, D.; Rambla, J.L.; Beltran, K.S.O.; Taylor, M.; Zhang, B.; Ikeda, H. A chemical genetic roadmap to improved tomato flavor. Science 2017, 355, 391–394. [Google Scholar] [CrossRef]
- Tranchida-Lombardo, V.; Cigliano, R.A.; Anzar, I.; Landi, S.; Palombieri, S.; Colantuono, C.; Bostan, H.; Termolino, P.; Aversano, R.; Batelli, G.; et al. Wholegenome resequencing of two Italian tomato landraces reveals sequence variations in genes associated with stress tolerance, fruit quality and long shelf-life traits. DNA Res. 2018, 25, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Gonda, I.; Sun, H.; Ma, Q.; Bao, K.; Tieman, D.M.; Chang, E.; Fish, T.L.; Stromberg, K.A.; Sacks, G.L.; et al. The tomato pan-genome uncovers new genes and a rare allele regulating fruit flavor. Nat. Genet. 2019, 51, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Tanksley, S.D. Chromosomal evolution in the plant family Solanaceae. BMC Genom. 2010, 11, 182. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, C.J.; Carrari, F.; Bauke, A.; Overy, S.; Hill, S.A.; Quick, P.W.; Fernie, A.R.; Sweetlove, L.J. Fruit carbohydrate me-tabolism in an introgression line of tomato with increased fruit soluble solids. Plant Cell Physiol. 2005, 46, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Colanero, S.; Perata, P.; Gonzali, S. The atroviolacea Gene Encodes an R3-MYB Protein Repressing Anthocyanin Synthesis in Tomato Plants. Front. Plant Sci. 2018, 9, 830. [Google Scholar] [CrossRef] [Green Version]
- Tanksley, S.D. The Genetic, Developmental, and Molecular Bases of Fruit Size and Shape Variation in Tomato. Plant Cell 2004, 16, S181–S189. [Google Scholar] [CrossRef]
- Adhikari, P.; McNellie, J.; Panthee, D.R. Detection of Quantitative Trait Loci (QTL) Associated with the Fruit Morphology of Tomato. Genes 2020, 11, 1117. [Google Scholar] [CrossRef]
- Casas-Diaz, A.V.; Hewitt, J.D.; Lapushner, D. Effects of parthenocarpy on fruit quality in tomato. J. Am. Soc. Hortic. Sci. 1987, 112, 634–637. [Google Scholar]
- Mazzucato, A.; Taddei, A.; Soressi, G. The parthenocarpic fruit (pat) mutant of tomato (Lycopersicon esculentum Mill.) sets seedless fruits and has aberrant anther and ovule development. Development 1998, 125, 107–114. [Google Scholar] [CrossRef]
- Acciarri, N.; Ferrari, V.; Vitelli, G.; Ficcadenti, N.; Pandolfini, T.; Spena, A.; Rotino, G.L. Effetto della partenocarpia in ibridi di pomodoro geneticamente modificati. L’Inf. Agrar. 2000, 4, 117–122. [Google Scholar]
- Beraldi, D.; Picarella, M.E.; Soressi, G.P.; Mazzucato, A. Fine mapping of the parthenocarpic fruit (pat) mutation in tomato. Theor. Appl. Genet. 2003, 108, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Rotino, G.L.; Acciarri, N.; Sabatini, E.; Mennella, G.; Scalzo, R.L.; Maestrelli, A.; Molesini, B.; Pandolfini, T.; Scalzo, J.; Mezzetti, B.; et al. Open field trial of genetically modified parthenocarpic tomato: Seedlessness and fruit quality. BMC Biotechnol. 2005, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Ewitzel, K.; Eneugart, S.; Eruppel, S.; Schreiner, M.; Ewiesner, M.; Ebaldermann, S. Recent progress in the use of ‘omics technologies in brassicaceous vegetables. Front. Plant Sci. 2015, 6, 244. [Google Scholar] [CrossRef] [Green Version]
- Francisco, M.; Soengas, P.; Velasco, P.; Bhadauria, V.; Cartea, M.; Rodríguez, V.M. Omics Approach to Identify Factors Involved in Brassica Disease Resistance. Curr. Issues Mol. Biol. 2015, 19, 31–42. [Google Scholar]
- Park, S.-Y.; Lim, S.-H.; Ha, S.-H.; Yeo, Y.; Park, W.T.; Kwon, D.Y.; Park, S.U.; Kim, J.K. Metabolite Profiling Approach Reveals the Interface of Primary and Secondary Metabolism in Colored Cauliflowers (Brassica oleracea L. ssp. botrytis). J. Agric. Food Chem. 2013, 61, 6999–7007. [Google Scholar] [CrossRef]
- Ding, Y.; Jian, Y. An Orange Cauliflower Hybrid ‘Jinyu 60’ with High Β-Carotene. Acta Hortic. 2010, 856, 261–264. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Xu, J.; Xu, Y.; Liu, L. Cultivar identification and genetic diversity analysis of cauliflower with molecular markers. Acta Hortic. 2011, 918, 315–321. [Google Scholar] [CrossRef]
- Lv, H.; Wang, Q.; Zhang, Y.; Yang, L.; Fang, Z.; Wang, X.; Liu, Y.; Zhuang, M.; Lin, Y.; Yu, H.; et al. Linkage map construction using InDel and SSR markers and QTL analysis of heading traits in Brassica oleracea var. capitata L. Mol. Breed. 2014, 34, 87–98. [Google Scholar] [CrossRef]
- Ogura, H. Studies on the new male sterility in Japanese radish with special reference to the utilisation of this sterility towards the pratical raising of hybrid seeds. Mem. Fac. Agric. Kagoshima Univ. 1968, 6, 39–78. [Google Scholar]
- Dey, S.S.; Bhatia, R.; Parkash, C.; Sharma, S.; Dabral, M.; Mishra, V.; Bhardwaj, I.; Sharma, K.; Sharma, V.K.; Kumar, R. Alteration in important quality traits and antioxidant activities in Brassica oleracea with Oguracybrid cytoplasm. Plant Breed. 2017, 136, 400–409. [Google Scholar] [CrossRef]
- Acciarri, N.; Cardi, T. Brassica. In Italian Contribution to Plant Genetic and Breeding; Scarascia Mugnozza, G.T., Pagnotta, M.A., Eds.; Tipolitografia Quatrini A&F: Viterbo, Italy, 1998; pp. 481–486. [Google Scholar]
- Sun, B.; Zheng, A.; Jiang, M.; Xue, S.; Yuan, Q.; Jiang, L.; Chen, Q.; Li, M.; Wang, Y.; Zhang, Y.; et al. CRISPR/Cas9-mediated mutagenesis of homologous genes in Chinese kale. Sci. Rep. 2018, 8, 16786. [Google Scholar] [CrossRef]
- Bolger, A.; Scossa, F.; Bolger, M.E.; Lanz, C.; Maumus, F.; Tohge, T.; Quesneville, H.; Alseekh, S.; Sørensen, I.; Lichtenstein, G.; et al. The genome of the stress-tolerant wild tomato species Solanum pennellii. Nat. Genet. 2014, 46, 1034–1038. [Google Scholar] [CrossRef]
- Wang, X.; Gao, L.; Jiao, C.; Stravoravdis, S.; Hosmani, P.S.; Saha, S.; Zhang, J.; Mainiero, S.; Strickler, S.R.; Catala, C.; et al. Genome of Solanum pimpinellifolium provides insights into structural variants during tomato breeding. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Razali, R.; Bougouffa, S.; Morton, M.J.L.; Lightfoot, D.J.; Alam, I.; Essack, M.; Arold, S.T.; Kamau, A.A.; Schmöckel, S.M.; Pailles, Y.; et al. The Genome Sequence of the Wild Tomato Solanum pimpinellifolium Provides Insights into Salinity Tolerance. Front. Plant Sci. 2018, 9, 1402. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Wang, C.; Zhang, X.; Zhang, W.; Jiang, H.; Yao, X.; Liu, L.; Wen, Z.; Niu, G.; Shan, X. Draft genome sequence of cauliflower (Brassica oleracea L. var. botrytis) provides new insights into the C genome in Brassica species. Hortic. Res. 2019, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.-H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liu, D.; Wang, X.; Ji, C.; Cheng, F.; Liu, B.; Hu, Z.; Chen, S.; Pental, D.; Ju, Y.; et al. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat. Genet. 2016, 48, 1225–1232. [Google Scholar] [CrossRef]
- Bayer, P.E.; Hurgobin, B.; Golicz, A.A.; Chan, C.K.; Yuan, Y.; Lee, H.; Renton, M.; Meng, J.; Li, R.; Long, Y.; et al. Assembly and comparison of two closely related Brassica napus genomes. Plant Biotechnol. J. 2017, 15, 1602–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosal, S.S.; Wani, S. Plant Genetic Transformation and Transgenic Crops: Methods and Applications. In Biotechnologies of Crop Improvement; Springer: Cham, Switzerland, 2018; Volume 2, pp. 1–23. [Google Scholar]
- Di Matteo, A.; Manuela, M.; Sacco, A.; Frusciante, L.; Barone, A. Genetic Transformation in Tomato: Novel Tools to Improve Fruit Quality and Pharmaceutical Production. In Genetic Transformation; InTech: Wheeling, IL, USA, 2011. [Google Scholar]
- Krieger, E.K.; Allen, E.; Gilbertson, L.A.; Roberts, J.K.; Hiatt, W.; Sanders, R.A. The Flavr Savr Tomato, an Early Example of RNAi Technology. HortScience 2008, 43, 962–964. [Google Scholar] [CrossRef]
- Gerszberg, A.; Hnatuszko-Konka, K.; Kowalczyk, T.; Kononowicz, A.K. Tomato (Solanum lycopersicum L.) in the service of biotechnology. Plant. Cell Tissue Organ. Cult. 2015, 120, 881–902. [Google Scholar] [CrossRef] [Green Version]
- Barg, R.; Shabtai, S.; Salts, Y. Transgenic Tomato (Lycopersicom esculentum). In Biotechnology in Agriculture and Forestry 47, Transgenic Crop; Bajaj, A., Ed.; Springer: New York, NY, USA, 2012; Volume 2, pp. 219–233. [Google Scholar]
- Rosati, C.; Aquilani, R.; Dharmapuri, S.; Pallara, P.; Marusic, C.; Tavazza, R.; Bouvier, F.; Camara, B.; Giuliano, G. Metabolic engineering of beta-carotene and lycopene content in tomato fruit. Plant J. 2000, 24, 413–420. [Google Scholar] [CrossRef]
- Long, M.; Millar, D.J.; Kimura, Y.; Donovan, G.; Rees, J.; Fraser, P.D.; Bramley, P.M.; Bolwell, G.P. Metabolite profiling of carotenoid and phenolic pathways in mutant and transgenic lines of tomato: Identification of a high antioxidant fruit line. Phytochemistry 2006, 67, 1750–1757. [Google Scholar] [CrossRef]
- Giuliano, G. Plant carotenoids: Genomics meets multi-gene engineering. Curr. Opin. Plant Biol. 2014, 19, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Wurbs, D.; Ruf, S.; Bock, R. Contained metabolic engineering in tomatoes by expression of carotenoid biosynthesis genes from the plastid genome. Plant J. 2006, 49, 276–288. [Google Scholar] [CrossRef]
- Apel, W.; Bock, R. Enhancement of Carotenoid Biosynthesis in Transplastomic Tomatoes by Induced Lycopene-to-Provitamin A Conversion. Plant Physiol. 2009, 151, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orzaez, D.; Medina, A.; Torre, S.; Fernández-Moreno, J.P.; Rambla, J.L.; Fernández-Del-Carmen, A.; Butelli, E.; Martin, C.; Granell, A. A Visual Reporter System for Virus-Induced Gene Silencing in Tomato Fruit Based on Anthocyanin Accumulation. Plant Physiol. 2009, 150, 1122–1134. [Google Scholar] [CrossRef] [Green Version]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.-P.; Matros, A.; Peterek, S.; Schijlen, E.G.W.M.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- Bassolino, L.; Zhang, Y.; Schoonbeek, H.-J.; Kiferle, C.; Perata, P.; Martin, C. Accumulation of anthocyanins in tomato skin extends shelf life. New Phytol. 2013, 200, 650–655. [Google Scholar] [CrossRef] [Green Version]
- Giovinazzo, G.; Ingrosso, I.; Taurino, M.; Santino, A. Metabolic Engineering for Functional Foods: Tomato Fruits and Stilbenes. In Natural Products; Springer: Cham, Switzerland, 2013; pp. 1581–1597. [Google Scholar]
- Giovinazzo, G.; D’Amico, L.; Paradiso, A.; Bollini, R.; Sparvoli, F.; De Gara, L. Antioxidant metabolite profiles in tomato fruit constitutively expressing the grapevine stilbene synthase gene. Plant Biotechnol. J. 2004, 3, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Y.; Chen, S.; Tian, H.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. Lycopene Is Enriched in Tomato Fruit by CRISPR/Cas9-Mediated Multiplex Genome Editing. Front. Plant Sci. 2018, 9, 559. [Google Scholar] [CrossRef]
- D’Ambrosio, C.; Stigliani, A.L.; Giorio, G. CRISPR/Cas9 editing of carotenoid genes in tomato. Transgenic Res. 2018, 27, 367–378. [Google Scholar] [CrossRef]
- Deng, L.; Wang, H.; Sun, C.; Li, Q.; Jiang, H.; Du, M.; Li, C.-B.; Li, C. Efficient generation of pink-fruited tomatoes using CRISPR/Cas9 system. J. Genet. Genom. 2018, 45, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Čermák, T.; Baltes, N.J.; Čegan, R.; Zhang, Y.; Voytas, D.F. High-frequency, precise modification of the tomato genome. Genome Biol. 2015, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, S.; Arai, C.; Takayama, M.; Matsukura, C.; Ezura, H. Efficient increase of ɣ-aminobutyric acid (GABA) content in tomato fruits by targeted mutagenesis. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Li, R.; Li, X.; Fu, D.; Zhu, B.; Tian, H.; Luo, Y.; Zhu, H. Multiplexed CRISPR/Cas9-mediated metabolic engineering of γ-aminobutyric acid levels in Solanum lycopersicum. Plant. Biotechnol. J. 2018, 16, 415–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gago, C.; Drosou, V.; Paschalidis, K.; Guerreiro, A.; Miguel, G.; Antunes, D.; Hilioti, Z. Targeted gene disruption coupled with metabolic screen approach to uncover the LEAFY COTYLEDON1-LIKE4 (L1L4) function in tomato fruit metabolism. Plant Cell Rep. 2017, 36, 1065–1082. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Nishizawa-Yokoi, A.; Endo, M.; Mikami, M.; Toki, S. CRISPR/Cas9-mediated mutagenesis of the RIN locus that regulates tomato fruit ripening. Biochem. Biophys. Res. Commun. 2015, 467, 76–82. [Google Scholar] [CrossRef]
- Ito, Y.; Nishizawa-Yokoi, A.; Endo, M.; Mikami, M.; Shima, Y.; Nakamura, N.; Kotake-Nara, E.; Kawasaki, S.; Toki, S. Re-evaluation of the rin mutation and the role of RIN in the induction of tomato ripening. Nat. Plants 2017, 3, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhu, B.; Pirrello, J.; Xu, C.; Zhang, B.; Bouzayen, M.; Chen, K.; Grierson, D. Roles of RIN and ethylene in tomato fruit ripening and ripening-associated traits. New Phytol. 2020, 226, 460–475. [Google Scholar] [CrossRef]
- Gao, Y.; Wei, W.; Zhao, X.; Tan, X.; Fan, Z.; Zhang, Y.; Jing, Y.; Meng, L.; Zhu, B.; Zhu, H.; et al. A NAC transcription factor, NOR-like1, is a new positive regulator of tomato fruit ripening. Hortic. Res. 2018, 5, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Tavano, E.C.D.R.; Lammers, M.; Martinelli, A.P.; Angenent, G.C.; De Maagd, R.A. Re-evaluation of transcription factor function in tomato fruit development and ripening with CRISPR/Cas9-mutagenesis. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Lang, Z.; Wang, Y.; Tang, K.; Tang, D.; Datsenka, T.; Cheng, J.; Zhang, Y.; Handa, A.K.; Zhu, J.-K. Critical roles of DNA demethylation in the activation of ripening-induced genes and inhibition of ripening-repressed genes in tomato fruit. Proc. Natl. Acad. Sci. USA 2017, 114, E4511–E4519. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhu, G.; Li, R.; Yan, S.; Fu, D.; Zhu, B.; Tian, H.; Luo, Y.; Zhu, H. The RNA Editing Factor SlORRM4 Is Required for Normal Fruit Ripening in Tomato. Plant Physiol. 2017, 175, 1690–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. CRISPR/Cas9-mediated mutagenesis of lncRNA1459 alters tomato fruit ripening. Plant J. 2018, 94, 513–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uluisik, S.; Chapman, N.H.; Smith, R.; Poole, M.; Adams, G.; Gillis, R.B.; Besong, T.M.D.; Sheldon, J.; Stiegelmeyer, S.; Perez-Fons, L.; et al. Genetic improvement of tomato by targeted control of fruit softening. Nat. Biotechnol. 2016, 34, 950–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Samsulrizal, N.H.; Yan, C.; Allcock, N.S.; Craigon, J.; Blanco-Ulate, B.; Ortega-Salazar, I.; Marcus, S.E.; Bagheri, H.M.; Perez-Fons, L.; et al. Characterisation of CRISPR mutants targeting genes modulating pectin degradation in ripening tomato. Plant Physiol. 2018, 179, 544–557. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.-H.; Wang, B.; Li, N.; Tang, Y.; Yang, S.; Yang, T.; Xu, J.; Guo, C.; Yan, P.; Wang, Q.; et al. CRISPR/Cas9-induced Targeted Mutagenesis and Gene Replacement to Generate Long-shelf Life Tomato Lines. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Liu, J.; Chen, Y.; Tang, H.; Wang, Y.; He, Y.; Ou, Y.; Sun, X.; Wang, S.; Yao, Y. Tomato SlAN11 regulates flavonoid biosynthesis and seed dormancy by interaction with bHLH proteins but not with MYB proteins. Hortic. Res. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuluaga, D.L.; Gonzali, S.; Loreti, E.; Pucciariello, C.; Degl’Innocenti, E.; Guidi, L.; Alpi, A.; Perata, P. Arabidopsis thaliana MYB75/PAP1 transcription factor induces anthocyanin production in transgenic tomato plants. Funct. Plant Biol. 2008, 35, 606–618. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB–bHLH–WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Naves, E.R.; Silva, L.D.; Sulpice, R.; Araújo, W.; Nunes-Nesi, A.; Peres, L.E.; Zsögön, A. Capsaicinoids: Pungency beyond Capsicum. Trends Plant Sci. 2019, 24, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Martín-Pizarro, C.; Posé, D. Genome Editing as a Tool for Fruit Ripening Manipulation. Front. Plant Sci. 2018, 9, 1415. [Google Scholar] [CrossRef]
- Nugent, G.D.; Coyne, S.; Nguyen, T.T.; Kavanagh, T.A.; Dix, P.J. Nuclear and plastid transformation of Brassica oleracea var. botrytis (cauliflower) using PEG-mediated uptake of DNA into protoplasts. Plant Sci. 2006, 170, 135–142. [Google Scholar] [CrossRef]
- Nguyen, K.; Jordi, W.; Van Dun, K.; Schepers, F.; Davelaar, E.; Stoopen, G.; Dix, P.J.; Kane, E.J. Delayed Senescence in Cauliflower Transformed with an Autoregulated Isopentenyl Transferase Gene. Int. J. Plant Sci. 2008, 169, 339–347. [Google Scholar] [CrossRef]
- Kowalczyk, T.; Gerszberg, A.; Durańska, P.; Biłas, R.; Hnatuszko-Konka, K. High efficiency transformation of Brassica oleracea var. botrytis plants by Rhizobium rhizogenes. AMB Express 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Van Eck, J.; Li, L. Use of the cauliflower or gene for improving crop nutritional quality. Biotechnol. Annu. Rev. 2008, 14, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Lawrenson, T.; Shorinola, O.; Stacey, N.; Li, C.; Østergaard, L.; Patron, N.; Uauy, C.; Harwood, W. Induction of targeted, heritable mutations in barley and Brassica oleracea using RNA-guided Cas9 nuclease. Genome Biol. 2015, 16, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Liu, M.; Li, Q.; Si, J.; Ren, X.; Song, H. Efficient BoPDS Gene Editing in Cabbage by the CRISPR/Cas9 System. Hortic. Plant J. 2019, 5, 164–169. [Google Scholar] [CrossRef]
- Ma, C.; Zhu, C.; Zheng, M.; Liu, M.; Zhang, D.; Liu, B.; Li, Q.; Si, J.; Ren, X.; Song, H. CRISPR/Cas9-mediated multiple gene editing in Brassica oleracea var. capitata using the endogenous tRNA-processing system. Hortic. Res. 2019, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murovec, J.; Guček, K.; Bohanec, B.; Avbelj, M.; Jerala, R. DNA-Free Genome Editing of Brassica oleracea and B. rapa Protoplasts Using CRISPR-Cas9 Ribonucleoprotein Complexes. Front. Plant Sci. 2018, 9, 1594. [Google Scholar] [CrossRef] [PubMed]
- Schouten, H.J.; Tikunov, Y.; Verkerke, W.; Finkers, R.; Bovy, A.; Bai, Y.; Visser, R.G. Breeding Has Increased the Diversity of Cultivated Tomato in The Netherlands. Front. Plant Sci. 2019, 10, 1606. [Google Scholar] [CrossRef]
- Luby, J.J.; Shaw, D.V. Plant Breeders’ Perspectives on Improving Yield and Quality Traits in Horticultural Food Crops. HortScience 2009, 44, 20–22. [Google Scholar] [CrossRef]
- Van Vu, T.; Das, S.; Tran, M.T.; Hong, J.C.; Kim, J.-Y. Precision Genome Engineering for the Breeding of Tomatoes: Recent Progress and Future Perspectives. Front. Genome Ed. 2020, 2, 2. [Google Scholar] [CrossRef]
- Dias, J.S.; Ortiz, R. Transgenic Vegetable Breeding for Nutritional Quality and Health Benefits. Food Nutr. Sci. 2012, 3, 1209–1219. [Google Scholar] [CrossRef] [Green Version]
- Kato-Nitta, N.; Maeda, T.; Inagaki, Y.; Tachikawa, M. Expert and public perceptions of gene-edited crops: Attitude changes in relation to scientific knowledge. Palgrave Commun. 2019, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jorasch, P. Potential, Challenges, and Threats for the Application of New Breeding Techniques by the Private Plant Breeding Sector in the EU. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Jorasch, P. Will the EU stay out of step with science and the rest of the world on plant breeding innovation? Plant Cell Rep. 2019, 39, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Lassoued, R.; Phillips, P.W.; Macall, D.M.; Hesseln, H.; Smyth, S.J. Expert opinions on the regulation of plant genome editing. Plant Biotechnol. J. 2021, 19, 1104–1109. [Google Scholar] [CrossRef]
- Anders, S.; Cowling, W.; Pareek, A.; Gupta, K.J.; Singla-Pareek, S.L.; Foyer, C.H. Gaining Acceptance of Novel Plant Breeding Technologies. Trends Plant Sci. 2021, 26, 575–587. [Google Scholar] [CrossRef]
- Miladinovic, D.; Antunes, D.; Yildirim, K.; Bakhsh, A.; Cvejić, S.; Kondić-Špika, A.; Jeromela, A.M.; Opsahl-Sorteberg, H.-G.; Zambounis, A.; Hilioti, Z. Targeted plant improvement through genome editing: From laboratory to field. Plant Cell Rep. 2021, 40, 935–951. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Milestones in vegetable breeding focused on tomato and cauliflower.

Figure 2.
Examples of tomato diversity for SLL and SLC types on the top and on the bottom of the picture, respectively.
Figure 2.
Examples of tomato diversity for SLL and SLC types on the top and on the bottom of the picture, respectively.

Figure 3.
Examples of cauliflower types grouped by colour. From left to right and from top to bottom: green cauliflower, violet/purple cauliflower, green Romanesco cauliflower, orange Romanesco cauliflower, white and light yellow/pale cauliflower.
Figure 3.
Examples of cauliflower types grouped by colour. From left to right and from top to bottom: green cauliflower, violet/purple cauliflower, green Romanesco cauliflower, orange Romanesco cauliflower, white and light yellow/pale cauliflower.


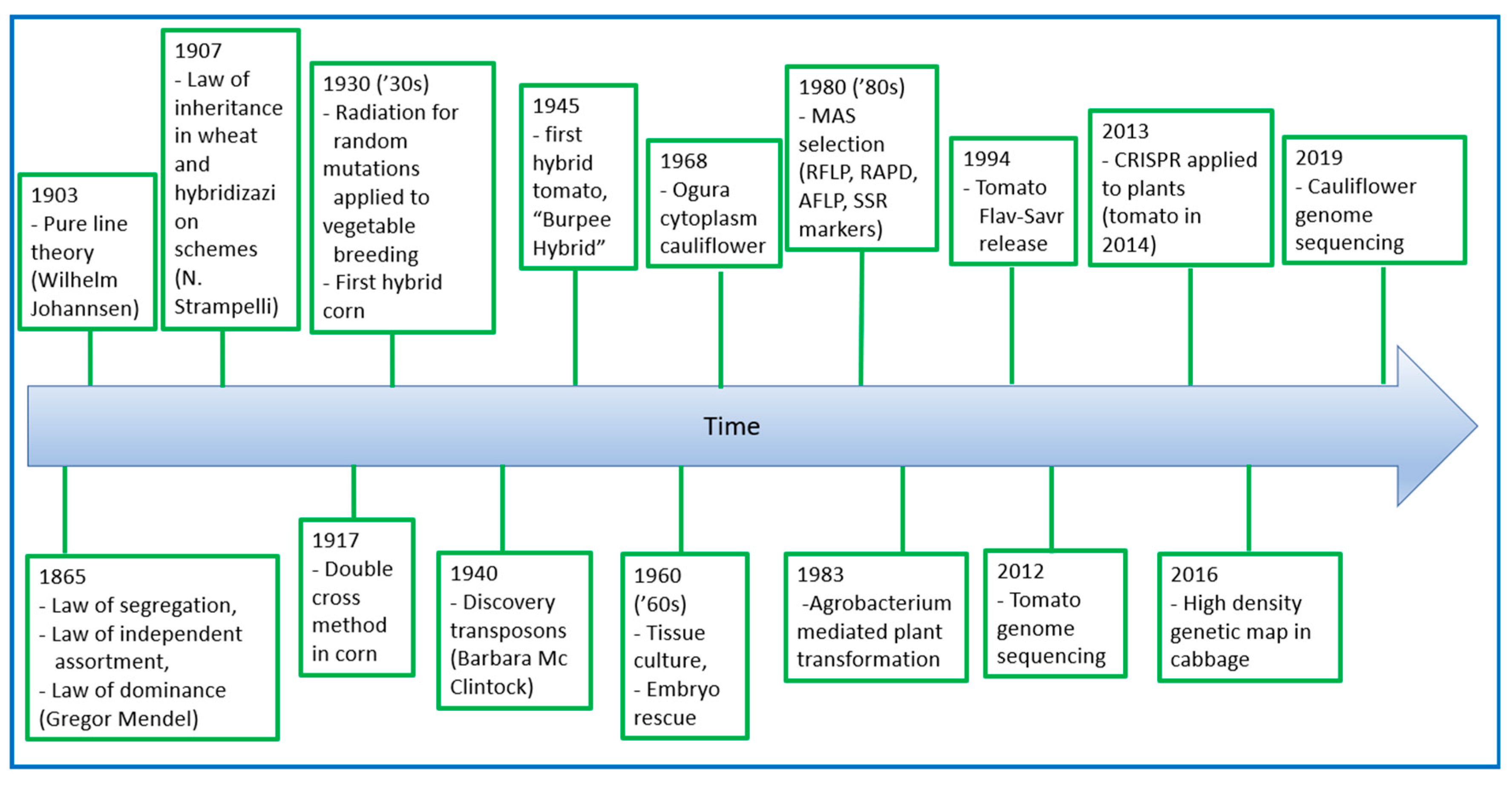{kind=link}
{kind=link}
{kind=link}
Table 1.
Information about some genome sequences available in tomato, cauliflower, and related species.
Table 1.
Information about some genome sequences available in tomato, cauliflower, and related species.
| Species | Accession/Cultivar/Landrace | Sequencing Technology | Genome Size | References |
|---|---|---|---|---|
| Solanum lycopersicum | Cultivar ‘Heinz 1706’ | Sanger and ‘next generation’ technologies’ (454/Roche GS FLX, SOLiD™ sequencing, Illumina sequencing) | 900 Mb | [28] |
| S. lycopericum | Varieties San Marzano and Vesuviano | Illumina HiSeq | 900 Mb | [63] |
| S. lycopersicum S. pimpinellifolium S. cheesmaniae, S. galapagense | Pan-genome of 725 accessions belonging to wild species, early domesticates, large-fruited heirlooms, modern elite cultivars and hybrids | Illumina NextSeq | Total of 306 Gb of contigs | [27] |
| S. pennellii | LA0716 | Illumina | 942 Mb | [88] |
| S. pimpinellifolium | LA2093 | Illumina | 923 Mb | [89] |
| S. pimpinellifolium | LA0480 | Illumina HiSeq2000 | 811 Mb | [90] |
| B. oleracea var. botrytis | Inbred line C-8 | PacBio and Illumina HiSeq X Ten | 584.60 Mb | [91] |
| B. rapa ssp. pekinensis | Line Chiifu-401-42 | Illumina GA II | 288 Mb | [92] |
| B. juncea var. tumida | Variety T84−66 | Illumina | 955 Mb | [93] |
| B. nigra | Double haploid line YZ12151 | Illumina | 591 Mb | [93] |
| B. napus | Cultivar Tapidor | Illumina | 635 Mb | [94] |
Table 2.
Examples of organoleptic and nutritional quality traits modified by genome editing in tomato.
Table 2.
Examples of organoleptic and nutritional quality traits modified by genome editing in tomato.
| Species | Traits | Metabolic Pathways | Target Genes | Genome Editing System | Transformation Technology | References |
|---|---|---|---|---|---|---|
| Solanum lycopersicum | Fruit colour | Carotenoids | SGR1, LCY-E, LCY-B1, LCY-B2, Blc | CRISPR/Cas9 | Agrobacterium tumefaciens-mediated | [110] |
| Carotenoids | Psy1 | CRISPR/Cas9 | A. tumefaciens-mediated | [111] | ||
| Flavonoids | MYB12 | CRISPR/Cas9 | A. tumefaciens-mediated | [112] | ||
| Anthocyanins | ANT1 | CRISPR/Cas9 | A. tumefaciens-mediated | [113] | ||
| GABA content in fruits | aminobutiric acid | GAD2, GAD3 | CRISPR/Cas9 | A. tumefaciens-mediated | [114] | |
| aminobutiric acid | GABA-TP1, GABA-TP2, GABA-TP3, CAT9, SSADH | CRISPR/Cas9 | A. tumefaciens-mediated | [115] | ||
| Metabolite content | SSC, fiber, fructose, ascorbic acid, total phenol, carotene, oxalic acid | L1L4 | ZFN | A. tumefaciens-mediated | [116] | |
| Ripening | Ripening regulatory system | RIN | CRISPR/Cas9 | A. tumefaciens | [117,118,119] | |
| Ripening regulatory system | NOR-like1 | CRISPR/Cas9 | A. tumefaciens-mediated | [120] | ||
| Ripening regulatory system | SBP-CNR, NAC-NOR | CRISPR/Cas9 | A. tumefaciens-mediated | [27] | ||
| Ripening regulatory system | AP2a, MAC-NOR, FUL1/TDR4, FUL2/MBP7 | CRISPR/Cas9 | A. tumefaciens-mediated | [121] | ||
| Ripening regulatory system | DML2 | CRISPR/Cas9 | A. tumefaciens-mediated | [122] | ||
| Ripening regulatory system | ORRM4 | CRISPR/Cas9 | A. tumefaciens-mediated | [123] | ||
| Ripening regulatory system | lncRNA1459 | CRISPR/Cas9 | A. tumefaciens-mediated | [124] | ||
| Fruit softening | PL | CRISPR/Cas9 | A. tumefaciens-mediated | [125] | ||
| Fruit softening | PL, PG2a, TBG4 | CRISPR/Cas9 | A. tumefaciens-mediated | [126] | ||
| Shelf life | Alc | CRISPR/Cas9 | A. tumefaciens-mediated | [127] | ||
| S. pimpinellifolium | Metabolite content | Lycopene | CycB | CRISPR/Cas9 | A. tumefaciens-mediated | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Natalini, A.; Acciarri, N.; Cardi, T. Breeding for Nutritional and Organoleptic Quality in Vegetable Crops: The Case of Tomato and Cauliflower. Agriculture 2021, 11, 606. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070606
AMA Style
Natalini A, Acciarri N, Cardi T. Breeding for Nutritional and Organoleptic Quality in Vegetable Crops: The Case of Tomato and Cauliflower. Agriculture. 2021; 11(7):606. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070606
Chicago/Turabian StyleNatalini, Alessandro, Nazzareno Acciarri, and Teodoro Cardi. 2021. "Breeding for Nutritional and Organoleptic Quality in Vegetable Crops: The Case of Tomato and Cauliflower" Agriculture 11, no. 7: 606. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070606
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.