
Increased Activity of 5-Enolpyruvylshikimate-3-phosphate Synthase (EPSPS) Enzyme Describe the Natural Tolerance of Vulpia myuros to Glyphosate in Comparison with Apera spica-venti

 , and
, and
Abstract
:1. Introduction
2. Material and Method
2.1. Seed Source
2.2. Dose-Response and Spray Retention Study
2.2.1. Dose-Response Study
2.2.2. Spray Retention Assay
2.3. Shikimic Acid Accumulation Assays
2.3.1. Whole Plant Shikimic Acid Accumulation
2.3.2. Accumulation of Shikimic Acid in Excised Leaves
2.4. EPSPS Enzyme Sensitivity
2.5. Statistical Analysis
3. Results
3.1. Dose-Response and Spray Retention Study
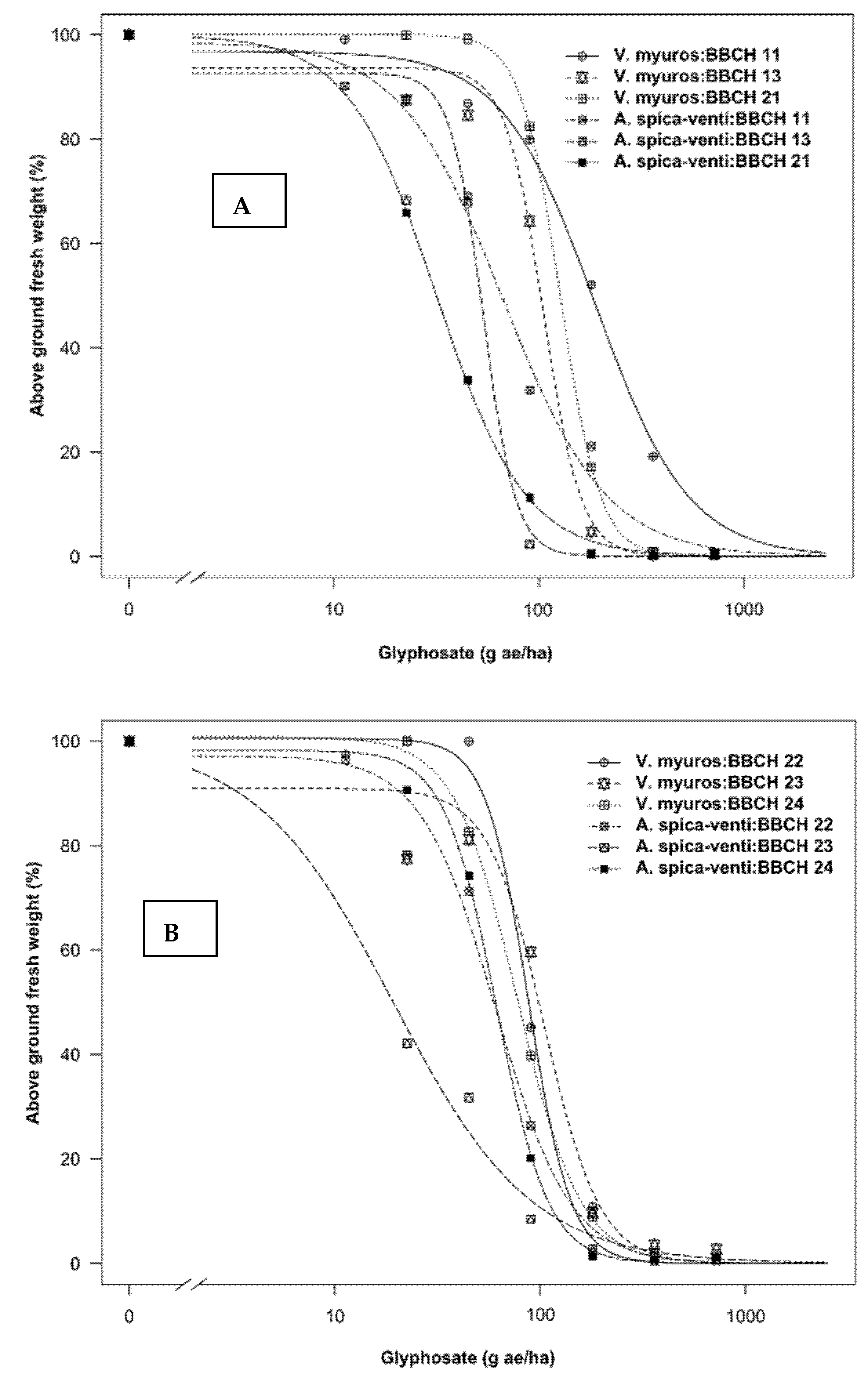
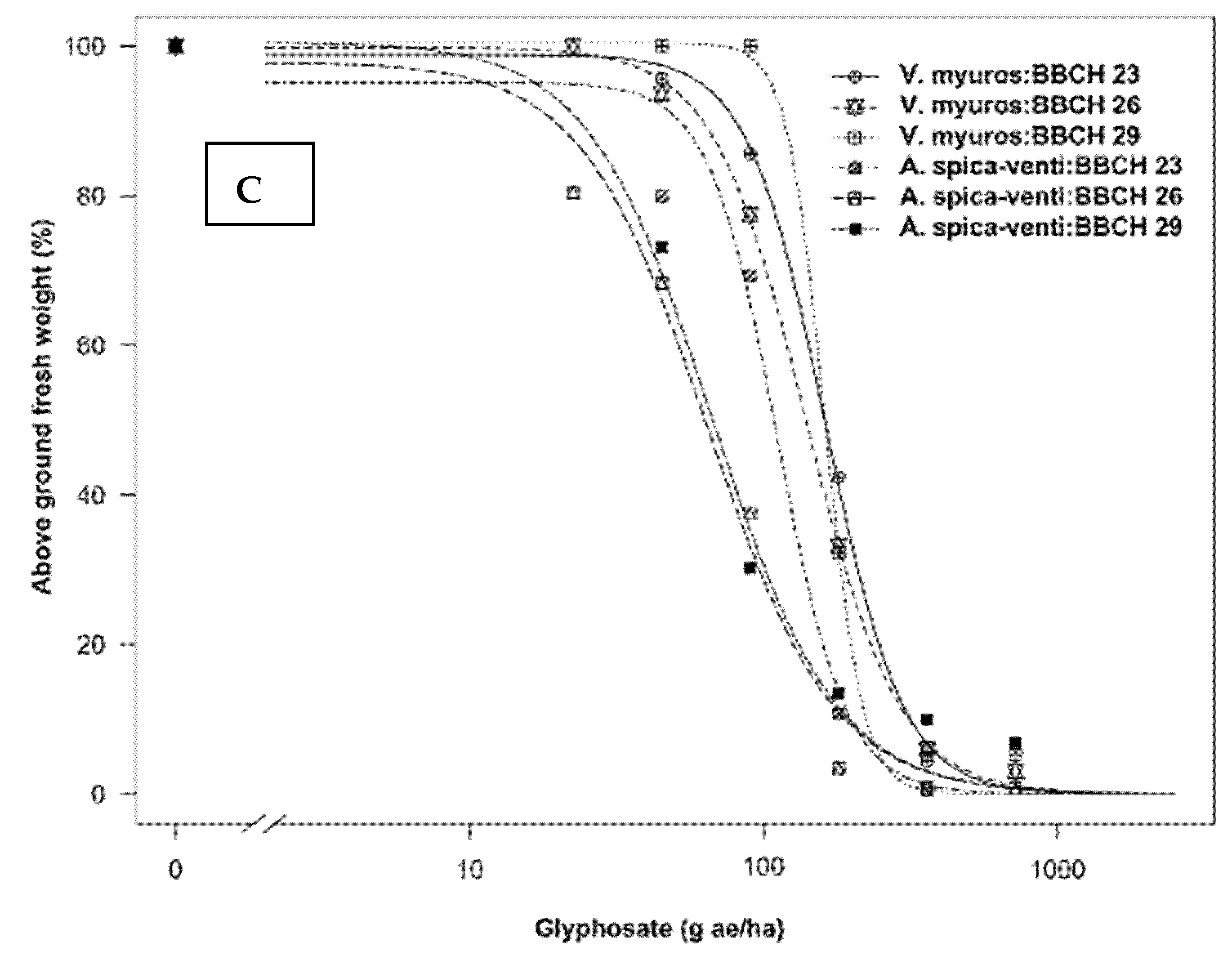
3.1.1. Dose-Response Assay
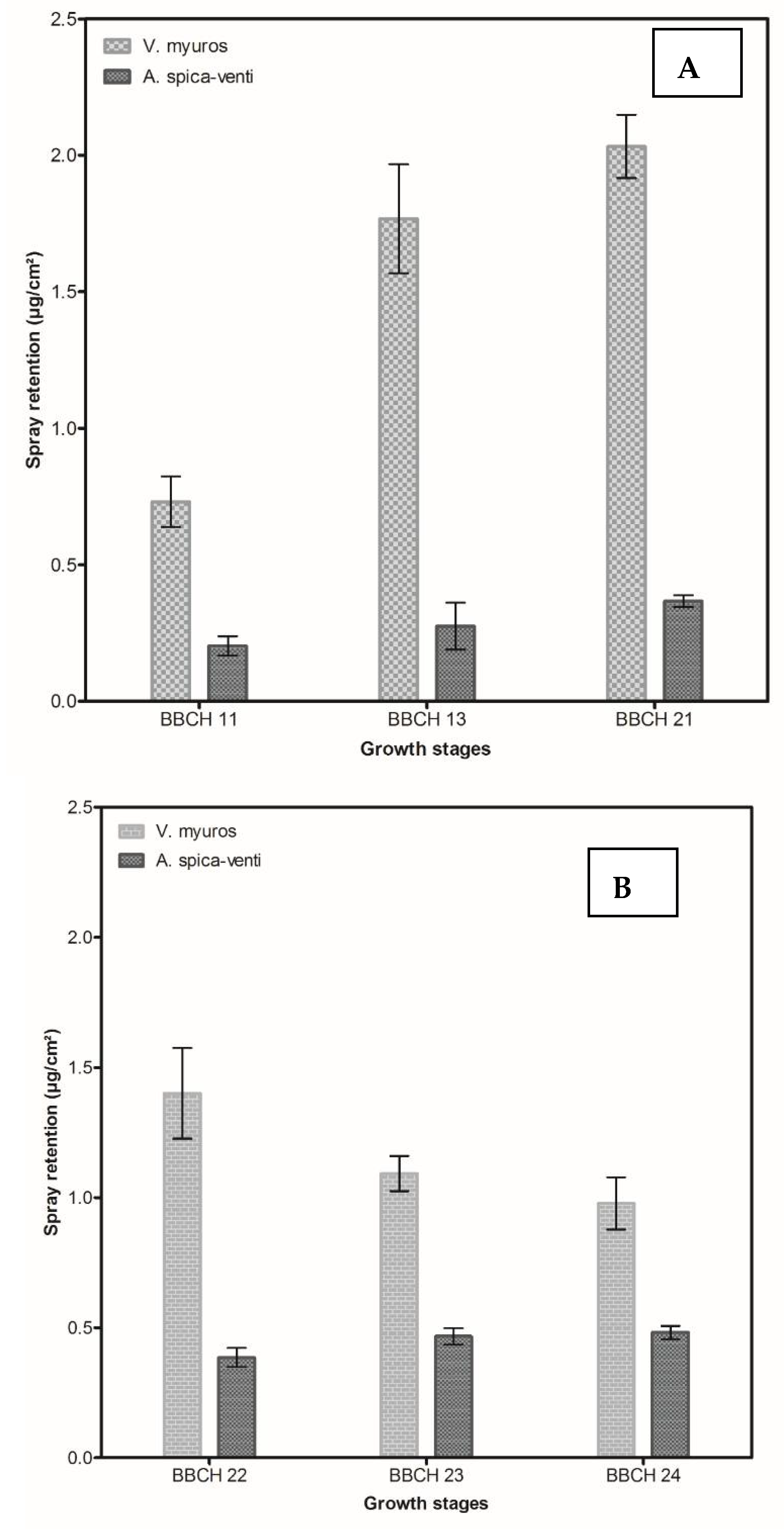
3.1.2. Spray Retention Assay
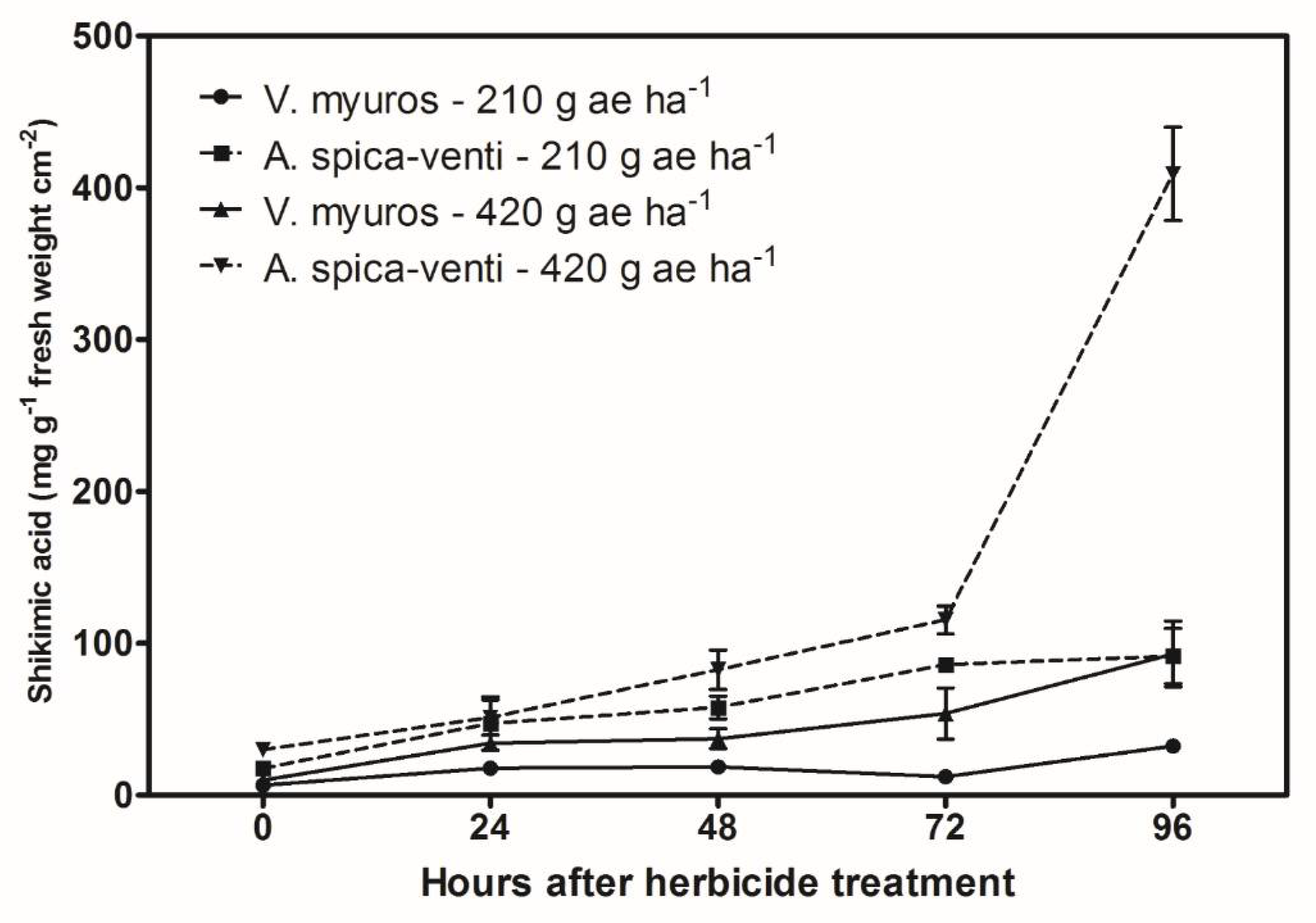
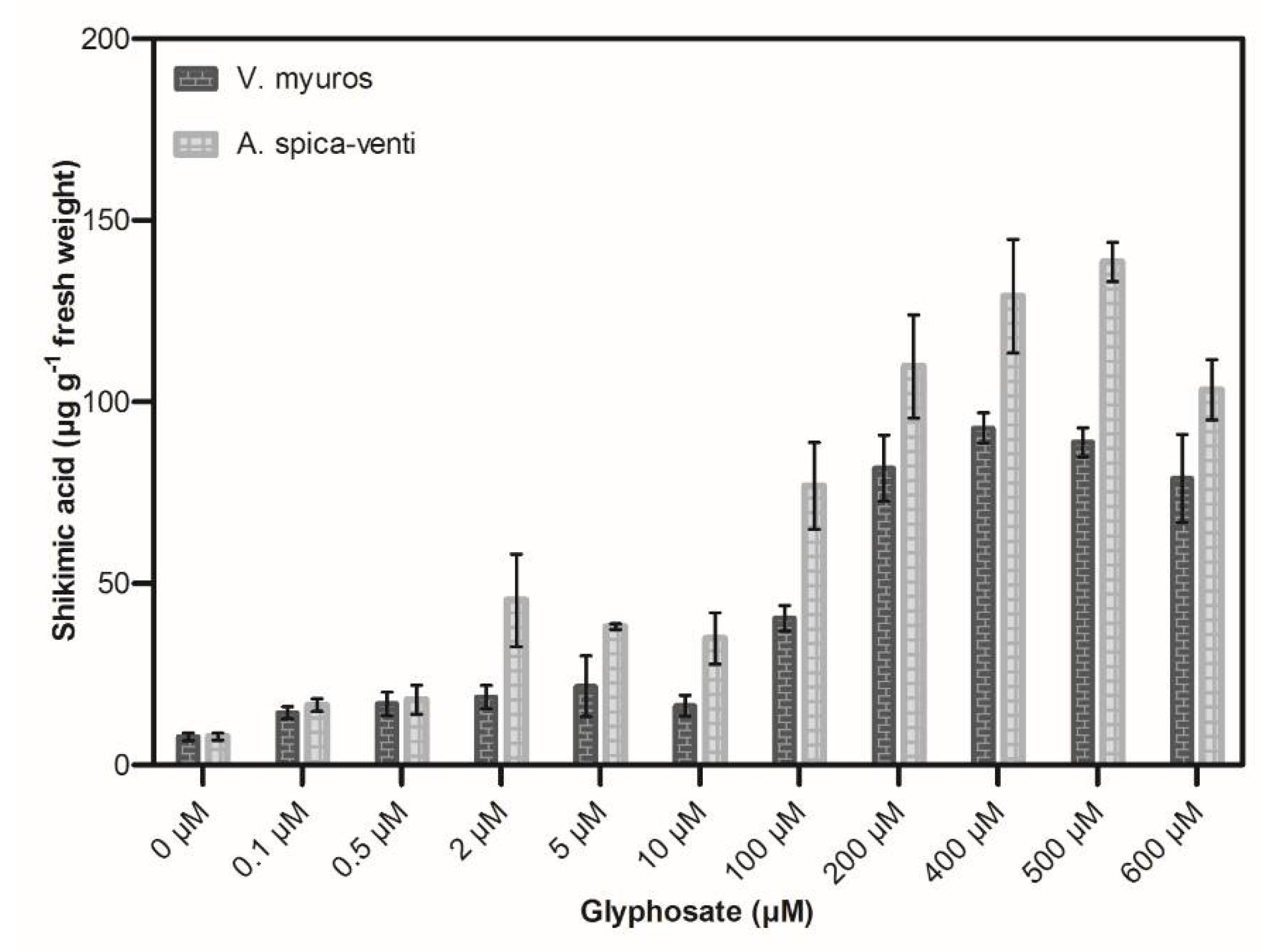
3.2. Shikimic Acid Accumulation Assays
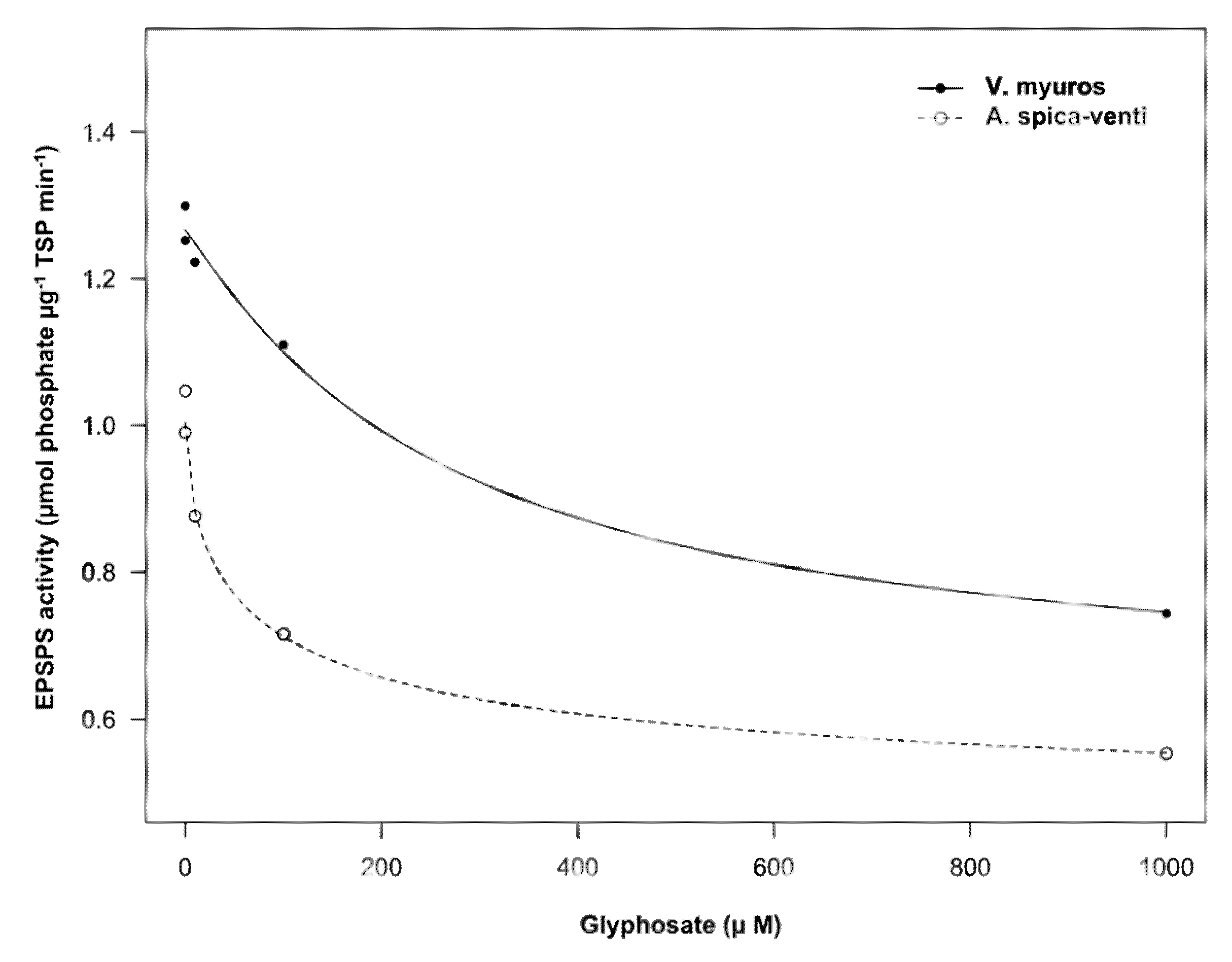
3.3. EPSPS Enzyme Sensitivity
4. Discussion
4.1. Dose-Response and Spray Retention Study
4.1.1. Dose-Response Study
4.1.2. Spray Retention Study
4.2. Shikimic Acid Accumulation Assays
4.3. EPSPS Enzyme Sensitivity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akhter, M.J. Biology and Management of Vulpia Myuros in Arable Farming. Ph.D. Thesis, Aarhus University, Aarhus, Denmark, 16 December 2020. [Google Scholar]
- Mathiassen, S.K.; Henriksen, K.E. Experience on Vulpia myuros in Denmark-biology, seed survival, herbicide efficacy and selectivity. In Proceedings of the Tagungsunterlagen, 51. Fachtagung des DLG-Ausschusses “Gräser, Klee und Zwischenfrüchte”, Bonn, Germany, 5 November 2010; Susanne, S., Ed.; Fachzeentrum Land-und Ernahrungswirtschaft Eschboorner Landstrabe 122: Frankfurt am Main, Germany, 2010. [Google Scholar]
- Hull, R.; Mathiassen, S.K.; Moss, S.R. Herbicidal control of Vulpia myuros (Rat’s-tail fescue) in glasshouse screening tests. Asp. Appl. Biol. 2011, 106, 75–81. [Google Scholar]
- Akhter, M.J.; Jensen, P.K.; Mathiassen, S.K.; Melander, B.; Kudsk, P. Biology and Management of Vulpia myuros—An Emerging Weed Problem in No-Till Cropping Systems in Europe. Plants 2020, 9, 715. [Google Scholar] [CrossRef] [PubMed]
- Javaid Akhter, M.; Melander, B.; Mathiassen, S.K.; Labouriau, R.; Vendelbo Nielsen, S.; Kudsk, P. Growth and Phenology of Vulpia Myuros in Comparison with Apera spica-venti, Alopecurus Myosuroides and Lolium Multiflorum in Monoculture and in Winter Wheat. Plants 2020, 9, 1495. [Google Scholar] [CrossRef] [PubMed]
- Ball, D.A.; Frost, S.M.; Fandrich, L.; Tarasoff, C.; Mallory-Smith, C. Biological Attributes of Rattail Fescue (Vulpia myuros). Weed Sci. 2008, 56, 26–31. [Google Scholar] [CrossRef]
- Akhter, M.J.; Kudsk, P.; Mathiassen, S.K.; Melander, B. Rattail fescue (Vulpia myuros) interference and seed production as affected by sowing time and crop density in winter wheat. Weed Sci. 2021, 69, 52–61. [Google Scholar] [CrossRef]
- Scherner, A.; Melander, B.; Kudsk, P. Vertical distribution and composition of weed seeds within the plough layer after eleven years of contrasting crop rotation and tillage schemes. Soil Tillage Res. 2016, 161, 135–142. [Google Scholar] [CrossRef]
- Kudsk, P.; Mathiassen, S.K. Pesticide regulation in the European Union and the glyphosate controversy. Weed Sci. 2020, 68, 214–222. [Google Scholar] [CrossRef]
- Bowran, D.; Wallace, A. Chemical weed management of Vulpia. Plant Prot. Q. 1996, 11, 211–212. [Google Scholar]
- Ball, D.A.; Hulting, A.G.; Lyon, D.J. Rattail Fescue: Biology and Management in Pacific Northwest Wheat Cropping Systems. 2018. Available online: https://pubs.extension.wsu.edu/rattail-fescue-biology-and-management-in-pacific-northwest-wheat-cropping-systems (accessed on 4 June 2020).
- Steinrücken, H.C.; Amrhein, N. The herbicide glyphosate is a potent inhibitor of 5-enolpyruvylshikimic acid-3-phosphate synthase. Biochem. Biophys. Res. Commun. 1980, 94, 1207–1212. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Corniani, N.; Silva, F.M.L.; Watson, S.B.; Howell, J.L.; Shaner, D.L. Biochemical markers and enzyme assays for herbicide mode of action and resistance studies. Weed Sci. 2015, 63, 23–63. [Google Scholar] [CrossRef] [Green Version]
- Sammons, R.D.; Gaines, T.A. Glyphosate resistance: State of knowledge. Pest Manag. Sci. 2014, 70, 1367–1377. [Google Scholar] [CrossRef] [Green Version]
- Gaines, T.A.; Zhang, W.; Wang, D.; Bukun, B.; Chisholm, S.T.; Shaner, D.L.; Nissen, S.J.; Patzoldt, W.L.; Tranel, P.J.; Culpepper, A.S. Gene amplification confers glyphosate resistance in Amaranthus palmeri. Proc. Natl. Acad. Sci. USA 2010, 107, 1029–1034. [Google Scholar] [CrossRef] [Green Version]
- Moretti, M.L.; Van Horn, C.R.; Robertson, R.; Segobye, K.; Weller, S.C.; Young, B.G.; Johnson, W.G.; Douglas Sammons, R.; Wang, D.; Ge, X. Glyphosate resistance in Ambrosia trifida: Part 2. Rapid response physiology and non-target-site resistance. Pest Manag. Sci. 2018, 74, 1079–1088. [Google Scholar] [CrossRef]
- Yu, Q.; Shane Friesen, L.J.; Zhang, X.Q.; Powles, S.B. Tolerance to acetolactate synthase and acetyl-coenzyme A carboxylase inhibiting herbicides in Vulpia bromoides is conferred by two co-existing resistance mechanisms. Pestic. Biochem. Physiol. 2004, 78, 21–30. [Google Scholar] [CrossRef]
- Schroeder, J. Minutes of the WSSA Board of Directors Meeting Chicago, IL February 12, 1998. Weed Sci. 1998, 46, 628–630. [Google Scholar] [CrossRef]
- Crease, G.J.; Hall, F.R.; Thacker, J.R.M. Reflection of agricultural sprays from leaf surfaces. J. Environ. Sci. Health B 1991, 26, 383–407. [Google Scholar] [CrossRef]
- Ball, D.A.; Frost, S.M.; Bennett, L.H.; Thill, D.C.; Rauch, T.; Jemmett, E.; Mallory-Smith, C.; Cole, C.; Yenish, J.P.; Rood, R. Control of rattail fescue (Vulpia myuros) in winter wheat. Weed Technol. 2007, 21, 583–590. [Google Scholar] [CrossRef]
- Babineau, M.; Mathiassen, S.K.; Kristensen, M.; Kudsk, P. Fitness of ALS-Inhibitors Herbicide Resistant Population of Loose Silky Bentgrass (Apera spica-venti). Front. Plant Sci. 2017, 8, 1660. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Shaner, D.L. Rapid determination of glyphosate injury to plants and identification of glyphosate-resistant plants. Weed Technol. 1998, 12, 527–530. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e146021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritz, C.; Streibig, J.C. Bioassay analysis using R. J. Stat. Softw. 2005, 12. [Google Scholar] [CrossRef] [Green Version]
- Jemmett, E.D.; Thill, D.C.; Rauch, T.A.; Ball, D.A.; Frost, S.M.; Bennett, L.H.; Yenish, J.P.; Rood, R.J. Rattail fescue (Vulpia myuros) control in chemical-fallow cropping systems. Weed Technol. 2008, 22, 435–441. [Google Scholar] [CrossRef]
- Michitte, P.; De Prado, R.; Espinoza, N.; Ruiz-Santaella, J.P.; Gauvrit, C. Mechanisms of resistance to glyphosate in a ryegrass (Lolium multiflorum) biotype from Chile. Weed Sci. 2007, 55, 435–440. [Google Scholar] [CrossRef]
- Pérez-López, M.; González-Torralva, F.; Cruz-Hipólito, H.; Santos, F.; Domínguez-Valenzuela, J.A.; De Prado, R. Characterization of glyphosate-resistant tropical sprangletop (Leptochloa virgata) and its alternative chemical control in Persian lime orchards in Mexico. Weed Sci. 2014, 62, 441–450. [Google Scholar] [CrossRef]
- Wallace, A. The biology of Australian weeds. 30. Vulpia bromoides ((L.) SF Gray) and V. myuros ((L.) CC Gmelin). Plant Prot. Q. 1997, 12, 18–28. [Google Scholar]
- Warwick, S.I.; Black, L.D.; Zilkey, B.F. Biology of canadian weeds: 72. Apera spica-venti. Can. J. Plant Sci. 1985, 65, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Holloway, P.J. Physicochemical factors influencing the adjuvant-enhanced spray deposition and coverage of foliage-applied agrochemicals. In Interactions between Adjuvants, Agrochemicals and Target Organisms; Springer: Berlin/Heidelberg, Germany, 1994; pp. 83–106. [Google Scholar]
- Mathiassen, S.K. XI Effects of new adjuvants, N32 and pH of the spray solution on herbicide efficacy. Appl. Crop Protec. 2016, 94, 119–123. [Google Scholar]
- Powell, E.S.; Orson, J.H.; Miller, P.C.H.; Kudsk, P.; Mathiassen, S.K. Defining the size of target for air induction nozzles. Defining the size of target for air induction nozzles. Asp. Appl. Biol. 2002, 66, 65–72. [Google Scholar]
- Koo, S.-J.; Kim, J.-S.; Lee, J.-H. Foliar retention of the herbicide pyribenzoxim (1% EC), and its effects on herbicidal activity and rice phytotoxicity. Korean J. Weed Sci. 1998, 18, 304–313. [Google Scholar]
- Salas, R.A.; Dayan, F.E.; Pan, Z.; Watson, S.B.; Dickson, J.W.; Scott, R.C.; Burgos, N.R. EPSPS gene amplification in glyphosate-resistant Italian ryegrass (Lolium perenne ssp. multiflorum) from Arkansas. Pest Manag. Sci. 2012, 68, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.A. The shikimate/arogenate pathway: Link between carbohydrate metabolism and secondary metabolism. Physiol. Plant. 1986, 66, 164–168. [Google Scholar] [CrossRef]
- Zimdahl, R.L. Weed-Crop Competition: A Review; International Plant Protection Center: Corvallis, OR, USA, 1980. [Google Scholar]
- Perez-Jones, A.; Park, K.W.; Colquhoun, J.; Mallory-Smith, C.; Shaner, D. Identification of glyphosate-resistant Italian ryegrass (Lolium multiflorum) in Oregon. Weed Sci. 2005, 53, 775–779. [Google Scholar] [CrossRef]
- Fernández-Moreno, P.T.; Alcántara-de la Cruz, R.; Smeda, R.J.; De Prado, R. Differential resistance mechanisms to glyphosate result in fitness cost for Lolium perenne and L. multiflorum. Front. Plant Sci. 2017, 8, 1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alarcón-Reverte, R.; García, A.; Watson, S.B.; Abdallah, I.; Sabaté, S.; Hernández, M.J.; Dayan, F.E.; Fischer, A.J. Concerted action of target-site mutations and high EPSPS activity in glyphosate-resistant junglerice (Echinochloa colona) from California. Pest Manag. Sci. 2015, 71, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Kong, W.; Johnston, R.M.; Nabukalu, P.; Wu, G.; Poehlman, W.L.; Goff, V.H.; Isaacs, K.; Lee, T.-H.; Guo, H. The evolution of an invasive plant, Sorghum halepense L. (‘Johnsongrass’). Front. Genet. 2020, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Soltis, D.E.; Buggs, R.J.A.; Doyle, J.J.; Soltis, P.S. What we still don’t know about polyploidy. Taxon 2010, 59, 1387–1403. [Google Scholar] [CrossRef]
- Bunnell, B.T.; Baker, R.D.; McCarty, L.B.; Hall, D.W.; Colvin, D.L. Differential Response of Five Bahiagrass (Paspalum notatum) Cultivars to Metsulfuron1. Weed Technol. 2003, 17, 550–553. [Google Scholar] [CrossRef]
- Akhter, M.J.; Abdurruhman, A.M.; Mathiassen, S.K.; Kudsk, P. Low Effectiveness of Prosulfocarb and Mesosulfuron-methyl+ Iodosulfuron-methyl against Vulpia myuros. Plants 2020, 10, 1186. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Experiment | Treatment (Glyphosate Doses; Growth Stage (BBCH)) | Environment |
|---|---|---|---|
| Dose-response | Experiment 1 | Glyphosate rate ranging from 11.3 to 720 g ha−1; BBCH 11, BBCH 13, BBCH 21 | Unheated glasshouse |
| Experiment 2 | Glyphosate rate ranging from 11.3 to 720 g ha−1; BBCH 22, BBCH 23, BBCH 24 | Unheated glasshouse | |
| Experiment 3 | Glyphosate rate ranging from 11.3 to 720 g ha−1; BBCH 22, BBCH 26, BBCH 29 | Outdoor under natural conditions | |
| Spray retention | Experiment 1 | Glyphosate rate at 90 g ha−1 in mixture with fluorescent dye at a concentration of 200 g ha−1; BBCH 11, BBCH 13, BBCH 21 | Unheated glasshouse, Laboratory |
| Experiment 2 | Glyphosate rate at 90 g ha−1 in mixture with fluorescent dye at a concentration of 200 g ha−1; BBCH 11, BBCH 13, BBCH 21 | Unheated glasshouse, Laboratory | |
| Experiment 3 | Glyphosate rate at 90 g ha−1 in mixture with fluorescent dye at a concentration of 200 g ha−1; BBCH 22, BBCH 23, BBCH 24 | Outdoor under natural conditions, Laboratory | |
| Whole plant shikimic acid accumulation | Glyphosate rate at 210 g ha−1 and 420 g ha−1; BBCH 23 | Laboratory | |
| Accumulation of shikimic acid in excised leaves | Glyphosate rate ranging from 0 to 600 μM; BBCH 23 to BBCH 25 | Laboratory | |
| EPSPS enzyme sensitivity | Glyphosate rate ranging from 0 to 1000 μM; BBCH 23 | Laboratory |
| Growth Stage | ||||
|---|---|---|---|---|
| Species | BBCH 11 | BBCH 13 | BBCH 21 | |
| Experiment 1 | V. myuros | 188 (25.2) | 105 (11.5) | 127 (10.3) |
| A. spica-venti | 67 (10.0) | 53 (6.2) | 32 (3.9) | |
| Tolerance indices | TI (p value) | 2.8 (p = 0.002) | 2.0 (p = 0.003) | 4.0 (p < 0.001) |
| Growth stages | Species | BBCH 22 | BBCH 23 | BBCH 24 |
| Experiment 2 | V. myuros | 88 (4.6) | 105 (12.8) | 78 (4.7) |
| A. spica-venti | 61 (5.6) | 19 (5.8) | 61 (3.5) | |
| Tolerance indices | TI (p value) | 1.4 (p = 0.006) | 5.4 (p = 0.016) | 1.3 (p = 0.015) |
| Growth stages | Species | BBCH 23 | BBCH 26 | BBCH 29 |
| Experiment 3 | V. myuros | 163 (13.1) | 140 (10.4) | 165 (38.6) |
| 1 | 2 | 3 | ||
| A. spica-venti | 111 (8.9) | 65 (6.5) | 61 (2.3) | |
| Tolerance indices b | TI (p value) | 1.4 (p = 0.0099) | 2.16 (p < 0.001) | 2.72 (p = 0.011) |
| Regression Parameter Estimates a | ||||
|---|---|---|---|---|
| Species | b | c (µmol phosphate µg−1 TSP min−1) | d (µmol phosphate µg−1 TSP min−1) | I50 (μM) |
| Vulpia myuros | 1.1 (0.51) | 0.6 (0.18) | 1.3 (0.01) | 264.0 (191) |
| Apera spica-venti | 0.6 (0.21) | 0.4 (0.11) | 1.0 (0.02) | 81.1 (75.8) |
| Significance level b | p = 0.3957 | p = 0.444 | p < 0.001 | p = 0.3834 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhter, M.J.; Mathiassen, S.K.; Bekalu, Z.E.; Brinch-Pedersen, H.; Kudsk, P. Increased Activity of 5-Enolpyruvylshikimate-3-phosphate Synthase (EPSPS) Enzyme Describe the Natural Tolerance of Vulpia myuros to Glyphosate in Comparison with Apera spica-venti. Agriculture 2021, 11, 725. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080725
Akhter MJ, Mathiassen SK, Bekalu ZE, Brinch-Pedersen H, Kudsk P. Increased Activity of 5-Enolpyruvylshikimate-3-phosphate Synthase (EPSPS) Enzyme Describe the Natural Tolerance of Vulpia myuros to Glyphosate in Comparison with Apera spica-venti. Agriculture. 2021; 11(8):725. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080725
Chicago/Turabian StyleAkhter, Muhammad Javaid, Solvejg Kopp Mathiassen, Zelalem Eshetu Bekalu, Henrik Brinch-Pedersen, and Per Kudsk. 2021. "Increased Activity of 5-Enolpyruvylshikimate-3-phosphate Synthase (EPSPS) Enzyme Describe the Natural Tolerance of Vulpia myuros to Glyphosate in Comparison with Apera spica-venti" Agriculture 11, no. 8: 725. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080725