
Response of Cover Crops to Phytopythium vexans, Phytophthora nicotianae, and Rhizoctonia solani, Major Soilborne Pathogens of Woody Ornamentals


Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Screening of Cover Crop Germination
2.2. Pathogens: Culture and Inoculum Preparation
2.3. Greenhouse Bioassay for Pathogenicity Test
2.4. Assessment of Cover Crop Health
2.5. Data Analysis
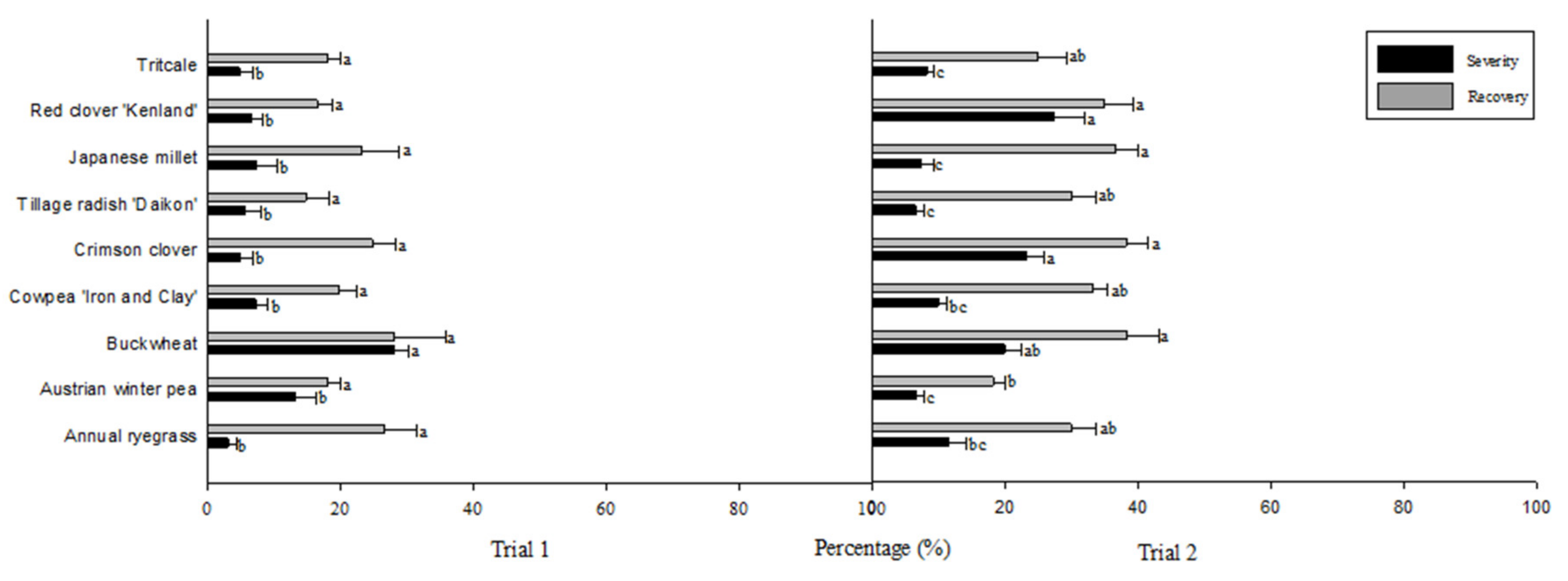
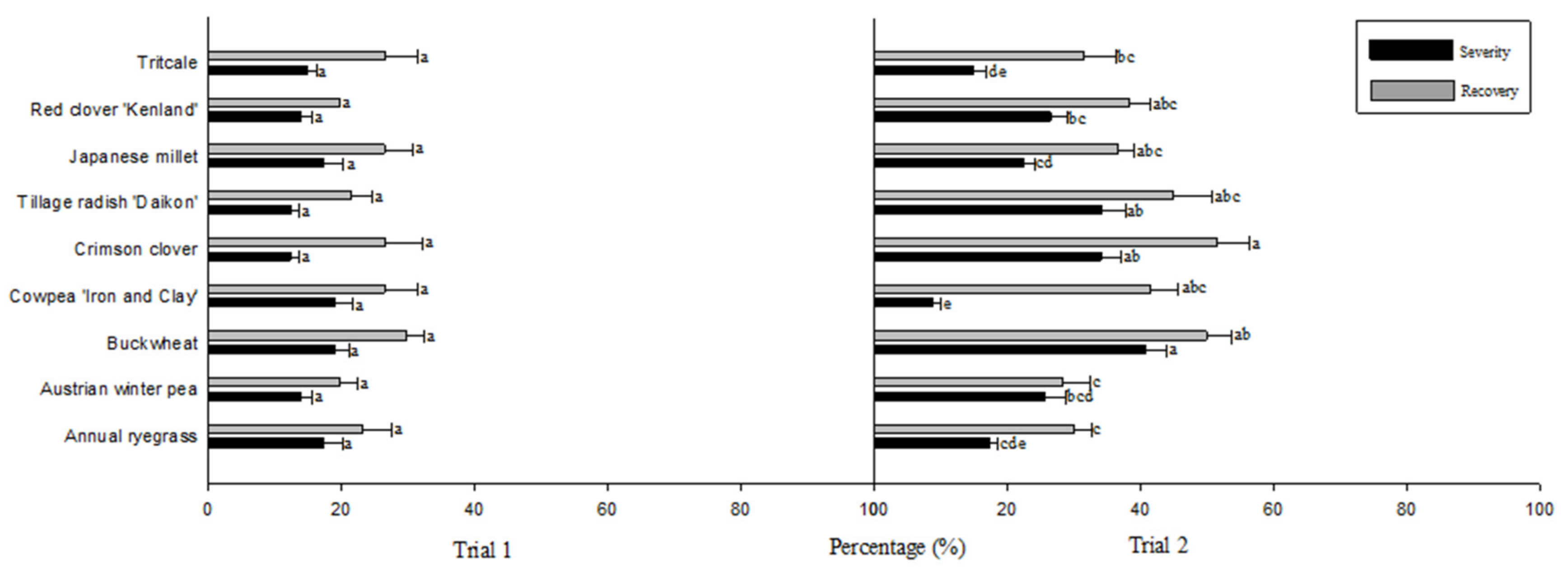
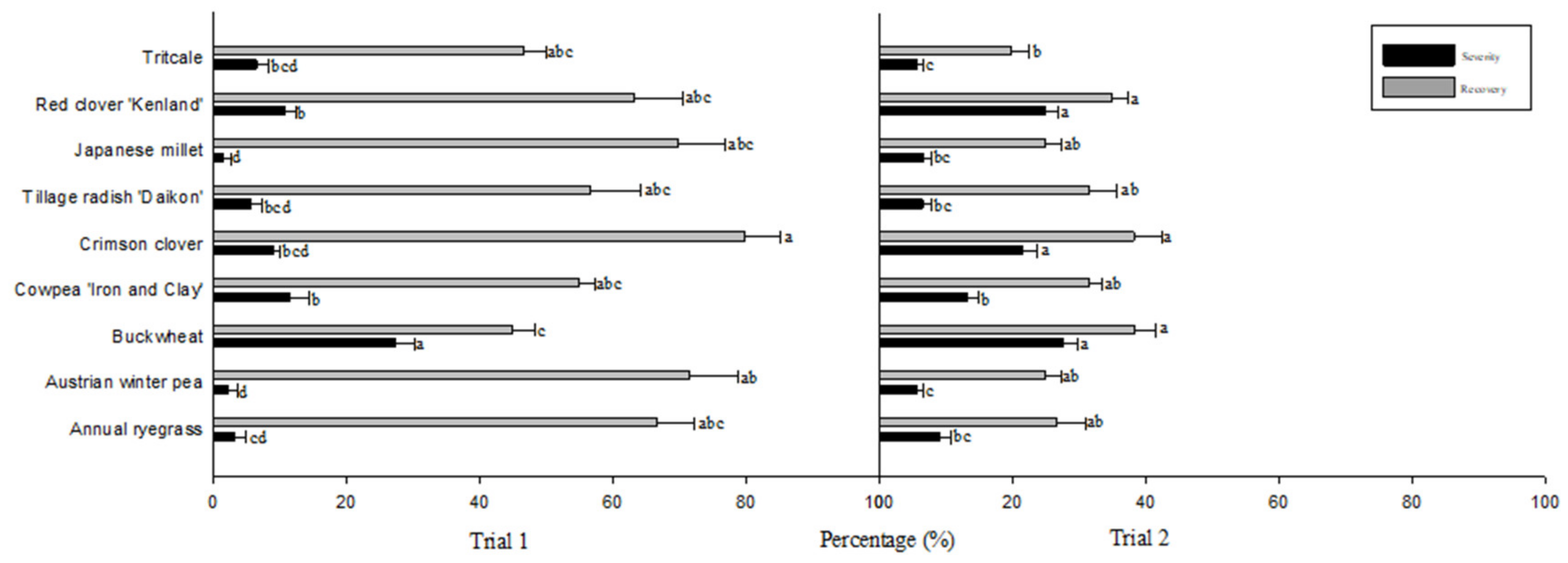
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United States Department of Agriculture. Census of Agriculture. 2017. Available online: https://www.nass.usda.gov/Publications/AgCensus/2017/index.php#full_report (accessed on 8 September 2020).
- Mokhtar, M.M.; El-Mougy, N.S. Biocompost application for controlling soilborne plant pathogens—A review. Int. J. Eng. Innov. Technol. 2014, 4, 61–68. [Google Scholar]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soilborne diseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Cacciola, S.O.; di San Lio, G.M. Management of citrus diseases caused by Phytophthora spp. In Integrated Management of Diseases Caused by Fungi, Phytoplasma and Bacteria; Ciancio, A., Mukerji, K.G., Eds.; Springer Science Business Media: New York, NY, USA, 2008; pp. 61–84. [Google Scholar]
- Baysal-Gurel, F.; Liyanapathiranage, P.; Mullican, J. Biofumigation: Opportunities and challenges for control of soilborne diseases in nursery production. Plant Health Prog. 2018, 19, 332–337. [Google Scholar] [CrossRef]
- Hu, J.H.; Hong, C.X.; Stromberg, E.L.; Moorman, G.W. Mefenoxam sensitivity and fitness analysis of Phytophthora nicotianae isolates from nurseries in Virginia, USA. Plant Pathol. 2008, 57, 728–736. [Google Scholar] [CrossRef]
- Moralejo, E.; Perez-Sierra, A.M.; Alvarez, L.A.; Belbahri, L.; Lefort, F.; Descals, E. Multiple alien Phytophthora taxa discovered on diseased ornamental plants in Spain. Plant Pathol. 2009, 58, 100–110. [Google Scholar] [CrossRef]
- Pane, A.; Martini, P.; Chimento, A.; Rapetti, S.; Savona, S.; Grasso, F.M.; Cacciola, S.O. Phytophthora species on ornamental plants in Italy. J. Plant Pathol. 2005, S87, 301. [Google Scholar]
- Gallup, C.A.; Sullivan, M.; Shew, H.D. Black Shank of tobacco. Plant Health Instr. 2006. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Liyanapathiranage, P.; Panth, M.; Avin, F.A.; Simmons, T. First report of Phytopythium vexans causing root and crown rot on Flowering Cherry in Tennessee. Plant Dis. 2020, 105, 232. [Google Scholar] [CrossRef] [PubMed]
- Benfradj, N.; Migliorini, D.; Luchi, N.; Santini, A.; Boughalleb-M’Hamdi, N. Occurrence of Pythium and Phytopythium species isolated from citrus trees infected with gummosis disease in Tunisia. Arch. Phytopathol. Plant Prot. 2017, 50, 286–302. [Google Scholar] [CrossRef]
- Panth, M.; Baysal-Gurel, F.; Avin, F.A.; Simmons, T. Identification, chemical and biological management of Phytopythium vexans, the causal agent of Phytopythium root and crown rot of woody ornamentals. Plant Dis. 2020, 105, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Polat, Z.; Awan, Q.N.; Hussain, M.; Akgul, D.S. First report of Phytopythium vexans causing root and collar rot of kiwifruit in Turkey. Plant Dis. 2017, 101, 1058. [Google Scholar] [CrossRef]
- Rodriguez-Padron, C.; Siverio, F.; Perez-Sierra, A.; Rodriguez, A. Isolation and pathogenicity of Phytophthora species and Phytopythium vexans recovered from avocado orchards in the Canary Islands, including Phytophthora niederhauserii as a new pathogen of avocado. Phytopathol. Mediter. 2018, 57, 89–106. [Google Scholar] [CrossRef]
- Farr, D.F.; Bills, G.F.; Chamuris, G.P.; Rossman, A.Y. Fungi on Plant and Plant Products in the United States; American Phytopathological Society: St. Paul, MN, USA, 1995. [Google Scholar]
- Hyakumachi, M.; Priyatmojo, A.; Kubota, M.; Fukui, H. New anastomosis groups, AG-T and AG-U, of binucleate Rhizoctonia spp. causing root and stem rot of cut-flower and miniature roses. Phytopathology 2005, 95, 784–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinehart, T.A.; Copes, W.E.; Toda, T.; Cubeta, M.A. Genetic characterization of binucleate Rhizoctonia species causing web blight on azalea in Mississippi and Alabama. Plant Dis. 2007, 91, 616–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzola, M.; Freilich, S. Prospects for biological soilborne disease control: Application of indigenous versus synthetic microbiomes. Phytopathology 2017, 107, 256–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawadi, S.; Baysal-Gurel, F.; Addesso, K.M.; Oliver, J.B.; Simmons, T. Impact of cover crop usage on soilborne diseases in field nursery production. Agronomy 2019, 9, 753. [Google Scholar] [CrossRef] [Green Version]
- Baysal-Gurel, F.; Liyanapathiranage, P. Pathogenecity of Rhizoctonia solani and Phytophthora nicotianae to Brassicaceae cover crops. Arch. Phytopathol. Plant Prot. 2019, 52, 288–302. [Google Scholar] [CrossRef]
- Panth, M.; Baysal-Gurel, F.; Simmons, T.; Addesso, K.M.; Witcher, A. Impact of winter cover crop usage in soilborne disease suppressiveness in woody ornamental production system. Agronomy 2020, 10, 995. [Google Scholar] [CrossRef]
- Finney, D.M.; Buyer, J.S.; Kaye, J.P. Living cover crops have immediate impacts on soil microbial community structure and function. J. Soil Water Conserv. 2017, 72, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Anosheh, H.P.; Sadeghi, H.; Emam, Y. Chemical priming with urea and KNO3 enhances maize hybrids (Zea mays L.) seed viability under abiotic stress. J. Crop Sci. Biotechnol. 2011, 14, 289–295. [Google Scholar] [CrossRef]
- Holmes, K.A.; Benson, D.M. Evaluation of Phytophthora parasitica var. nicotianae for biocontrol of Phytophthora parasitica on Catharanthus roseus. Plant Dis. 1994, 78, 193–199. [Google Scholar]
- Baysal-Gurel, F.; Simmons, T.; Liyanapathiranage, P.; Kabir, M.N. Evaluation of Biorational Products and Fungicides for the Control of Rhizoctonia Root Rot of Viburnum. Plant Disease Management Report No. 11: OT003. Online publication. Am. Phytopath. Soc. 2017. [Google Scholar]
- Jeffers, S.N.; Martin, S.B. Comparison of two media selective for Phytophthora and Pythium species. Plant Dis. 1986, 70, 1038–1043. [Google Scholar] [CrossRef]
- Ferguson, A.J.; Jeffers, S.N. Detecting multiple species of Phytophthora in container mixes from ornamental crop nurseries. Plant Dis. 1999, 83, 1129–1136. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, W.A.; Shew, H.D.; Melton, T.A. A Semi-Selective Medium to Isolate Rhizoctonia solani from Soil and Tissue; Plant Path. Extension, NC State Extension: Raleigh, NC, USA, 2001; pp. 1–2. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. PCR Protocols: A Guide to Methods and Applications; Gelfand, D.H., Innis, M.A., Shinsky, J.J., White, T.J., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Sarrantonio, M.; Gallandt, E. The Role of Cover Crops in North American Cropping Systems. J. Crop Prod. 2003, 8, 53–74. [Google Scholar] [CrossRef]
- Lazzeri, L.; Baruzzi, G.; Malaguti, L.; Antoniacci, L. Replacing methyl bromide in annual strawberry production with glucosinolate-containing green manure crops. Pest. Manag. Sci. 2003, 59, 983–990. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using brassica green manures. Crop Prot. 2006, 26, 1067–1077. [Google Scholar] [CrossRef]
- Bakker, M.G.; Acharya, J.; Moorman, T.B.; Robertson, A.E.; Kaspar, T.C. The potential for cereal rye cover crops to host corn seedling pathogens. Phytopathology 2016, 106, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A.; Kaspar, T.; Leandro, L.; Mueller, D.; Acharya, J. Disease risks associated with cover crops and soybean production. Proc. Integr. Crop Manag. Conf. 2017, 21, 125–128. Available online: https://lib.dr.iastate.edu/icm/2017/proceedings/21 (accessed on 1 November 2020).
- Stivers-Young, L.J.; Tucker, F.A. Cover-cropping practices of vegetable producers in western New York. HortTechnology 1999, 9, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Mallory, E.B.; Posner, J.L.; Baldock, J.O. Performance, economics, and adoption of cover crops in Wisconsin cash grain rotations: On-farm trials. Am. J. Altern. Agric. 1998, 13, 2–11. [Google Scholar] [CrossRef]
- Grazieli, A.D.S. Studies on the Impact of cover Crops on Soybean Productivity and Root Rot Disease. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2019. Available online: https://lib.dr.iastate.edu/cgi/viewcontent.cgi?article=8396&context=etd (accessed on 28 April 2021).
- Ristaino, J.B.; Johnston, S.A. Ecologically based approaches to management of Phytophthora blight on bell pepper. Plant Dis. 1999, 83, 1080–1089. [Google Scholar] [CrossRef] [Green Version]
- McGiffin, M. Cowpea (Vigna unguiculata) Cover Crop; University of California’s Sustainable Agriculture Research and Extension Programs: Davis, CA, USA, 2005; Available online: http://ceriverside.ucanr.edu/files/96022.pdf (accessed on 2 November 2020).
- Cohen, M.F.; Yamasaki, H.; Mazzola, M. Brassica napus seed meal soil amendment modifies microbial community structure, nitric oxide production and incidence of Rhizoctonia root rot. Soil Biol. Biochem. 2005, 37, 1215–1227. [Google Scholar] [CrossRef]
- Motisi, N.; Montfort, F.; Faloya, V.; Lucas, P.; Dore, T. Growing Brassica juncea as a cover crop, then incorporating its residues provides complementary control of Rhizoctonia root rot of sugar beet. Field Crop Res. 2009, 113, 238–245. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Liyanapathiranage, P.; Addesso, M. Effect of Brassica crop-based biofumigation on soilborne disease suppression in woody ornamentals. Can. J. Plant Pathol. 2019, 8, 138. [Google Scholar] [CrossRef]
- Turrini, A.; Sbrana, C.; Avio, L.; Njeru, E.M.; Bocci, G.; Bàrberi, P.; Giovannetti, M. Changes in the composition of native root arbuscular mycorrhizal fungal communities during a short-term cover crop-maize succession. Biol. Fertil. Soils 2016, 53, 643–653. [Google Scholar] [CrossRef]
- Lehman, R.M.; Taheri, W.I.; Osborne, S.L.; Buyer, J.S.; Douds, D.D., Jr. Fall cover cropping can increase arbuscular mycorrhizae in soils supporting intensive agricultural production. Agric. Ecosyst. Environ. Appl. Soil Ecol. 2012, 61, 300–304. [Google Scholar] [CrossRef]
- Pfleger, F.L.; Linderman, R.G. Mycorrhiza and Plant Growth; The American Phytopathological Society Press: St. Paul, MN, USA, 2000; p. 360. [Google Scholar]
- Plotkin, J. Use of cover crops and green manures to attract beneficial insects. In Integrated Pest Management Program; University of Connecticut: Storrs, CT, USA, 1999; Retrieved: 11 May 2020. [Google Scholar]
- Vukicevich, E.; Lowery, T.; Bowen, P.; Urbez-Torres, J.R.; Hart, M. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agron. Sustain. Dev. 2016, 36, 48. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Cover Crop | Scientific Name | Seed Source | Growing Season | Germination Rate (%) * |
|---|---|---|---|---|
| Crimson clover | Trifolium incarnatum L. | Summitville Grain & Feed | Cool | 87.5 |
| Triticale | ×Triticosecale Wittm. ex A. Camus. | Summitville Grain & Feed | Cool | 97.5 |
| Austrian winter pea | Pisum sativum var. arvense (L.) Poir | Summitville Grain & Feed | Cool | 100.0 |
| Annual ryegrass | Lolium multiflorum L. | Summitville Grain & Feed | Cool | 100.0 |
| Red clover ‘Kenland’ | T. pratense L. | Summitville Grain & Feed | Cool | 82.5 |
| White clover | T. repens L. | Summitville Grain & Feed | Cool | 22.5 |
| Buckwheat | Fagopyrum esculentum Moench | Summitville Grain & Feed | Warm | 100.0 |
| Turnip ‘Pointer’ | Brassica rapa subsp. rapa L. | Summitville Grain & Feed | Cool | 67.5 |
| Tillage radish ‘Daikon’ | Raphanus sativus L. | Summitville Grain & Feed | Cool | 97.5 |
| Cowpea ‘Iron and Clay’ | Vigna unguiculata (L.) Walp. | Summitville Grain & Feed | Warm | 95.0 |
| Japanese millet | Echinochloa esculenta (A.Braun) H.Scholz | Summitville Grain & Feed | Warm | 97.5 |
| Browntop millet | Urochloa ramosa L. | Summitville Grain & Feed | Warm | 62.5 |
| Proso millet | Panicum miliaceum L. | Summitville Grain & Feed | Warm | 25.0 |
| Pearl millet | Pennisetum glaucum (L.) R. Br. | Summitville Grain & Feed | Warm | 62.5 |
| Treatment | Height of Cover Crops (cm) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annual Ryegrass | Austrian Winter Pea | Buckwheat | Cowpea ‘Iron and Clay’ | Crimson Clover | Tillage Radish ‘Daikon’ | Japanese Millet | Red Clover ‘Kenland’ | Triticale | ||||||||||
| Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | |
| Phytophthora nicotianae | 20.9 ± 0.7 b X | 15.9 ± 0.8 bc | 31.4 ± 0.7 bc | 24.6 ± 1.9 b | 22.2 ± 0.6 a | 20.9 ± 0.8 a | 18.5 ± 0.9 a | 27.6 ± 0.9 a | 8.5 ± 0.1 a | 6.8 ± 0.2 a | 13.9 ± 0.6 a | 10.2 ± 0.4 a | 25.6 ± 0.7 a | 18.7 ± 0.4 a | 4.7 ± 0.1 a | 4.63 ± 0.1 a | 24.9 ± 0.7 b | 18.9 ± 0.7 b |
| Rhizoctonia solani | 21.7 ± 0.7 a | 19.1 ± 0.8 b | 27.0 ± 0.5 c | 28.8 ± 1.8 ab | 21.5 ± 1.2 a | 22.7 ± 2.4 a | 20.0 ± 1.4 a | 14.5 ± 2.0 b | 8.6 ± 0.2 a | 5.6 ± 0.6 ab | 13.7 ± 0.7 a | 10.7 ± 0.5 a | 30.7 ± 5.7 a | 20.2 ± 1.3 a | 5.2 ± 0.4 a | 4.76 ± 0.1 a | 24.3 ± 0.8 b | 24.2 ± 1.9 a |
| Phytopythium vexans | 20.6 ± 0.6 b | 14.6 ± 0.2 c | 36.9 ± 2.8 ab | 26.97 ± 2.5 ab | 23.9 ± 1.2 a | 20.7 ± 2.4 a | 18.4 ± 1.7 a | 23.1 ± 1.5 a | 7.7 ± 0.2 b | 5.1 ± 0.2 b | 14.0 ± 0.6 a | 8.9 ± 0.5 a | 23.2 ± 1.8 a | 18.3 ± 0.7 a | 4.9 ± 0.1 a | 4.78 ± 0.4 a | 30.7 ± 1.5 a | 18.7 ± 0.7 b |
| Control | 20.6 ± 1.0 b | 16.9 ± 0.5 ab | 40.8 ± 0.3 a | 33.4 ± 1.6 a | 21.9 ± 0.1 a | 22.7 ± 1.3 a | 16.2 ± 0.9 a | 24.1 ± 0.8 a | 6.9 ± 0.2 c | 5.9 ± 0.1 ab | 13.9 ± 0.6 a | 9.6 ± 0.5 a | 26.6 ± 0.9 a | 17.8 ± 0.3 a | 4.5 ± 0.1 a | 4.21 ± 0.2 a | 27.8 ± 2.3 ab | 20.2 ± 0.6 ab |
| F value | 0.41 | 10.5 | 17.55 | 3.56 | 1.2 | 0.32 | 1.52 | 16.08 | 17.98 | 4.46 | 0.31 | 2.71 | 1.05 | 1.73 | 1.63 | 1.47 | 4.03 | 5.37 |
| df | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| p value | 0.7479 | 0.0002 | <0.0001 | 0.0327 | 0.3351 | 0.8099 | 0.2403 | <0.001 | <0.0001 | 0.0148 | 0.8169 | 0.0726 | 0.3942 | 0.1934 | 0.2135 | 0.2521 | 0.0216 | 0.0071 |
| Treatment | Total Plant Fresh Weight of Cover Crops (g) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annual Ryegrass | Austrian Winter Pea | Buckwheat | Cowpea ‘Iron and Clay’ | Crimson Clover | Tillage Radish ‘Daikon’ | Japanese Millet | Red Clover ‘Kenland’ | Triticale | ||||||||||
| Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | |
| Phytophthora nicotianae | 4.0 ± 0.2 a x | 3.8 ± 0.6 ab | 16.2 ± 1.1 a | 14.9 ± 1.6 b | 7.2 ± 0.6 ab | 4.1 ± 0.9 a | 10.2 ± 0.4 a | 27.4 ± 2.7 a | 2.5 ± 0.1 a | 2.0 ± 0.3 a | 11.6 ± 1.1 a | 9.5 ± 0.6 ab | 6.9 ± 0.5 a | 3.9 ± 0.4 ab | 1.0 ± 0.1 a | 1.9 ± 0.2 ab | 10.1 ± 0.5 a | 11.6 ± 1.9 ab |
| Rhizoctonia solani | 3.9 ± 0.5 ab | 4.5 ± 0.4 a | 9.6 ± 0.8 c | 21.0 ± 1.9 a | 3.5 ± 0.4 c | 3.4 ± 0.5 a | 5.4 ± 1.2 b | 10.7 ± 1.6 b | 2.9 ± 0.4 a | 1.0 ± 0.2 b | 7.0 ± 1.3 b | 10.3 ± 0.6 a | 6.5 ± 0.9 a | 5.2 ± 0.2 a | 0.7 ± 0.1 a | 1.2 ± 0.2 c | 8.6 ± 1.3 a | 7.3 ± 1.9 b |
| Phytopythium vexans | 2.7 ± 0.4 ab | 2.8 ± 0.2 b | 12.3 ± 1.1 bc | 15.4 ± 1.1 b | 8.3 ± 0.5 a | 3.4 ± 0.6 a | 10.2 ± 0.9 a | 25.5 ± 2.9 a | 3.8 ± 0.5 a | 1.1 ± 0.2 b | 7.5 ± 1.5 ab | 7.3 ± 0.7 b | 3.3 ± 0.6 b | 3.7 ± 0.3 b | 0.8 ± 0.1 a | 1.3 ± 0.3 c | 6.8 ± 0.8 a | 9.7 ± 0.7 ab |
| Control | 2.4 ± 0.4 b | 4.1 ± 0.3 ab | 13.4 ± 0.3 ab | 21.6 ± 3.1 a | 4.9 ± 0.9 bc | 7.2 ± 0.5 a | 9.9 ± 0.6 a | 21.9 ± 2.0 a | 2.7 ± 0.3 a | 1.3 ± 0.2 ab | 8.7 ± 0.5 ab | 9.9 ± 0.6 a | 6.6 ± 0.4 a | 3.2 ± 0.4 b | 0.9 ± 0.1 a | 2.1 ± 0.3 a | 9.2 ± 1.2 a | 16.3± 2.6 a |
| F value | 4.35 | 3.51 | 10.06 | 3.12 | 12.61 | 7.44 | 8.18 | 10.47 | 2.71 | 5.01 | 3.15 | 4.45 | 7.97 | 6.48 | 2.07 | 3.72 | 1.97 | 4.06 |
| df | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| p value | 0.0163 | 0.0341 | 0.0003 | 0.0488 | <0.0001 | 0.0016 | 0.0009 | 0.0002 | 0.0724 | 0.0094 | 0.0478 | 0.015 | 0.0011 | 0.003 | 0.1365 | 0.0282 | 0.1507 | 0.021 |
| Treatment | Total Plant Root Weight of Cover Crops (g) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annual Ryegrass | Austrian Winter Pea | Buckwheat | Cowpea ‘Iron and Clay’ | Crimson Clover | Tillage Radish ‘Daikon’ | Japanese Millet | Red Clover ‘Kenland’ | Triticale | ||||||||||
| Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | |
| Phytophthora nicotianae | 2.2 ± 0.1 a x | 3.1 ± 0.5 a | 9.2 ± 0.6 a | 11.0 ± 1.1 a | 0.6 ± 0.1 a | 1.4 ± 0.3 ab | 2.8 ± 0.2 a | 12.2 ± 1.0 ab | 1.5 ± 0.1 a | 1.1 ± 0.2 a | 1.0 ± 0.2 a | 4.8 ± 0.4 a | 3.8 ± 0.6 a | 3.1 ± 0.4 a | 0.5 ± 0.1 a | 1.1 ± 0.1 ab | 8.2 ± 0.4 a | 10.5 ± 1.8 a |
| Rhizoctonia solani | 2.1 ± 0.3 a | 3.7 ± 0.4 a | 5.8 ± 0.7 b | 14.6 ± 1.6 a | 0.3 ± 0.0 a | 1.0 ± 0.2 b | 1.5 ± 0.3 c | 4.1 ± 0.7 c | 1.4 ± 0.2 a | 0.4 ± 0.1 b | 0.9 ± 0.4 a | 4.1 ± 0.5 a | 3.5 ± 0.6 a | 2.9 ± 0.2 a | 0.4 ± 0.1 a | 0.7 ± 0.1 c | 6.6 ± 1.2 ab | 6.3 ± 1.8 a |
| Phytopythium vexans | 1.4 ± 0.3 ab | 2.2 ± 0.2 a | 6.4 ± 0.8 ab | 11.0 ± 0.7 a | 1.0 ± 0.4 a | 0.8 ± 0.3 b | 1.6 ± 0.3 bc | 13.1 ± 1.1 a | 2.0 ± 0.3 a | 0.6 ± 0.1 b | 1.0 ± 0.3 a | 3.0 ± 0.5 a | 1.1 ± 0.2 b | 2.1 ± 0.2 ab | 0.4 ± 0.1 a | 0.8 ± 0.2 c | 4.6 ± 0.6 b | 8.4 ± 0.7 a |
| Control | 1.1 ± 0.2 b | 3.0 ± 0.3 a | 6.9 ± 0.8 ab | 13.2 ± 1.9 a | 1.0 ± 0.3 a | 2.0 ± 0.3 a | 2.5 ± 0.3 ab | 8.9 ± 0.9 b | 1.7 ± 0.2 a | 0.8 ± 0.1 ab | 1.0 ± 0.1 a | 4.1 ± 0.4 a | 3.8 ± 0.1 a | 1.6 ± 0.3 b | 0.5 ± 0.0 a | 1.2 ± 0.2 a | 5.8 ± 0.9 ab | 12.0 ± 1.2 a |
| F value | 4.77 | 3 | 4.17 | 1.66 | 1.75 | 4.43 | 6.68 | 19.34 | 1.73 | 4.97 | 0.02 | 2.87 | 9.53 | 6.44 | 0.91 | 3.19 | 3.51 | 3.05 |
| df | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| p value | 0.0115 | 0.055 | 0.019 | 0.2085 | 0.1886 | 0.0152 | 0.0026 | < 0.001 | 0.1928 | 0.0098 | 0.9953 | 0.0622 | 0.0004 | 0.0031 | 0.4524 | 0.0459 | 0.034 | 0.0525 |
| Cover Crop | Germination Rate of Different Cover Crops with the Respective Treatments (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Phytophthora nicotianae | Phytopythium vexans | Rhizoctonia solani | Control | |||||
| Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | |
| Annual Ryegrass | 95.0 ± 2.2 a x | 73.3 ± 8.8 a–d | 85.0 ± 5.0 ab | 90.0 ± 2.6 ab | 91.7 ± 6.5 a | 81.7 ± 6.0 abc | 95.0 ± 2.2 a | 95.0 ± 3.4 a |
| Austrian Winter Pea | 96.7 ± 3.3 a | 98.3 ± 1.7 a | 88.3 ± 6.0 a | 76.7 ± 6.7 abc | 83.3 ± 5.6 ab | 91.7 ± 3.1 a | 81.7 ± 3.1 ab | 95.0 ± 2.2 a |
| Buckwheat | 88.3 ± 7.5 a | 53.3 ± 10.2 cd | 56.7 ± 8.8 cd | 40.0 ± 5.8 de | 28.3 ± 3.1 d | 30.0 ± 4.5 d | 75.0 ± 8.5 ab | 71.7 ± 4.8 b |
| Cowpea ‘Iron and Clay’ | 96.7 ± 2.1 a | 86.7 ± 7.6 ab | 81.7 ± 3.1 abc | 83.3 ± 4.9 abc | 45.0 ± 7.6 cd | 55.0 ± 7.2 cd | 81.7 ± 4.0 ab | 93.3 ± 3.3 a |
| Crimson Clover | 86.7 ± 3.3 a | 51.7 ± 3.1 d | 76.7 ± 4.9 a-d | 31.7 ± 4.8 e | 75.0 ± 6.7 abc | 28.3 ± 4.0 d | 91.7 ± 4.8 ab | 71.7 ± 4.0 b |
| Tillage Radish ‘Daikon’ | 96.7 ± 2.1 a | 80 ± 5.8 abc | 53.3 ± 5.6 d | 56.7 ± 10.5 b-e | 65.0 ± 9.2 abc | 70.0 ± 11.0 abc | 81.7 ± 3.1 ab | 96.7 ± 2.1 a |
| Japanese Millet | 96.7 ± 3.3 a | 98.3 ± 1.7 a | 95.0 ± 2.2 a | 95.0 ± 3.4 a | 95.0 ± 3.4 a | 88.3 ± 4.8 ab | 86.7 ± 4.9 ab | 96.7 ± 2.1 a |
| Red Clover ‘Kenland’ | 86.7 ± 4.9 a | 68.3 ± 6.0 bcd | 60.0 ± 6.8 bcd | 50.0 ± 14.8 cde | 53.3 ± 5.6 bcd | 85.0 ± 5.6 abc | 71.7 ± 4.8 b | 75.0 ± 5.0 b |
| Triticale | 86.7 ± 3.3 a | 90.0 ± 3.7 ab | 83.3 ± 4.9 ab | 75.0 ± 7.6 a-d | 68.3 ± 10.5 abc | 56.7 ± 12.6 bcd | 86.7 ± 4.9 ab | 95.0 ± 2.2 a |
| F value | 1.57 | 8.22 | 7.32 | 8.67 | 10.34 | 11.35 | 2.41 | 10.95 |
| df | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 |
| p value | 0.1594 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0295 | <0.0001 |
| Cover Crop | Post Emergence Damping-off of Different Cover Crops with the Respective Treatments (%) | |||||
|---|---|---|---|---|---|---|
| Phytophthora nicotianae | Phytopythium vexans | Rhizoctonia solani | ||||
| Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | |
| Annual Ryegrass | 0 a x | 0 b | 6.7 ± 3.3 a | 0 a | 1.7 ± 1.7 c | 0 a |
| Austrian Winter Pea | 1.7 ± 1.7 a | 0 b | 6.7 ± 2.1 a | 0 a | 5.0 ± 2.2 b | 0 a |
| Buckwheat | 3.3 ± 2.1 a | 0 b | 5.0 ± 2.2 a | 1.7 ± 1.7 a | 5.0 ± 2.2 b | 5.0 ± 2.2 a |
| Cowpea ‘Iron and Clay’ | 5.0 ± 3.4 a | 3.3± 2.1 a | 8.3 ± 4.0 a | 3.3 ± 2.1 a | 13.3 ± 5.6 a | 0 a |
| Crimson Clover | 8.3 ± 3.1 a | 0 b | 6.7 ± 3.3 a | 0 a | 10.0 ± 3.7 b | 1.7 ± 1.7 a |
| Tillage Radish ‘Daikon’ | 6.7 ± 3.3 a | 0 b | 5.0 ± 2.2 a | 0 a | 13.3 ± 3.3 a | 3.3 ± 3.3 a |
| Japanese Millet | 6.7 ± 2.1 a | 0 b | 3.3 ± 2.1 a | 0 a | 1.7 ± 1.7 c | 0 a |
| Red Clover ‘Kenland’ | 10.0 ± 3.7 a | 0 b | 0 a | 1.7 ± 1.7 a | 6.7 ± 2.1 b | 6.7 ± 2.1 a |
| Triticale | 8.3 ± 4.0 a | 3.33 ± 2.1 a | 0 a | 1.7 ± 1.7 a | 6.7 ± 2.1 b | 1.7 ± 1.7 a |
| F value | 1.36 | 2.19 | 1.4 | 1.03 | 2.17 | 2.1 |
| df | 8 | 8 | 8 | 8 | 8 | 8 |
| p value | 0.2383 | 0.0465 | 0.2245 | 0.4263 | 0.0482 | 0.0555 |
| Plant Growth Parameter | Root Rot Disease Severity (%) | |||||
|---|---|---|---|---|---|---|
| Phytopythium vexans | Phytophthora nicotianae | Rhizoctonia solani | ||||
| Trial 1 | Trial 2 | Trial 1 | Trial 2 | Trial 1 | Trial 2 | |
| Height | R2 = 0.01630 p = 0.9069 | R2 = −0.28598 p = 0.0379 | R2 = 0.25443 p = 0.0634 | R2 = −0.213 p = 0.1217 | R2 = −0.18175 p = 0.1884 | R2 = −0.43242 p = 0.0011 |
| Total fresh weight | R2 = −0.09847 p = 0.4787 | R2 = −0.43648 p = 0.0011 | R2 = 0.18645 p = 0.1770 | R2 = −0.381 p = 0.0045 | R2 = −0.37014 p = 0.0059 | R2 = −0.55140 p = <0.0001 |
| Total root weight | R2 = −0.17408 p = 0.2080 | R2 = −0.50503 p = 0.0001 | R2 = −0.06612 p = 0.6348 | R2 = −0.496 p = 0.0001 | R2 = −0.45395 p = 0.0006 | R2 = −0.56087 p = <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panth, M.; Witcher, A.; Baysal-Gurel, F. Response of Cover Crops to Phytopythium vexans, Phytophthora nicotianae, and Rhizoctonia solani, Major Soilborne Pathogens of Woody Ornamentals. Agriculture 2021, 11, 742. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080742
Panth M, Witcher A, Baysal-Gurel F. Response of Cover Crops to Phytopythium vexans, Phytophthora nicotianae, and Rhizoctonia solani, Major Soilborne Pathogens of Woody Ornamentals. Agriculture. 2021; 11(8):742. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080742
Chicago/Turabian StylePanth, Milan, Anthony Witcher, and Fulya Baysal-Gurel. 2021. "Response of Cover Crops to Phytopythium vexans, Phytophthora nicotianae, and Rhizoctonia solani, Major Soilborne Pathogens of Woody Ornamentals" Agriculture 11, no. 8: 742. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080742