
Effects of Dietary Supplementation of Humic Substances on Production Parameters, Immune Status and Gut Microbiota of Laying Hens

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Production Parameters Screening
2.3. Egg Shell Analysis
2.4. Homogenization of Cecum and Isolation of Total RNA of IgA, IGF-2 and MUC-2 Genes
2.5. Relative Expression of IgA, IGF-2 and MUC-2 Genes in Quantitative Real-Time PCR (qRT-PCR)
2.6. Phagocyte Activity Testing
2.7. Identification of Lymphocyte Subpopulations
2.8. Flow Cytometric Analysis
2.9. Intestinal Bacteria Analysis
2.10. Statistical Analysis
3. Results
3.1. Production Parameters
3.2. Eggshell Mineral Analysis
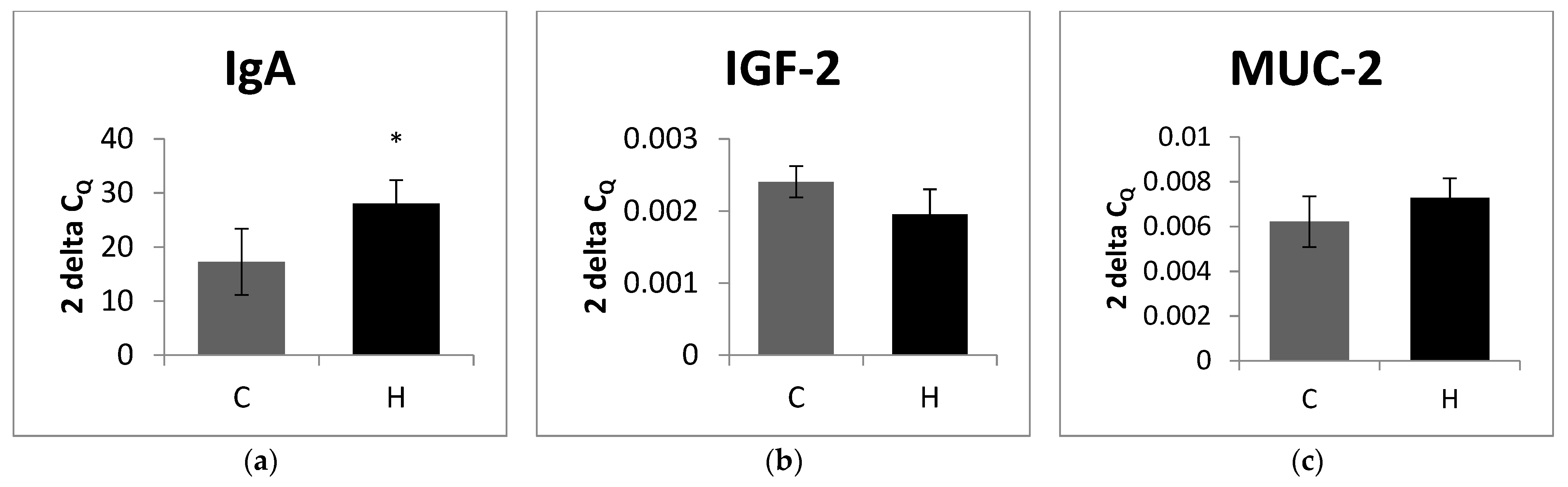
3.3. The Relative Expression of Selected Genes
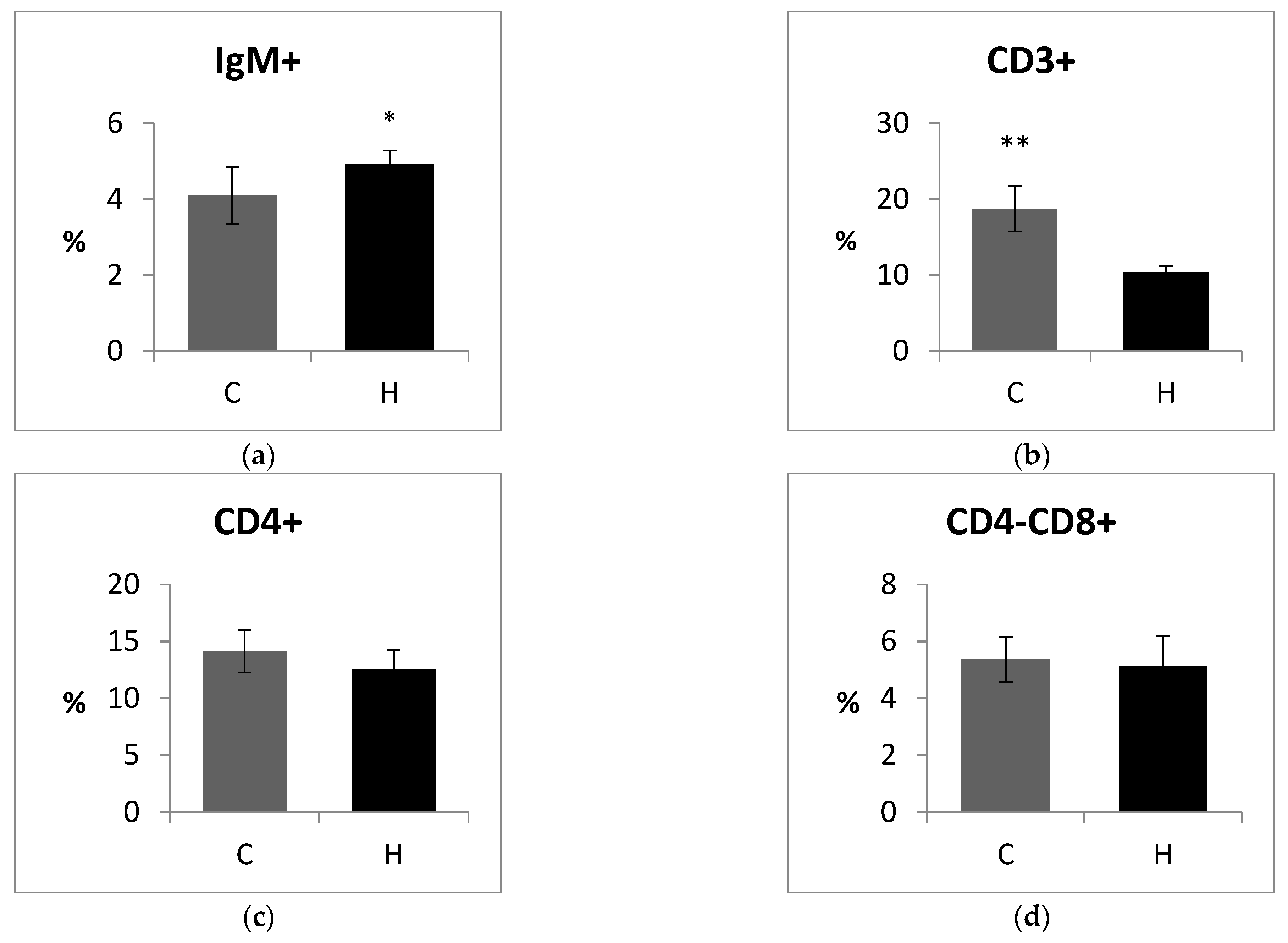
3.4. Cellular Immune Response
3.5. Intestinal Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, P.; Xu, C.; Xing, W.; Sun, L.; Kaplan, D.I.; Fujitake, N.; Yeager, C.M.; Schwehr, K.A.; Santschi, P.H. Radionuclide uptake by colloidal and particulate humic acids obtained from 14 soils collected worldwide. Sci. Rep. 2018, 8, 4795. [Google Scholar] [CrossRef] [PubMed]
- Mihajlović, V.; Tomić, T.; Tubić, A.; Jazić, J.M.; Tubmas, I.I.; Sunjka, D.; Lacić, S.; Teodorović, I. The impact of humic acid on toxicity of individual herbicides and their mixtures to aquatic macrophytes. Environ. Sci. Pollut. Res. 2019, 26, 23571–23582. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wen, J.; Hu, Y.; Fang, Y.; Zhang, H.; Xing, L.; Wang, Y.; Zeng, G. Humic substances from green waste compost: An effective washing agent for heavy metal (Cd, Ni) removal from contaminated sediments. J. Hazard. Mater. 2019, 366, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Volkov, I.V.; Polyakov, E.V. Interaction of humic acids with microelements/radionuclides in sorption systems. Radiochemistry 2020, 62, 141–160. [Google Scholar] [CrossRef]
- Maruf, M.T.; Rasul, G.A.M. Influence of humic acid and sulfur on the bioavailability of some micronutrients in calcareous soils. Plant Arch. 2019, 19, 1785–1794. [Google Scholar]
- Edmonds, M.S.; Moreland, J.S. Effect of supplemental humic and butyric acid on performance and mortality in broilers raised under various environmental conditions. J. Appl. Poult. Res. 2014, 23, 260–267. [Google Scholar] [CrossRef]
- Arif, M.; Alagawany, M.; Abd El-Hack, M.E.; Saeed, M.; Arain, M.A.; Elnesr, S.S. Humic acid as a feed additive in poultry diets: A review. Iran. J. Vet. Res. 2019, 20, 167–172. [Google Scholar] [PubMed]
- van Rensburg, C.E. The Antiinflammatory properties of humic substances: A mini review. Phytother. Res. 2015, 29, 791–795. [Google Scholar] [CrossRef] [Green Version]
- Marcinčáková, D.; Mačanga, J.; Nagy, J.; Marcinčák, S.; Popelka, P.; Vašková, J.; Jaďuttová, I.; Mellen, M. Effect of supplementation of the diet with humic acids on growth performance and carcass yield of broilers. Folia Vet. 2015, 59, 165–168. [Google Scholar]
- Domínguez-Negrete, A.; Gómez-Rosales, S.; Angeles, M.d.L.; López-Hernández, L.H.; Reis-de Souza, T.C.; López-García, Y.; Zavala-Franco, A.; Téllez-Isaias, G. Effect of the addition of humic substances as growth promoter in broiler chickens under two feeding regimens. Animals 2019, 9, 1101. [Google Scholar] [CrossRef] [Green Version]
- Semjon, B.; Marcinčáková, D.; Koréneková, B.; Bartkovský, M.; Nagy, J.; Turek, P.; Marcinčák, S. Multiple factorial analysis of physicochemical and organoleptic properties of breast and thigh meat of broilers fed a diet supplemented with humic substances. Poult. Sci. 2020, 99, 1750–1760. [Google Scholar] [CrossRef]
- Hudák, M.; Semjon, B.; Marcinčáková, D.; Bujňák, L.; Na’, P.; Koréneková, B.; Nagy, J.; Bartkovský, M.; Marcinčák, S. Effect of Broilers Chicken Diet Supplementation with Natural and Acidified Humic Substances on Quality of Produced Breast Meat. Animals 2021, 11, 1087. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Hincke, M.T.; Prithiviraj, B.; Critchley, A. A review of the varied uses of macroalgae as dietary supplements in selected poultry with special reference to laying Hen and broiler chickens. J. Mar. Sci. Eng. 2020, 8, 536. [Google Scholar] [CrossRef]
- Hahn, J.; Dehghan, M.; Drouin-Chartier, J.P.; Mentz, R.J.; Jneid, H.; Virani, S.S.; Wilson Tang, W.H.; Krittanawong, C. Egg consumption and risk of cardiovascular disease: A critical review. Curr. Emerg. Hosp. Med. Rep. 2021, 9, 1–13. [Google Scholar] [CrossRef]
- Wan, Y.; Ma, R.; Khalid, A.; Chai, L.; Qi, R.; Liu, W.; Li, J.; Li, Y.; Zhan, K. Effect of the pellet and mash feed forms on the productive performance, egg quality, nutrient metabolism, and intestinal morphology of two laying hen breeds. Animals 2021, 11, 701. [Google Scholar] [CrossRef]
- Wang, J.; Yue, H.; Wu, S.; Zhang, H.; Qi, G. Nutritional modulation of health, egg quality and environmental pollution of the layers. Anim. Nutr. 2017, 3, 91–96. [Google Scholar] [CrossRef]
- Arafat, R.Y.; Khan, S.H.; Abbas, G.; Iqbal, J. Effect of dietary humic acid via drinking water on the performance and egg quality of commercial layers. Am. J. Life Sci. 2015, 3, 26–30. [Google Scholar]
- Vlčková, J.; Tůmová, E.; Míková, K.; Englmaierová, M.; Okrouhlá, M.; Chodová, D. Changes in the quality of eggs during storage depending on the housing system and the age of hens. Poult. Sci. 2019, 98, 6187–6193. [Google Scholar] [CrossRef] [PubMed]
- Nagaraju, R.; Reddy, B.S.; Gloridoss, R.; Suresh, B.N.; Ramesh, C. Effect of dietary supplementation of humic acids on performance of broilers. Indian J. Anim. Sci. 2014, 84, 447–452. [Google Scholar]
- Mudroňová, D.; Karaffová, V.; Pešulová, T.; Koščová, J.; Maruščáková, I.C.; Bartkovský, M.; Marcinčáková, D.; Ševčíková, Z.; Marcinčák, S. The effect of humic substances on gut microbiota and immune response of broilers. Food Agric. Immunol. 2020, 31, 137–149. [Google Scholar] [CrossRef]
- Hanafy, M.M.; El-Sheikh, A.M.H. The effect of dietary humic acid supplementation on some productive and physiological traits of laying hens. Egypt. Poult. Sci. 2018, 28, 1043–1058. [Google Scholar]
- Arif, M.; Rehman, A.; El-Hack, M.E.A.; Saeed, M.; Khan, F.; Akhtar, M.; Swelum, A.A.; Saadeldin, I.M.; Alowaimer, A.N. Growth, carcass traits, cecal microbial counts, and blood chemistry of meat-type quail fed diets supplemented with humic acid and black cumin seeds. Asian Aust. J. Anim. Sci. 2018, 31, 1930–1938. [Google Scholar] [CrossRef] [Green Version]
- Islam, K.; Schuhmacher, A.; Gropp, J. Humic acid substances in animal agriculture. Pak. J. Nutr. 2005, 4, 126–134. [Google Scholar]
- Arpášová, H.; Kačániová, M.; Pistová, V.; Gálik, B.; Fik, M.; Hleba, L. Effect of probiotics and humic Aaid on egg production and quality parameters of laying hens eggs. Sci. Pap. Anim. Sci. Biotechnol. 2016, 49, 1–9. [Google Scholar]
- Sopoliga, I.; Hreško-Šamudovská, A.; Demeterová, M.; Na’, P.; Marcin, A.; Skalická, M. Effect of humic substances on the production parameters of pheasant hens. Acta Fytotech. Zootech. 2016, 19, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Lohmann-Breeders Home Page. Available online: https://www.ltz.de/de-wAssets/docs/management-guides/en/Cage/Brown/LTZ-Management-Guide-LB-Classic-EN.pdf (accessed on 7 June 2021).
- An Official Website of the European Union. Available online: https://op.europa.eu/en/publication-detail/-/publication/72709682-c5e2-42a4-948d-1877344bb582/language-en (accessed on 7 June 2021).
- Semjon, B.; Bartkovský, M.; Marcinčáková, D.; Klempová, T.; Bujňák, L.; Hudák, M.; Ja’uttová, I.; Čertík, M.; Marcinčák, S. Effect of Solid-State Fermented Wheat Bran Supplemented with Agrimony Extract on Growth Performance, Fatty Acid Profile, and Meat Quality of Broiler Chickens. Animals 2020, 10, 942. [Google Scholar] [CrossRef] [PubMed]
- Skalicka, M.; Korenekova, B.; Na’, P.; Šály, J. Influence of chromium and cadmium addition on quality of Japanese quail eggs. Acta Vet. Brno. 2008, 77, 503–508. [Google Scholar] [CrossRef] [Green Version]
- Associatition of Official Analytical Chemists International. Official Methods of Analysis, 17th ed.; AOAC: Arlington, TX, USA, 2001. [Google Scholar]
- Karaffová, V.; Bobíková, K.; Levkut, M.; Revajová, V.; Ševčíková, Z.; Levkut, M. The influence of Farmatan® and Flimabend® on the mucosal immunity of broiler chicken. Poult. Sci. 2019, 98, 1161–1166. [Google Scholar] [CrossRef]
- Lammers, A.; Wieland, W.H.; Kruijt, L.; Jansma, A.; Straetemans, T.; Schots, A.; den Hartog, G.; Parmentier, H.K. Successive immunoglobulin and cytokine expression in the small intestine of juvenile chicken. Dev. Comp. Immunol. 2010, 34, 1254–1262. [Google Scholar] [CrossRef]
- Smirnov, A.; Tako, E.; Ferket, P.R.; Uni, Z. Mucin gene expression and mucin content in the chicken intestinal goblet cells are affected by in ovo feeding of carbohydrates. Poult Sci. 2006, 85, 669–673. [Google Scholar] [CrossRef]
- Mudronová, D.; Karaffová, V.; Košcová, J.; Bartkovský, M.; Marcincáková, D.; Popelka, P.; Klempová, T.; Certík, M.; Macanga, J.; Marcincák, S. Effect of fungal gamma-linolenic acid and beta-carotene containing prefermented feed on immunity and gut of broiler chicken. Poult. Sci. 2018, 97, 4211–4218. [Google Scholar] [CrossRef]
- De Boever, S.; Vangestel, C.; De Backer, P.; Croubels, S.; Sys, S.U. Identification and validation of housekeeping genes as internal control for gene expression in an intravenous LPS inflammation model in chickens. Vet. Immunol. Immunopathol. 2008, 122, 312–317. [Google Scholar] [CrossRef]
- BD Biosciences Home Page. Available online: https://www.bdbiosciences.com/ds/pm/tds/341060.pdf (accessed on 7 June 2021).
- Bertram, E.M. Characterization of duck thrombocytes. Res. Vet. Sci. 1998, 64, 267–270. [Google Scholar] [CrossRef]
- Zhang, A.R.; Pirzado, S.A.; Liu, G.H.; Chen, Z.M.; Chang, W.H.; Cai, H.Y.; Bryden, W.L.; Zheng, A.J. Dietary supplementation with sodium humate improves egg quality and immune function of laying hens. J. Appl. Anim. Nutr. 2020, 2, 93–99. [Google Scholar] [CrossRef]
- Aksu, T.; Bozkurt, A.S. Effect of dietary essential oils and/or humic acids on broiler performance, microbial population of intestinal content and antibody titres in the summer season. Kafkas Univ. Vet. Fak. Derg. 2009, 15, 185–190. [Google Scholar]
- Kocabağli, N.; Alp, M.; Acar, N.; Kahraman, R. The effects of dietary humate supplementation on broiler growth and carcass yield. Poult. Sci. 2002, 81, 227–230. [Google Scholar] [CrossRef]
- Rath, N.C.; Huff, W.E.; Huff, G.R. Effects of humic acid on broiler chickens. Poult. Sci. 2006, 85, 410–414. [Google Scholar] [CrossRef]
- Kiczorowska, B.; Samolinska, W.; Kwiecien, M.; Winiarska-Mieczan, A.; Rusinek-Prystupa, E.; Al-Yasiry, A.R.M. Nutritional value and the content of minerals in eggs produced in large-scale, courtyard and organic systems. J. Elem. 2015, 20, 887–895. [Google Scholar] [CrossRef]
- Klöcking, R. Humic substances as potential therapeutics. In Humic Substances in the Global Environment; Senesi, N., Miano, T.M., Eds.; Elsevier: Amsterdam, The Netherlands, 1994; pp. 1245–1257. [Google Scholar]
- Hassan, S.B.; Aigbodion, V.S.; Patrick, S.N. Development of polyester/eggshell particulate composites. Tribol. Ind. 2012, 34, 217–225. [Google Scholar]
- Ergin, O.; Coskun, I.; Ocak, N.; Erener, G. Effects of dietary humic substances on egg production and egg shell quality of hens after peak laying period. Afr. J. Biotechnol. 2009, 8, 1155–1159. [Google Scholar]
- Tancho, A. The Effect of Humic Acid Supplement on Egg Performance and Egg Quality of Laying Hen during 28–47 Weeks of Age. Available online: http://www.kmitl.ac.th/agritech30/example.doc (accessed on 7 June 2021).
- Weber, T.E.; van Sambeek, D.M.; Gabler, N.K.; Kerr, B.J.; Moreland, S.; Johal, S.; Edmonds, M.S. Effects of dietary humic and butyric acid on growth performance and response to lipopolysaccharide in young pigs. J. Anim. Sci. 2014, 92, 4172–4179. [Google Scholar] [CrossRef] [Green Version]
- Sanmiguel, R.P.; Rondón, I.B. Supplementation with humic substances affects the innate immunity in layer hens in posfasting phase. Rev. MVZ Córdoba 2016, 21, 5198–5210. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Liu, J.J.; Lu, F.J.; Yang, M.L.; Lee, Y.; Huang, T.S. The effect of humic acid on the adhesibility of neutrophils. Thromb. Res. 2002, 108, 67–76. [Google Scholar] [CrossRef]
- Riede, U.N.; Zeck-Kapp, G.; Freudenberg, N.; Keller, H.U.; Seubert, B. Humate-induced activation of human granulocytes. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1991, 60, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Luthala, M. Chicken CD4, CD8αβ, and CD8αα T cell co-receptor molecules. Poult. Sci. 1998, 77, 1858–1873. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.K.; Gu, M.J.; Ko, K.H.; Bae, S.; Kim, G.; Jin, G.D.; Kim, E.B.; Kong, Y.Y.; Park, T.S.; Jung, H.J.; et al. Regulation of CD4+CD8−CD25+ and CD4+CD8+CD25+ T cells by gut microbiota in chicken. Sci. Rep. 2018, 8, 8627. [Google Scholar] [CrossRef]
- Salah, H.; Masour, E.S.; Reham, R.R.; El Hamid, E.S.A. Study on the effect of humic acid on growth performance, immunological, some blood parameters and control intestinal closterdium in broiler chickens. Zag. Vet. J. 2015, 43, 102–109. [Google Scholar] [CrossRef]
- Vetvicka, V.; Baigorri, R.; Zamarreño, A.M.; Garcia-Mina, J.M.; Yvin, J.C. Glucan and Humic Acid: Synergistic Effects on the Immune System. J. Med. Food. 2010, 13, 863–869. [Google Scholar] [CrossRef]
- Semantic Scholar Home Page. Effects of Humic Acid on Animals and Humans. An Overview of Literature and a Review of Current Research. Available online: https://www.semanticscholar.org/paper/Effects-of-Humic-Acid-on-Animals-and-Humans-An-of-a/cc09ed93ae53f20b014315d0a53da6de278c36de (accessed on 7 June 2021).
- Shermer, C.L.; Maciorowski, K.G.; Bailey, C.A.; Byers, F.M.; Ricke, S.C. Caecal metabolites and microbial populations in chickens consuming diets containing a mined humate compound. J. Sci. Food Agric. 1998, 77, 479–486. [Google Scholar] [CrossRef]
- Buzoleva, L.S.; Sidorenko, M.L. Vliianie organicheskikh veshchestv guminovykh kislot na razmnozhenie énterobakteriĭ [Effect of organic composition of humic acids on Enterobacteria multiplication]. Zh. Mikrobiol. Epidemiol. Immunobiol. 2001, 2, 89–91. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Diet | |
| Corn grain, % | 27.50 | |
| Wheat grain, % | 31.00 | |
| Barley grain, % | 7.50 | |
| Soybean meal, % | 11.00 | |
| Corn gluten meal, % | 1.65 | |
| Rapeseed meal, % | 1.70 | |
| Sunflower cake, % | 3.20 | |
| Sunflower oil, % | 3.00 | |
| Wheat bran, % | 5.00 | |
| Limestone, % | 6.00 | |
| Monocalcium Phosphate, % | 1.25 | |
| Salt, % | 0.20 | |
| Premix of amino acids, vitamins, and minerals *, % | 1.00 | |
| Ingredients | As Fed | Dry matter |
| Dry matter, g·kg−1 | 899.00 | 1000.00 |
| Crude protein, g·kg−1 | 157.50 | 175.19 |
| Crude fat, g·kg−1 | 44.23 | 49.20 |
| Crude fiber, g·kg−1 | 48.48 | 53.93 |
| Ash, g·kg−1 | 128.60 | 143.05 |
| Starch, g·kg−1 | 414.80 | 461.40 |
| Calcium, g·kg−1 | 38.00 | 42.27 |
| Phosphorus, g·kg−1 | 4.80 | 5.34 |
| Sodium, g·kg−1 | 1.50 | 1.67 |
| Copper, mg·kg−1 | 18.08 | 20.11 |
| Zinc, mg·kg−1 | 65.90 | 73.30 |
| Manganese, mg·kg−1 | 87.55 | 97.39 |
| Primer | Sequence 5′–3′ | Annealing/Temperature Time | References |
|---|---|---|---|
| IgA For | GTCACCGTCACCTGGACTACA | 55 °C/30 s | [32] |
| IgA Rev | ACCGATGGTCTCCTTCACATC | ||
| Muc 2 For | GCTGATTGTCACTCACGCCTT | 54 °C/1 min | [33] |
| Muc 2 Rev | ATCTGCCTGAATCACAGGTGC | ||
| IGF-2 For | CTCTGCTGGAAACCTACTGT | 55 °C/30 s | [34] |
| IGF-2 Rev | GAGTACTTGGCATGAGATGG | ||
| GAPDH For | CCTGCATCTGCCCATTT | 59 °C/30 s | [35] |
| GAPDH Rev | GGCACGCCATCACTATC |
| Type | Fluorochrome | Clone | Isotype | Concentration | Amount/5 × 105 cells |
|---|---|---|---|---|---|
| anti-CD3 | FITC | CT-3 | IgG1 κ | 0.5 mg·mL−1 | 2 μL |
| anti-CD4 | FITC | CT-4 | IgG1, κ | 0.5 mg·mL−1 | 2 μL |
| anti-CD8a | R-PE | CT-8 | IgG1, κ | 0.1 mg·mL−1 | 1 μL |
| anti-CD45 | APC | LT-40 | IgM, κ | 0.1 mg·mL−1 | 5 μL |
| anti-IgM | R-PE | M-1 | IgG2b κ | 0.1 mg·mL−1 | 1 μL |
| Variable | C | H | p-Value |
|---|---|---|---|
| Laying rate, % | 84.29 ± 7.87 b | 95.91 ± 6.98 a | 0.013 |
| Daily egg mass, g/hen/day | 47.50 ± 4.37 b | 57.06 ± 4.05 a | 0.001 |
| Egg weight, g | 56.36 ± 0.39 b | 59.50 ± 0.67 a | <0.001 |
| Feed consumption, g/hen/day | 115.86 ± 2.27 | 116.00 ± 1.53 | 0.892 |
| Feed conversion ratio | 2.46 ± 0.22 b | 2.04 ± 0.14 a | 0.001 |
| Variable | C | H | p-Value |
|---|---|---|---|
| Calcium, g·kg−1 | 57.66 ± 0.27 b | 59.22 ± 1.09 a | 0.014 |
| Magnesium, g·kg−1 | 4.40 ± 0.25 | 4.67 ± 0.46 | 0.281 |
| Phosphorus, g·kg−1 | 2.69 ± 0.35 b | 4.77 ± 1.14 a | 0.005 |
| Sodium, g·kg−1 | 0.96 ± 0.04 | 0.91 ± 0.09 | 0.276 |
| Potassium, g·kg−1 | 0.61 ± 0.06 | 0.55 ± 0.10 | 0.306 |
| Copper, mg·kg−1 | 36.68 ± 4.22 b | 46.69 ± 1.90 a | 0.001 |
| Zinc, mg·kg−1 | 21.14 ± 6.59 b | 31.96 ± 8.09 a | 0.049 |
| Manganese, mg·kg−1 | 26.36 ± 2.09 b | 30.09 ± 1.57 a | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mudroňová, D.; Karaffová, V.; Semjon, B.; Naď, P.; Koščová, J.; Bartkovský, M.; Makiš, A.; Bujňák, L.; Nagy, J.; Mojžišová, J.; et al. Effects of Dietary Supplementation of Humic Substances on Production Parameters, Immune Status and Gut Microbiota of Laying Hens. Agriculture 2021, 11, 744. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080744
Mudroňová D, Karaffová V, Semjon B, Naď P, Koščová J, Bartkovský M, Makiš A, Bujňák L, Nagy J, Mojžišová J, et al. Effects of Dietary Supplementation of Humic Substances on Production Parameters, Immune Status and Gut Microbiota of Laying Hens. Agriculture. 2021; 11(8):744. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080744
Chicago/Turabian StyleMudroňová, Dagmar, Viera Karaffová, Boris Semjon, Pavel Naď, Jana Koščová, Martin Bartkovský, Andrej Makiš, Lukáš Bujňák, Jozef Nagy, Jana Mojžišová, and et al. 2021. "Effects of Dietary Supplementation of Humic Substances on Production Parameters, Immune Status and Gut Microbiota of Laying Hens" Agriculture 11, no. 8: 744. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080744