
Establishment of a Bivector Genetic Transformation System in Recalcitrant Maize Inbred Lines


1
Shanghai Key Laboratory of Bio-Energy Crops, School of Life Sciences, Shanghai University, Shanghai 200444, China
2
State Key Laboratory of Plant Physiology and Biochemistry, National Maize Improvement Center, Beijing Key Laboratory of Crop Genetic Improvement, Joint International Research Laboratory of Crop Molecular Breeding, College of Agronomy and Biotechnology, China Agricultural University, Beijing 100193, China
*
Author to whom correspondence should be addressed.
Agriculture 2021, 11(7), 663; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070663
Submission received: 21 May 2021
/
Revised: 2 July 2021
/
Accepted: 12 July 2021
/
Published: 14 July 2021
Abstract
:Maize is an important grain crop with high nutritional value. An effective transformation system is crucial for the genetic improvement of maize traits, but many important maize inbred lines remained recalcitrant to transformation. In this study, we developed a bivector transformation system that worked well in two recalcitrant maize inbred lines. This system included an induction vector (ZmBBM-ZmWUS) and an indicator vector (GFP), using microprojectile bombardment technology combined with Agrobacterium-mediated transformation. We found that the Zheng58 and Mo17 recalcitrant inbred lines could be transformed with this system. The whole transformation cycle lasted only 52 days, 38 days less than the traditional transformation cycle. Additionally, it was possible to eliminate inference of the induction vector and obtained progenies with only the target gene. Our results suggested that the bivector system was an optimization of the current maize transformation methods and could potentially be used in genetic improvement of maize inbred lines.
1. Introduction
Maize is the most heavily produced cereal crop in the world [1]. It has high nutritional value [2]. The exploration of genetic transformation has contributed to the genetic improvement of maize inbred lines. Thus, establishing an efficient genetic transformation system is important for the improvement of maize.
Successful genetic transformation depends on the traits and genotypes of the maize inbred lines [3]. It was found that the Hi-II line produced calli and regenerated somatic embryos from immature embryos [4,5]. The Hi-II hybrid line is then widely utilized in commercial maize genetic transformation [3,4,6]. The regenerated maize A188, W117, W59E, A554, W153R’H99, HKI 163 and Mo17 were also developed from immature embryos through Agrobacterium tumefaciens-mediated mechanisms [7,8,9]. However, many maize inbred lines remained recalcitrant to transformation, such as Zheng58 and Chang 7–2.
In the past few years, many important maize inbred lines have been transformed effectively with morphogenetic genes. BABY BOOM (BBM) is a morphogenetic gene that promotes cell proliferation and morphogenesis during embryogenesis [10]. BBM is widely employed to obtain transgenic plants from tobacco (Nicotiana tabacum L.) [11], sweet pepper (C. annuum) varieties [12], and Theobroma cacao [13]. WUSCHEL (WUS) is the central meristem identity of shoot and floral meristems and plays an important role in maintaining structural and functional integrity [14]. WUS is widely utilized to obtain transgenic plants from tobacco [15], Coffea canephora [16], and cotton (Gossypium hirsutum L.) [17]. Co-expression of BBM/WUS can successfully induce transgenic plants from the maize B73 inbred line [18], sorghum (Sorghum bicolor) immature embryos [19], sugarcane (Saccharum officinarum) calli, and indica rice (Oryza sativa ssp indica) calli [20]. However, the strong expression of many morphogenetic genes often leads to the production of pleiotropic and harmful phenotypes, resulting in low fertility in transgenic T0 plants and subsequent offspring [21]. At present, effectively mitigating this situation is challenging.
This study reported the establishment of a bivector genetic transformation system for recalcitrant maize inbred lines. By employing Agrobacterium-mediated transformation and microprojectile bombardment technology, the bivector was transferred into immature embryos of recalcitrant maize inbred lines to successfully obtain transgenic plants. Our results showed that this system could successfully produce transgenic plants from the Zheng58 and Mo17 recalcitrant maize inbred lines with a significantly shorter transformation cycle. In addition, transgenic Zheng58 produced separated progenies without the existence of ZmBBM-ZmWUS morphogenetic genes.
2. Materials and Methods
2.1. Plant Materials
Four common inbred lines, namely, Zheng58, Mo17, Chang7-2 and B73, were maintained in the laboratory. After 14–16 days of pollination, when the young embryo grew to 1.8 mm–2.2 mm, the immature embryos of these inbred lines were harvested as acceptor materials for transformation. All plants were grown in the experimental field at the campus of Shanghai University.
2.2. Bivector Construction
The BABY BOOM (BBM), WUSCHEL (WUS) and Green fluorescent protein (GFP) genes were amplified using gene-specific primers. The maize Chitinase A1 promoter (CTA1pro), Nopaline synthase promoter (NOSpro), Auxin-inducible promoter (Axig1pro) and Phospholipid transferase protein promoter (PLTPpro) were reduplicated from the maize genome. The pHB vector was modified by our laboratory and stored long-term in our laboratory. The pHB vector contained the restriction sites EcoRI, HindIII, BamHI, and XbaI. The pCAMBIA3301 vector contained the PstI site. The bivector system contained an induction vector of pHB-ZmBBM-ZmWUS co-expression and an indicator vector of pCAMBIA3301-GFP (Table S1). The ZmCTA1pro::BBM cassette was ligated into the pMD18T vector to form the pMD18T-CB (pMD18T-ZmCTA1pro::BBM). Via the EcoRI and HindIII sites, the pMD18T-ZmCTA1pro::BBM vector was digested and ligated into the pHB vector to form the pHB-CB (pHB-ZmCTA1pro::BBM). The NOSpro::WUS cassette was ligated into the pMD18T vector to form the pMD18T-NW (pMD18T-NOSpro::WUS). Via the HindIII and BamHI sites, the pMD18T-NW (pMD18T-NOSpro::WUS) was digested and ligated into the pHB-CB vector to form the pHB-CB-NW (pHB-ZmCTA1pro::BBM-NOSpro::WUS). The Axig1pro::WUS cassette was ligated into the pMD18T vector to form the pMD18T-AW (pMD18T-Axig1pro::WUS). Via the HindIII and BamHI sites, the pMD18T-AW (pMD18T-Axig1pro::WUS) was digested and ligated into the pHB-CB vector to form the pHB-CB-AW (pHB-ZmCTA1pro::BBM- Axig1pro::WUS). The ZmPLTPpro::BBM cassette was ligated into the pMD18T vector to form the pMD18T-PB (pMD18T-ZmPLTPpro::BBM). Via the EcoRI and HindIII sites, the pMD18T-ZmPLTPpro::BBM vector was digested and ligated into the pHB vector to form the pHB-PB (pHB-ZmPLTPpro::BBM). Via the HindIII and XbaI sites, the pMD18T-AW (pMD18T-Axig1pro::WUS) was digested and ligated into the pHB-PB vector to form the pHB-PB-AW (pHB-ZmPLTPpro::BBM-Axig1pro::WUS). The GFP gene was recombined between the 35Spro promoter and the RbsT terminator into the pCAMBIA3301 vector to form the pCA-GFP (pCA-35Spro::GFP).
The linkage vectors contain ZmBBM, ZmWUS and GFP (Table S1). The pCA-GFP was recombined into the pHB-CB-NW (pHB-ZmCTA1pro::BBM-NOSpro::WUS) vector between the ZmCTA1pro::BBM cassette and NOSpro::WUS cassette to form the pHB-CB-NW-GFP vector (ZmCTA1pro::BBM-NOSpro::WUS-35Spro::GFP). The pCA-GFP was recombined into the pHB-CB-AW (pHB-ZmCTA1pro::BBM-Axig1pro::WUS) vector between the ZmCTA1pro::BBM cassette and Axig1pro::WUS cassette to form the pHB-CB-AW-GFP vector (ZmCTA1pro::BBM-Axig1pro::WUS-35Spro::GFP). The pCA-GFP was recombined into the pHB-PB-AW (pHB-ZmPLTPpro::BBM-Axig1pro::WUS) vector between the ZmPLTPpro::BBM cassette and Axig1pro::WUS cassette to form the pHB-PB-AW-GFP vector (ZmPLTPpro::BBM-Axig1pro::WUS-35Spro::GFP). Information of the primers is provided in Table S2.
2.3. Microprojectile Bombardment Technology
Plasmid DNAs are coated on the surface of tungsten powder. Under high pressure, coated plasmids were accelerated at high speed into specific maize tissues.
The pHB-CB-NW, pHB-CB-AW and pHB-PB-AW vectors were transformed into immature embryos of four inbred lines by microprojectile bombardment technology, according to El-Itriby et al. [22].
2.4. Agrobacterium-Mediated Transformation
The pHB-CB-NW-GFP, pHB-CB-AW-GFP, pHB-PB-AW-GFP, and pCA-GFP vectors were transformed into immature embryos of four inbred lines by Agrobacterium-mediated transformation. Agrobacterium-mediated transformation of maize immature embryos was performed, as described by Frame et al. [23].
2.5. Culture Media and Conditions for the Plant Genetic Transformation System
Immature embryos of Zheng58, Mo17, Chang7-2, and B73 inbred lines were placed at 4 °C overnight before transformation. After dissecting immature embryos, the embryos were transferred to hypertonic medium and cultured at 25 °C for 3 h. After the immature embryos were bombarded with the microprojectile bombardment technology, the embryos were cultured in hypertonic cultivation medium at 25 °C for 3 h. The immature embryos were transferred to IM medium and washed twice with 1 mL fresh IM medium. One milliliter of Agrobacterium tumefaciens liquid-infected immature embryos was mixed well in the dark for 5 min. The young embryos were transferred to cultivation medium to remove the excess bacterial liquid, their scutellas were kept upright, they were blown dry for 38 min in the super clean platform, and they were sealed into plates and cultured in the dark at 19 °C for 3 days. The immature embryos on cultivation medium were transferred to resting cultivation medium in the dark at 28 °C for 7 days. When calli formation occurred, the calli were transferred to pre-regeneration cultivation medium and cultured in the dark at 25 °C for 14 days. The calli were transferred from pre-regeneration cultivation medium to dark regeneration cultivation medium and later sealed with breathable film at 25 °C. The somatic embryos were transferred to light regeneration medium, sealed with light regeneration, and placed in a light incubator (25 °C, light intensity 80–100 me/m2/s, photoperiod 16:8). After somatic embryos grew into 3–5-cm seedlings, these embryos were placed in a light incubator with double distilled water.
After 2–3 days, the seedlings were transferred to small pots for culture (photoperiod 16:8, temperature 26 °C/22 °C (day/night), 350em-2s−1); approximately 10–14 days later, the seedlings identified as positive were transferred to a large pot for culture and to await pollination. After pollination, the seeds of the T1 generation were harvested over 35–40 days and dried in an oven at 30 °C. Hypertonic medium formulation was derived from El-Itriby et al. [22]. All other medium formulations were as described by Frame et al. [24].
2.6. Genomic DNA Extraction and PCR Assay
The hexadecyltrimethylammonium bromide method was used for genomic DNA extraction in the maize plants [25]. Through PCR, we determined whether the transgenic plants were positive. The existence of pHB vectors was determined by amplifying HPT, and the pCA-GFP vector was directly and specifically amplified with specific primers targeting GFP (Table S2). In general, the gene was amplified with Taq mix in a PCR thermocycler. The PCR products were electrophoresed in a 1% agarose gel with ethidium bromide and imaged by the Gel Doc XRS system (Bio-Rad). The sizes and locations of specific gene bands were compared to determine the existence of the relevant genes.
2.7. Southern Blot Analyses
To determine if transgenes were stably integrated in transgenic plants, Southern blotting was used after preliminary PCR analysis of the plants. For Southern blot hybridization, genomic DNA was extracted, digested, gel-fractionated, and transferred to nylon membranes, as described previously [26]. In the preparation of the molecular probe, primers from the pCA-GFP vector were used to amplify a PCR product as the template to generate a digoxigenin-dUTP-labeled probe (Table S2). Hybridization and signal detection were subsequently performed, utilizing the PCR DIG Probe Synthesis Kit and the DIG High Prime DNA Labeling and Detection Starter Kit II (Roche, Basel, Switzerland), according to the manufacturer’s instructions.
2.8. RNA Extraction and RT-qPCR Analysis
Total RNA of the maize tissues was extracted with TRIzol reagent (Tiangen, Beijing, China), according to a previously described protocol [27], and DNA was removed through treatment with RNase-Free DNase I (Takara, Shiga, Japan). Two micrograms of total RNA was reverse-transcribed into complementary DNA using ReverTra Ace reverse transcriptase (Toyobo, Osaka City, Japan). RT-qPCR primers were designed using the NCBI program (Premier Biosoft, Palo Alto, CA, USA).
RT-qPCR experiments were performed on three independent RNA sample sets, with Actin serving as an internal control gene. RT-qPCR was performed with SYBR Green Real-Time PCR Master Mix (Toyobo) using a Mastercycler ep realplex 2 (Eppendorf, Framingham, MA, USA), according to the standard protocol [28]. Relative quantifiable differences in gene expression were analyzed, as described previously [29].
3. Results
3.1. Bivector Construction for Genetic Transformation of Recalcitrant Maize Inbred Lines
Due to the harmful pleiotropic effects of morphogenetic genes [21], we constructed a bivector system containing an induction vector (ZmBBM-ZmWUS) and an indicator vector (GFP) to eliminate the influence of morphogenetic ZmBBM-ZmWUS in progeny generations.
We employed the strategies of calli-specific expression of ZmBBM and general expression of ZmWUS to construct the induction vector (ZmBBM-ZmWUS). Two promoters were used to drive ZmBBM. We found that, compared with the maize calli-specific Phospholipid transferase protein gene (ZmPLTP) [30], the maize Chitinase A1 gene (ZmCTA1) was also specifically expressed in calli but at a lower expression level (https://www.maizegdb.org/, accessed on 1 August 2019). Through reverse transcrition-quantitative PCR (RT-qPCR) analysis, we discovered that the ZmCTA1 and ZmPLTP gene had the highest expression in the calli followed by the silk, anther, and other tissues (such as root, stem, leaf, ear, immature seed, and embryo tissues) (Figure 1a,b). Therefore, we used ZmBBM driven by the promoter of the ZmCTA1 and ZmPLTP genes in the experimental group (ZmCTA1pro::BBM and ZmPLTPpro::BBM). Two promoters were used to drive ZmWUS. Compared with the maize auxin-inducible promoter (Axig1pro) [31], the Nopaline synthase promoter (NOSpro) [32] could produce a general expression without the induction of any hormone. Thus, we used ZmWUS driven by NOSpro and ZmAxig1pro in the experimental group (NOSpro::WUS and ZmAxig1pro::WUS).
The bivector system contained an induction vector of pHB-ZmBBM-ZmWUS co-expression and an indicator vector of pCAMBIA3301-GFP (Figure 2a–d). In the pHB-CB-NW vector, ZmCTA1pro drives ZmBBM and NOSpro drives ZmWUS (ZmCTA1pro::BBM-NOSpro::WUS) (Figure 2a). In the pHB-CB-AW vector, ZmCTA1pro drives ZmBBM, and ZmAxig1pro drives ZmWUS (ZmCTA1pro::BBM-Axig1pro::WUS) (Figure 2b). In the pHB-PB-AW vector, ZmPLTPpro drives ZmBBM and ZmAxig1pro drives ZmWUS (ZmPLTPpro::BBM-Axig1pro::WUS) (Figure 2c). The reporting pCAMBIA3301 vector (pCA-GFP), including Green fluorescent protein (GFP) [33], is that the cauliflower mosaic virus 35S promoter (35Spro) drives GFP (35Spro::GFP) (Figure 2d).
We also constructed linkage pHB vectors expressing ZmBBM, ZmWUS, and GFP as the pHB-CB-NW-GFP vector (ZmCTA1pro::BBM-NOSpro::WUS-35Spro::GFP) (Figure 2e), the pHB-CB-AW-GFP vector (ZmCTA1pro::BBM-Axig1pro::WUS-35Spro::GFP) (Figure 2f), and the pHB-PB-AW-GFP vector (ZmPLTPpro::BBM-Axig1pro::WUS-35Spro::GFP) (Figure 2g). These pHB-related vectors contain the Hygromycin phosphotransferase gene (HPT) [34] as the selectable marker gene of positive transgenic plants.
3.2. Culture Process of Genetic Transformation in Recalcitrant Maize Inbred Lines
For each of the four inbred lines (Zheng58, Mo17, Chang7-2 and B73), we designed three experimental groups for the bivector system test (pHB-NC+pCA-GFP or pHB-AC+pCA-GFP or pHB-AP+pCA-GFP), three experimental groups for the linkage vector test (pHB-NGC or pHB-AGC or pHB-AGP), and a control group (only pCA-GFP). So there were 24 experimental groups and 4 control groups (Table S3). Using microprojectile bombardment technology combined with Agrobacterium-mediated transformation, we successfully obtained transgenic Zheng58, Mo17 and Chang7-2 plants (Table 1). The target gene GFP was detected by amplifying a 410-bp band with specific primers (Table S2) in the transgenic T0 plants.
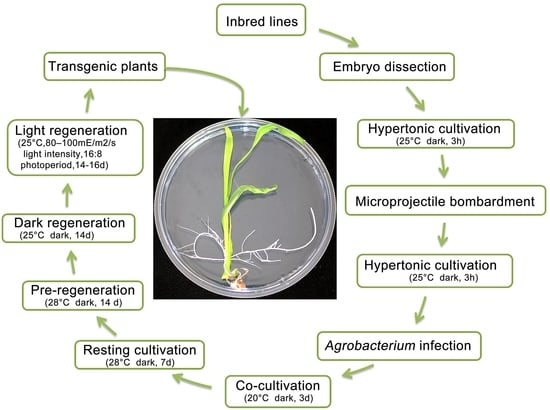
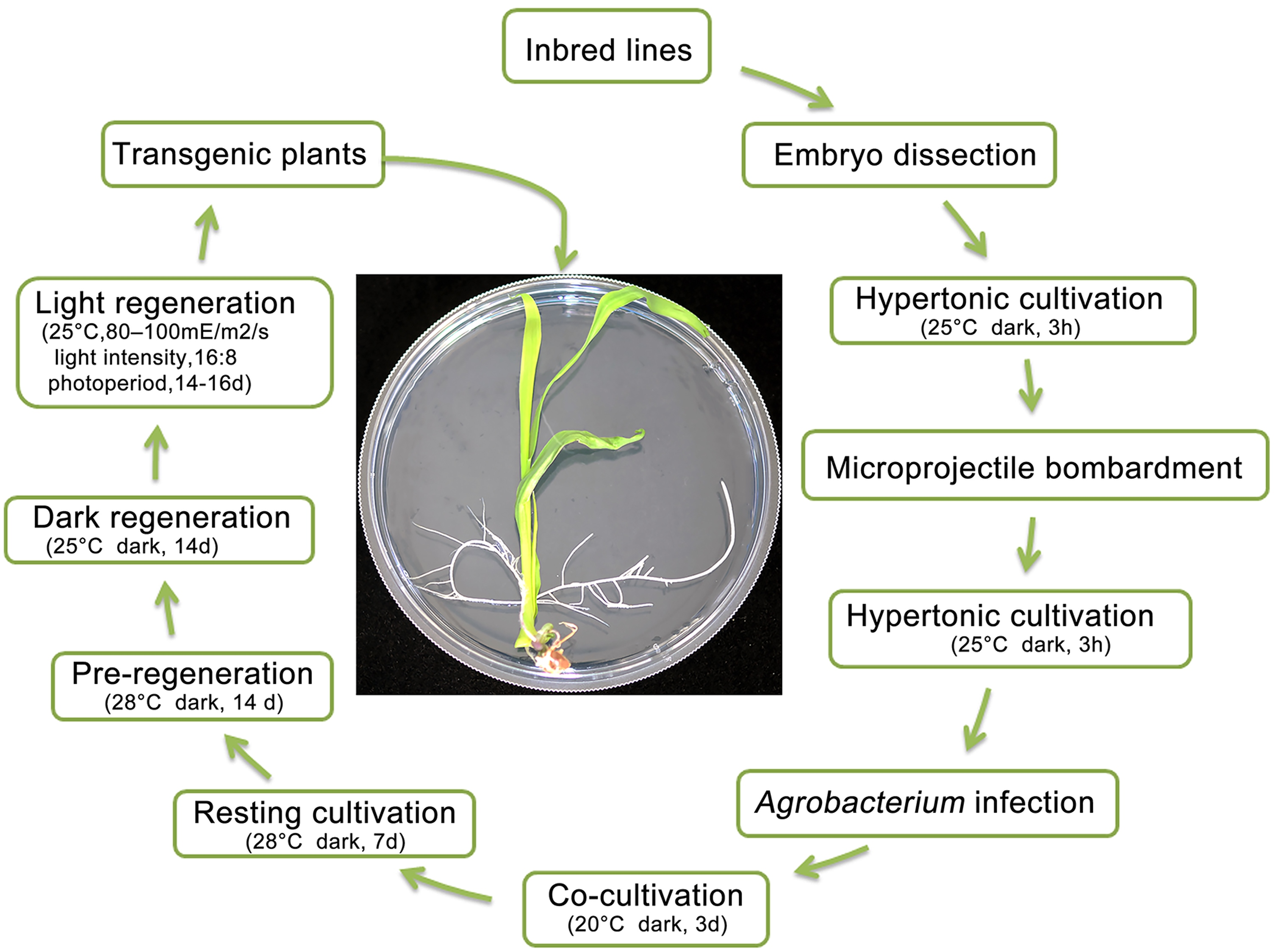
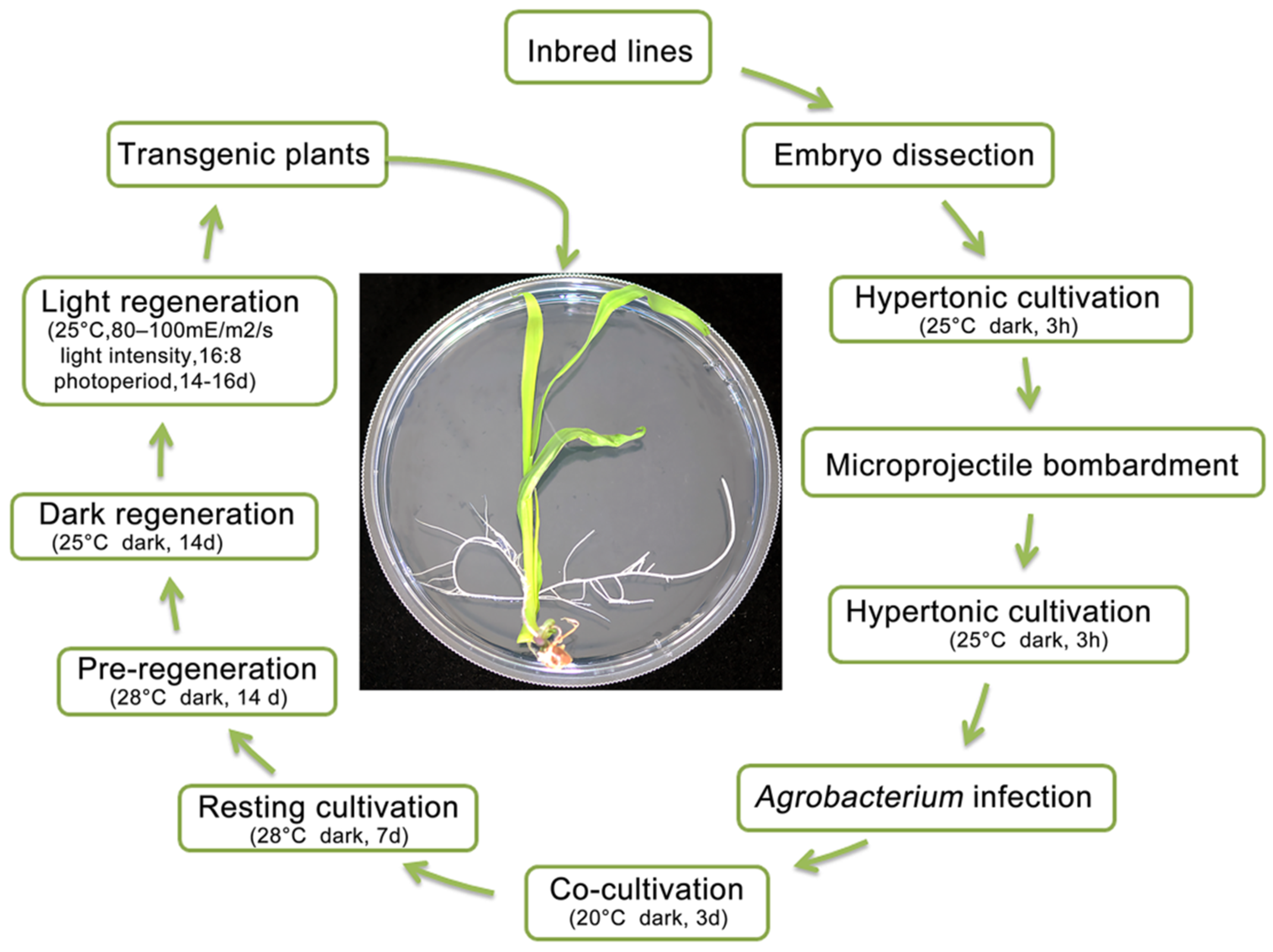
For the entire process of transformation culture, the transformation cycle for obtaining positive transgenic seedlings required approximately 52–66 days (Figure 3). In particular, we observed that the recalcitrant Zheng58 inbred line successfully produced transgenic plants after treatment with the bivector genetic transformation system, with the shortest period being 52 days, 38 days less than that required by traditional transformation [35]. Embryos were dissected from the seeds 14–16 days after self-pollination (DAP) of inbred lines. The length of immature embryos was 1.8–2.2 cm. After 3h in hypertonic cultivation medium, the pHB-CB-NW vector was transformed into immature embryos using microprojectile bombardment technology. After 3 h in hypertonic cultivation medium, the pCA-GFP vector was integrated into the maize genome via Agrobacterium infection. Transgenic embryo culture was undertaken in cocultivation medium for 3 days, resting cultivation medium for 7 days, no screening cultivation medium for 12 days (not a necessary step), pre-regeneration cultivation medium for 14 days (Figure 4a), dark regeneration cultivation medium for 14 days (Figure 4b), and light regeneration cultivation medium for 14–16 days (Figure 4c), and transgenic plants were grown in the experimental field (Figure 4d).
3.3. Transformation Frequencies of Maize Recalcitrant Inbred Lines
Three out of four inbred lines were able to produce transgenic plants (Zheng58, Mo17 and Chang7-2) (Table 1). Using the pHB-CB-NW vector and pCA-GFP vector, the recalcitrant inbred lines of Zheng58 successfully produced transgenic plants, with transformation ratios of 2.94%. With the pHB-CB-AW vector and pCA-GFP vector, transgenic plants were successfully obtained from the recalcitrant inbred lines Zheng58 and Mo17, with transformation ratios 0.14% and 0.66%, respectively. Via the linkage vector pHB-CB-AW-GFP, the recalcitrant inbred lines of Chang7-2 successfully produced transgenic plants, and the transformation ratios of Chang7-2 were 0.58%. Via the linkage vector pHB-PB-AW-GFP, the recalcitrant inbred line Mo17 successfully produced transgenic plants, and the transformation ratio of Mo17 was 1.57%.
Generally, we found that transgenic Zheng58, Mo17, and Chang7-2 all contained ZmCTA1pro::BBM. Furthermore, Zheng58 transformed with the bivector system, including ZmCTA1pro::BBM-NOSpro::WUS and 35Spro::GFP, exhibited the highest transformation ratio of all experimental groups (Table S3). Transformation frequencies [(number of T0 plants ÷ number of immature embryos) × 100] ranged from 0.14 to 2.94% (Table 1).
3.4. Progeny Analysis of Transgenic Zheng58 Plants
The transgenic Zheng58 plants carried the bivector system, including the induction vector (ZmCTA1pro::BBM-NOSpro::WUS) and the indicator vector (35Spro::GFP), exhibiting the highest transformation ratio. We identified the integration and expression of GFP in these positive transgenic lines of Zheng58. Via PCR, we detected the GFP gene by amplifying a 410-bp band with specific primers targeting GFP, and we detected ZmCTA1pro::BBM-NOSpro::WUS by amplifying a 546-bp band with specific primers targeting HPT (Table S2). By PCR identification, we detected both GFP and ZmCTA1pro::BBM-NOSpro::WUS in the 1#, 2#, 3#, and 4# transgenic lines (Figure 5a). Via Southern blot analysis, we detected the integrated copies of the GFP gene. The gel blot was probed with the DIG-labeled GFP cDNA fragment without the restriction enzyme sites of HindIII and EcoRI. By digesting the genome of the 1#, 2#, and 3# transgenic lines with HindIII (Figure 5b) and EcoRI (Figure 5c), the copy number of GFP was three, three, and one, respectively. RT-qPCR analysis showed that the GFP gene expressed in the 1#, 2#, 3#, and 4# transgenic lines (Figure 5d).
3.5. Progeny Segregation Analysis of Transgenic Zheng58 Lines
For the progeny seeds of lines 1#, 2#, 3#, and 4# of transgenic Zheng58 (T2-1#, T2-2#, T2-3# and T2-4#), we detected the target gene GFP by amplifying a 410-bp band with primers specific for GFP, and we detected ZmCTA1pro::BBM-NOSpro::WUS by amplifying a 546-bp band with specific primers targeting HPT (Figure 6, Table S2). The results showed that we obtained separated progenies only with the presence of GFP but not with ZmCTA1pro::BBM-NOSpro::WUS. In T2-1#, T2-2#, T2-3#, T2-4#, the numbers of segregated seeds were four, three, three, and three, respectively. Further, statistical analysis showed that the segregation ratio [(number of separated seeds ÷ number of T2 seeds) × 100] was in the range of 2.03 to 6.78% (Table 2).
4. Discussion
Maize plays a vital role as a food and feed crop worldwide [36]. Genetic transformation of many important maize inbred lines remains a challenge. Many recalcitrant maize inbred lines are still difficult to be transformed [21]. Enhanced maize genetic transformation procedures would help us to improve recalcitrant inbred lines. Therefore, we created a bivector genetic transformation system.
With the bivector system, we transferred ZmBBM-ZmWUS by microprojectile bombardment and transferred GFP via Agrobacterium-mediated transformation into maize immature embryos. Constitutive expression of ZmBBM and ZmWUS has severe pleiotropic effects, such as wrinkled leaves [37] and root thickening [10]. To reduce those effects on offsprings, we bombarded immature embryos with microprojectile bombardment technology to make induction genes express instantaneously. We used Agrobacterium-mediated transformation to infect immature embryos to ensure stable expression of indicator gene. After cultivation, the Zheng58 and Mo17 maize inbred lines yielded transgenic plants. In all experimental groups, the highest transformation frequencies were 2.94% for transgenic Zheng58, which were induced by ZmCTA1pro::BBM-NOSpro::WUS of the bivector genetic transformation system to successfully obtain transgenic plants. Unfortunately, the induction genes were also stably transformed but not instantaneously expressed (Figure 5). The different genotypes affect somatic embryogenesis [38]. The transformation ability of maize is controlled by nuclear genes [39]. At least one gene or gene cluster affects the formation of somatic embryos in tissue culture [40]. The Zheng 58 inbred line is not only tolerant of close planting, fertilizer, drought, lodging, and bacterial resistance, but is also the most widely used inbred line and exhibits a high yield [41]. Recalcitrant Zheng58 inbred lines successfully obtained transgenic plants by the bivector genetic transformation system, which may establish a foundation for applications in agriculture and commerce.
Expression of morphogenetic genes provides a reliable method for producing fertile transgenic plants. The whole process involves a lot of induction genes, such as using the combination of embryonic regulatory factors and meristem regulatory factors to enhance embryonic response [21]. In our study, we used the embryonic regulatory factor of ZmBBM and the meristem regulatory factor of ZmWUS as the induction genes. Under the expression of ZmBBM and ZmWUS, embryo formation was accelerated and the transformation cycle was shortened. The whole transformation cycle was 52–66 days for the transformation of Zheng58 with the bivector system, which was 38 days less than the period required by traditional transformation. In the traditional transformation cycle, A188, A634, H99, and W117 inbred lines could effectively produce transgenic plants by Agrobacterium tumefaciens-mediated procedures, and the transformation cycle was approximately 90 days [35]. Recently, using Agrobacterium- and biolistic-mediated methods, immature embryos of maize inbred B104 were observed to produce transgenic plants within 112–147 days [42]. The establishment of a bivector regeneration system considerably reduces the time of the transformation cycle.
It has been reported that ectopic expression of morphogenetic genes presents a harmful pleiotropic phenotype [21]. To eliminate the pleiotropic effect caused by continuous expression of BBM, heat shock-induced FLP gene expression was used to remove the BBM gene sequence between two FRT sites, thereby resulting in normal transgenic plants of Chinese white poplar (Populus tomentosa Carr.) [43]. In our study, the expression of BBM under the control of ZmCTA1pro was weaker than that of ZmPLTPpro, and the expression of ZmWUS was driven by ZmNospro. The expression pattern of ZmCTA1pro::BBM-Nospro::WUS was beneficial for eliminating the pleiotropic effect of morphogenetic genes. Utilizing the strengths of the bivector genetic transformation system, we could separate GFP and ZmBBM-ZmWUS. We detected only GFP but not ZmBBM-ZmWUS in T2 generation seeds, effectively mitigating the impact of ZmBBM-ZmWUS. High-copy-number transgenes are associated with functional and structural instability [44,45]. Based on the T2 generation seeds of transgenic Zheng58, the progeny segregation ratio was low. This low ratio might be related to the high-copy phenomenon.
Our research provided new ideas for the genetic transformation of maize and established a foundation for further research attempting to develop better maize lines and promote the development of maize breeding.
5. Conclusions
This study is the first attempt to establish a bivector genetic transformation system, which was successfully employed to obtain transgenic Zheng58 plants. This system not only provides a means of genetically transforming more recalcitrant maize inbred lines, but may also significantly reduce the time required by the transformation cycle. In addition, this system was able to eliminate the pleiotropic effects of ZmBBM-ZmWUS. This bivector maize genetic transformation system would contribute to the improvement of maize genetic breeding.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/agriculture11070663/s1, Table S1: The construction of the vectors. Table S2: Primers used in this study; Table S3: Summary data for Transformation Ratio of all trial groups.
Author Contributions
R.S. and W.Q. conceived and designed research. Y.G. and X.C. performed experiments. Y.G. collected and analyzed the data. Y.G. wrote the manuscript; R.S. and W.Q. edited and supported suggestions for the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (grants 31730065 and 31425019 to R.S.), the National Key Research and Development Program of China (grant 2016YFD0100503 to W.Q.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting this study’s findings are available by fair request from the corresponding author.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grants 31730065 and 31425019 to R.S.), the National Key Research and Development Program of China (grant 2016YFD0100503 to W.Q.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yao, D.; Qi, W.; Li, X.; Yang, Q.; Yan, S.; Ling, H.; Wang, D.; Wang, G.; Song, R. Maize opaque10 encodes a cereal-specific protein that is essential for the proper distribution of zeins in en-dosperm protein bodies. PLoS Genet. 2016, 12, e1006270. [Google Scholar] [CrossRef]
- Ishida, Y.; Hiei, Y.; Komari, T. Tissue culture protocols for gene transfer and editing in maize (Zea mays L.). Plant Biotechnol. 2020, 37, 121–128. [Google Scholar] [CrossRef]
- Yadava, P.; Abhishek, A.; Singh, R.; Singh, I.; Kaul, T.; Pattanayak, A.; Agrawal, P.K. Advances in maize transformation technologies and development of transgenic maize. Front. Plant Sci. 2017, 7, 1949. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, C.L.; Green, C.E. Establishment and maintenance of friable, embryogenic maize callus and the involvement of L-proline. Planta 1985, 164, 207–214. [Google Scholar] [CrossRef]
- Armstrong, C.L.; Green, C.E.; Phillips, R. Development and availability of germplasm with high Type II culture formation response. Maize Genet. Coop. News Lett. 1991, 65, 92–93. [Google Scholar]
- Vega, J.; Yu, W.; Kennon, A.R.; Chen, X.; Zhang, Z.J. Improvement of agrobacterium-mediated transformation in HI-II maize (Zea mays) using standard binary vectors. Plant Cell Rep. 2007, 27, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Saito, H.; Ohta, S.; Hiei, Y.; Komari, T.; Kumashiro, T. High efficiency transformation of maize (Zea mays L.) mediated by agrobacterium tumefaciens. Nat. Biotechnol. 1996, 14, 745–750. [Google Scholar] [CrossRef]
- Abhishek, A.; Kumari, R.; Karjagi, C.G.; Kumar, P.; Kumar, B.; Dass, S.; Kumar, R.S.; Ramteke, P.W. Tissue culture independent agrobacterium tumefaciens mediated in planta transformation method for tropical maize (Zea mays L.). Proc. Natl. Acad. Sci. India Sect. B Boil. Sci. 2016, 86, 375–384. [Google Scholar] [CrossRef]
- Jones, T.; Lowe, K.; Hoerster, G.; Anand, A.; Wu, E.; Wang, N.; Arling, M.; Lenderts, B. Maize transformation using the morphogenic genes baby boom and wuschel2. In Transgenic Plants: Methods and Protocols; Kumar, S., Barone, P., Smith, M., Eds.; Springer: New York, NY, USA, 2019; pp. 81–93. [Google Scholar]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.-M.; van Lammeren, A.A.M.; Miki, B.L.A.; et al. Ectopic expression of baby boom triggers a conversion from vegetative to embryonic growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, C.; Liu, Z.; Heidmann, I.; Supena, E.D.J.; Fukuoka, H.; Joosen, R.; Lambalk, J.; Angenent, G.; Scorza, R.; Custers, J.B.M.; et al. Heterologous expression of the baby boom AP2/ERF transcription factor enhances the regeneration capacity of tobacco (Nicotiana tabacum L.). Planta 2006, 225, 341–351. [Google Scholar] [CrossRef]
- Heidmann, I.; De Lange, B.; Lambalk, J.; Angenent, G.C.; Boutilier, K. Efficient sweet pepper transformation mediated by the BABY BOOM transcription factor. Plant Cell Rep. 2011, 30, 1107–1115. [Google Scholar] [CrossRef] [Green Version]
- Florez, S.L.; Erwin, R.L.; Maximova, S.N.; Guiltinan, M.J.; Curtis, W.R. Enhanced somatic embryogenesis in Theobroma cacao using the homologous baby boom transcription factor. BMC Plant Biol. 2015, 15, 121. [Google Scholar] [CrossRef] [Green Version]
- Laux, T.; Mayer, K.; Berger, J.; Jurgens, G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [CrossRef]
- Rashid, S.Z.; Yamaji, N.; Kyo, M. Shoot formation from root tip region: A developmental alteration by WUS in transgenic tobacco. Plant Cell Rep. 2007, 26, 1449–1455. [Google Scholar] [CrossRef]
- Arroyo-Herrera, A.; Gonzalez, A.K.; Moo, R.C.; Quiroz-Figueroa, F.R.; Loyola-Vargas, V.M.; Rodriguez-Zapata, L.; D′hondt, C.B.; Suárez-Solís, V.M.; Castaño, E. Expression of WUSCHEL in Coffea canephora causes ectopic morphogenesis and increases somatic embryogenesis. Plant Cell Tissue Organ. Cult. 2008, 94, 171–180. [Google Scholar] [CrossRef]
- Bouchabké-Coussa, O.; Obellianne, M.; Linderme, D.; Montes, E.; Maia-Grondard, A.; Vilaine, F.; Pannetier, C. Wuschel overexpression promotes somatic embryogenesis and induces organogenesis in cotton (Gossypium hirsutum L.) tissues cultured in vitro. Plant Cell Rep. 2013, 32, 675–686. [Google Scholar] [CrossRef]
- Mookkan, M.; Nelson-Vasilchik, K.; Hague, J.; Kausch, A.; Zhang, Z.J. morphogenic regulator-mediated transformation of maize inbred B. Curr. Protoc. Plant Biol. 2018, 3, e20075. [Google Scholar] [CrossRef]
- Mookkan, M.; Nelson-Vasilchik, K.; Hague, J.; Zhang, Z.J.; Kausch, A.P. Selectable marker independent transformation of recalcitrant maize inbred B73 and sorghum P898012 mediated by morphogenic regulators baby boom and WUSCHEL. Plant Cell Rep. 2017, 36, 1477–1491. [Google Scholar] [CrossRef] [Green Version]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.-J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic regulators Baby boom and WUSCHEL improve monocot transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef] [Green Version]
- Gordon-Kamm, B.; Sardesai, N.; Arling, M.; Lowe, K.; Hoerster, G.; Betts, S.; Jones, A.T. Using morphogenic genes to improve recovery and regeneration of transgenic plants. Plants 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- El-Itriby, H.A.; Assem, S.K.; Hussein, E.H.A.; Abdel-Calil, F.M.; Madkour, M.A. Regeneration and transformation of Egyptian maize inbred lines via immature embryo culture and a biolistic particle delivery system. Vitr. Cell. Dev. Biol. Anim. 2003, 39, 524–531. [Google Scholar] [CrossRef]
- Frame, B.R.; Shou, H.; Chikwamba, R.K.; Zhang, Z.; Xiang, C.; Fonger, T.M.; Pegg, S.E.K.; Li, B.; Nettleton, D.S.; Pei, D.; et al. Agrobacterium tumefaciens-mediated transformation of maize embryos using a standard binary vector system. Plant Physiol. 2002, 129, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Frame, B.; Main, M.; Schick, R.; Wang, K. Genetic transformation using maize immature zygotic embryos. Methods Mol. Biol. 2010, 710, 327–341. [Google Scholar] [CrossRef]
- Gao, H.; Smith, J.; Yang, M.; Jones, S.; Djukanovic, V.; Nicholson, M.G.; West, A.; Bidney, D.; Falco, S.C.; Jantz, D.; et al. Heritable targeted mutagenesis in maize using a designed endonuclease. Plant J. 2009, 61, 176–187. [Google Scholar] [CrossRef]
- Guan, Z.; Meng, X.; Sun, Z.; Xu, Z.; Song, R. Characterization of duplicated Dunaliella viridis SPT1 genes provides insights into early gene divergence after duplication. Gene 2008, 423, 36–42. [Google Scholar] [CrossRef]
- Wang, G.; Wang, G.; Zhang, X.; Wang, F.; Song, R. Isolation of high quality RNA from cereal seeds containing high levels of starch. Phytochem. Anal. 2012, 23, 159–163. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(–delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Anand, A.; Arling, M.L.; da Silva Conceicao, A.; Gordon-Kamm, W.J.; Hastings, C.E.; Hoerster, G.M.; Klein, T.M.; La Rota, C.M.; Lowe, K.S.; Tiwari, S.B. et al. Methods and Compositions for Rapid Plant Transformation. U.S. Patent US20170121722A1, 4 May 2017. [Google Scholar]
- Garnaat, C.; Lowe, K.S.; Roth, B. Zmaxig1-Specific Polynucleotides and Methods of Use. U.S. Patent WO2002006499A3, 13 July 2001. [Google Scholar]
- An, G. Development of plant promoter expression vectors and their use for analysis of differential activity of nopaline synthase promoter in transformed tobacco cells. Plant Physiol. 1986, 81, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Chalfie, M.; Tu, Y.; Euskirchen, G.; Ward, W.; Prasher, D. Green fluorescent protein as a marker for gene expression. Science 1994, 263, 802–805. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.A.; Vetsch, C.S.; Potts, D.E.; Lundquist, R.C. Transformation and inheritance of a hygromycin phosphotransferase gene in maize plants. Plant Mol. Biol. 1992, 18, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Hiei, Y.; Komari, T. Agrobacterium-mediated transformation of maize. Nat. Protoc. 2007, 2, 1614–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenhard, M.; Jürgens, G.; Laux, T. The WUSCHEL and SHOOTMERISTEMLESS genes fulfil complementary roles in Ara-bidopsis shoot meristem regulation. Development 2002, 129, 3195–3206. [Google Scholar] [CrossRef]
- Bohorova, N.; Luna, B.; Brito, R.M.; Huerta, L.D. Regeneration potential of tropical, subtropical, midaltitude, and highland maize inbreds. Maydica 1995, 40, 275–281. [Google Scholar]
- Tomes, D.T.; Smith, O.S. The effect of parental genotype on initiation of embryogenic callus from elite maize (Zea mays L.) germplasm. Theor. Appl. Genet. 1985, 70, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Willman, M.R.; Schroll, S.M.; Hodges, T.K. Inheritance of somatic embryogenesis and plantlet regeneration from primary (type 1) callus in maize. Vitr. Cell. Dev. Biol. Anim. 1989, 25, 95–100. [Google Scholar] [CrossRef]
- Raihan, M.S.; Liu, J.; Huang, J.; Guo, H.; Pan, Q.; Yan, J. Multi-environment QTL analysis of grain morphology traits and fine mapping of a kernel-width QTL in Zheng58 × SK maize population. Theor. Appl. Genet. 2016, 129, 1465–1477. [Google Scholar] [CrossRef]
- Raji, J.A.; Frame, B.; Little, D.; Santoso, T.J.; Wang, K.; Lagrimini, L.M. Agrobacterium- and biolistic-mediated transformation of maize b104 inbred. Adv. Struct. Saf. Stud. 2018, 1676, 15–40. [Google Scholar] [CrossRef]
- Deng, W.; Luo, K.; Li, Z.; Yang, Y. A novel method for induction of plant regeneration via somatic embryogenesis. Plant Sci. 2009, 177, 43–48. [Google Scholar] [CrossRef]
- Meyer, P.; Saedler, H. Homology-dependent gene silencing in plants. Annu. Rev. Plant Biol. 1996, 47, 23–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhi, L.; TeRonde, S.; Meyer, S.; Arling, M.L.; Iii, J.C.R.; Zhao, Z.-Y.; Jones, T.J.; Anand, A. Effect of Agrobacterium strain and plasmid copy number on transformation frequency, event quality and usable event quality in an elite maize cultivar. Plant Cell Rep. 2015, 34, 745–754. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Gene expression level of ZmCTA1 and ZmPLTP in various maize tissues. (a,b) RT-qPCR confirmed the expression level of ZmCTA1 and ZmPLTP in the root, stem, leaf, silk, ear, anther, immature seed, embryo and calli of maize. Actin was used as an internal control. Date are the mean values with standard error of mean (SEM), n = 3 individuals.
Figure 1.
Gene expression level of ZmCTA1 and ZmPLTP in various maize tissues. (a,b) RT-qPCR confirmed the expression level of ZmCTA1 and ZmPLTP in the root, stem, leaf, silk, ear, anther, immature seed, embryo and calli of maize. Actin was used as an internal control. Date are the mean values with standard error of mean (SEM), n = 3 individuals.

Figure 2.
Vector maps for the genetic transformation system. BBM, BABY BOOM; WUS, WUSCHEL; GFP, Green fluorescent protein; NOSpro, Nopaline synthase promoter; CTA1pro, maize Chitinase A1 promoter; Axig1pro, Auxin-inducible promoter; PLTPpro, Phospholipid transferase protein promoter; 35Spro, the cauliflower mosaic virus 35S promoter; TNOS, Nopaline synthase terminator; TRbst, Rbst terminator; T35S, cauliflower mosaic virus 35S terminator; BAR, Bialaphos resistance; HPT, Hygromycin phosphotransferase; LB, left border; RB, right border. A summary of the primers is given in Table S2. (a) T-DNA composition for the pHB-CB-NW vector. This vector shows that ZmCTA1pro drives ZmBBM and that NOSpro drives ZmWUS. (b) T-DNA composition for the pHB-CB-AW vector. In this vector, ZmCTA1pro drives ZmBBM and ZmAxig1pro drives ZmWUS. (c) T-DNA composition for the pHB-PB-AW vector. This vector shows that ZmPLTPpro drives ZmBBM and that ZmAxig1pro drives ZmWUS. (d) T-DNA composition for the pCA-GFP vector. This vector is a directive reporting pCAMBIA3301 vector including GFP driven by the 35S promoter (35Spro::GFP). (e) T-DNA composition for the pHB-CB-NW-GFP vector. In this vector, ZmCTA1pro drives ZmBBM, 35Spro drives GFP, and NOSpro drives ZmWUS. (f) T-DNA composition for the pHB-CB-NW-GFP vector. In the pHB-CGA vector, ZmCTA1pro drives ZmBBM, 35Spro drives GFP, and ZmAxig1pro drives ZmWUS. (g) T-DNA composition for the pHB-PB-AW-GFP vector. This vector shows that PLTPpro drives ZmBBM, 35Spro drives GFP, and ZmAxig1pro drives ZmWUS.
Figure 2.
Vector maps for the genetic transformation system. BBM, BABY BOOM; WUS, WUSCHEL; GFP, Green fluorescent protein; NOSpro, Nopaline synthase promoter; CTA1pro, maize Chitinase A1 promoter; Axig1pro, Auxin-inducible promoter; PLTPpro, Phospholipid transferase protein promoter; 35Spro, the cauliflower mosaic virus 35S promoter; TNOS, Nopaline synthase terminator; TRbst, Rbst terminator; T35S, cauliflower mosaic virus 35S terminator; BAR, Bialaphos resistance; HPT, Hygromycin phosphotransferase; LB, left border; RB, right border. A summary of the primers is given in Table S2. (a) T-DNA composition for the pHB-CB-NW vector. This vector shows that ZmCTA1pro drives ZmBBM and that NOSpro drives ZmWUS. (b) T-DNA composition for the pHB-CB-AW vector. In this vector, ZmCTA1pro drives ZmBBM and ZmAxig1pro drives ZmWUS. (c) T-DNA composition for the pHB-PB-AW vector. This vector shows that ZmPLTPpro drives ZmBBM and that ZmAxig1pro drives ZmWUS. (d) T-DNA composition for the pCA-GFP vector. This vector is a directive reporting pCAMBIA3301 vector including GFP driven by the 35S promoter (35Spro::GFP). (e) T-DNA composition for the pHB-CB-NW-GFP vector. In this vector, ZmCTA1pro drives ZmBBM, 35Spro drives GFP, and NOSpro drives ZmWUS. (f) T-DNA composition for the pHB-CB-NW-GFP vector. In the pHB-CGA vector, ZmCTA1pro drives ZmBBM, 35Spro drives GFP, and ZmAxig1pro drives ZmWUS. (g) T-DNA composition for the pHB-PB-AW-GFP vector. This vector shows that PLTPpro drives ZmBBM, 35Spro drives GFP, and ZmAxig1pro drives ZmWUS.

Figure 3.
Culture process of the bivector maize genetic transformation system. Embryos were isolated from the seeds of the Zheng58 inbred line at 14–16 DAP. The length of immature embryos was 1.8–2.2 cm. The pHB-CB-NW vector was transformed into immature embryos by microprojectile bombardment technology. The pCA-GFP vector was integrated into the maize genome via Agrobacterium infection. There are a series of culture media and conditions to obtain transgenic plants.
Figure 3.
Culture process of the bivector maize genetic transformation system. Embryos were isolated from the seeds of the Zheng58 inbred line at 14–16 DAP. The length of immature embryos was 1.8–2.2 cm. The pHB-CB-NW vector was transformed into immature embryos by microprojectile bombardment technology. The pCA-GFP vector was integrated into the maize genome via Agrobacterium infection. There are a series of culture media and conditions to obtain transgenic plants.

Figure 4.
Development of transgenic plants. (a) The calli were sprouted on the pre-regeneration medium. (b) Most embryos initially induced roots on dark regeneration medium. (c) Embryos absorbed light to grow plants on light regeneration medium and were cultured in growth chambers. (d) Transgenic plants were successfully obtained from immature maize embryos of Zheng58 in a greenhouse.
Figure 4.
Development of transgenic plants. (a) The calli were sprouted on the pre-regeneration medium. (b) Most embryos initially induced roots on dark regeneration medium. (c) Embryos absorbed light to grow plants on light regeneration medium and were cultured in growth chambers. (d) Transgenic plants were successfully obtained from immature maize embryos of Zheng58 in a greenhouse.

Figure 5.
Progeny analysis of transgenic Zheng58 plants. Zheng58 (Z58) served as a negative control. (a) PCR identification concerning the 4 lines of transgenic Zheng58. The plasmid of the pCA-GFP vector was treated as a positive control. (b) Genomic DNA of transgenic Zheng58 plants digested with HindIII for Southern blot analysis. (c) Genomic DNA of transgenic Zheng58 plants digested with EcoRI for Southern blot analysis. Red arrows identify the gene copies. (d) Comparison of the GFP expression level in the leaf tissue of Z58 and transgenic Zheng58. The transcript levels were normalized according to Actin. Values are the means with SEM; n = 3 individuals. See Table S2 for primers.
Figure 5.
Progeny analysis of transgenic Zheng58 plants. Zheng58 (Z58) served as a negative control. (a) PCR identification concerning the 4 lines of transgenic Zheng58. The plasmid of the pCA-GFP vector was treated as a positive control. (b) Genomic DNA of transgenic Zheng58 plants digested with HindIII for Southern blot analysis. (c) Genomic DNA of transgenic Zheng58 plants digested with EcoRI for Southern blot analysis. Red arrows identify the gene copies. (d) Comparison of the GFP expression level in the leaf tissue of Z58 and transgenic Zheng58. The transcript levels were normalized according to Actin. Values are the means with SEM; n = 3 individuals. See Table S2 for primers.

Figure 6.
Progeny segregation of the transgenic Zheng58 lines. Progeny transgenic Zheng58 of lines 1#, 2#, 3#, and 4# contained ZmCTA1pro::BBM-NOSpro::WUS or 35Spro::GFP. Seeds were recognized by amplifying GFP and HPT in the descendants of each line. The pCA-GFP vector and pHB vector were treated as positive controls. Zheng58 (Z58) was served as a negative control. See Table S2 for primers.
Figure 6.
Progeny segregation of the transgenic Zheng58 lines. Progeny transgenic Zheng58 of lines 1#, 2#, 3#, and 4# contained ZmCTA1pro::BBM-NOSpro::WUS or 35Spro::GFP. Seeds were recognized by amplifying GFP and HPT in the descendants of each line. The pCA-GFP vector and pHB vector were treated as positive controls. Zheng58 (Z58) was served as a negative control. See Table S2 for primers.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Transformation ratio of the recalcitrant maize inbred lines.
| Maize Inbred Lines | Vector | Number of Immature Embryos | Number of Positive T0 Plants | Transformation Ratio (%) |
|---|---|---|---|---|
| Zheng58 | pHB-CB-NW | 238 | 7 | 2.94 |
| pCA-GFP | ||||
| Zheng58 | pHB-CB-AW | 695 | 1 | 0.14 |
| pCA-GFP | ||||
| Mo17 | pHB-CB-AW | 301 | 2 | 0.66 |
| pCA-GFP | ||||
| Mo17 | pHB-PB-AW-GFP | 509 | 8 | 1.57 |
| Chang7-2 | data pHB-CB-AW-GFP | 692 | 4 | 0.58 |
The transformation ratio was calculated based on the number of transgenic positive T0 plants [(number of T0 plants ÷ number of embryos) × 100].
Table 2.
Segregation ratio of T2 transgenic Zheng58 seeds.
| Maize Inbred Line | Event | Number of Detected T2 Seeds | Number of Coexistent Seeds | Number of Separated Seeds | Segregation Ratio (%) |
|---|---|---|---|---|---|
| Zheng58 | 1# | 59 | 55 | 4 | 6.78% |
| 2# | 107 | 78 | 4 | 2.80% | |
| 3# | 68 | 38 | 3 | 4.41% | |
| 4# | 148 | 118 | 4 | 2.03% |
T2 seeds were progenies of transgenic Zheng58 of lines 1#, 2#, 3# and 4#. The number of coexisting seeds was the number of T2 transgenic Zheng58 seeds, including ZmCTA1pro::BBM-NOSpro::WUS and 35Spro::GFP. The number of separated seeds was the number of T2 transgenic Zheng58 seeds containing only 35Spro::GFP. The segregation ratio was calculated based on the number of separated seeds. [(Number of separated seeds ÷ number of T2 seeds) × 100].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gu, Y.; Chen, X.; Song, R.; Qi, W. Establishment of a Bivector Genetic Transformation System in Recalcitrant Maize Inbred Lines. Agriculture 2021, 11, 663. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070663
AMA Style
Gu Y, Chen X, Song R, Qi W. Establishment of a Bivector Genetic Transformation System in Recalcitrant Maize Inbred Lines. Agriculture. 2021; 11(7):663. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070663
Chicago/Turabian StyleGu, Yajing, Xuan Chen, Rentao Song, and Weiwei Qi. 2021. "Establishment of a Bivector Genetic Transformation System in Recalcitrant Maize Inbred Lines" Agriculture 11, no. 7: 663. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070663
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.