
Crosstalk between Nutrient Signalling Pathways and Immune Responses in Rice


1
Centre for Research in Agricultural Genomics (CRAG) CSIC-IRTA-UAB-UB, Campus Universitat Autònoma de Barcelona (UAB), 08193 Bellaterra (Cerdanyola del Vallés), Spain
2
Consejo Superior de Investigaciones Científicas (CSIC), 08016 Barcelona, Spain
*
Author to whom correspondence should be addressed.
†
Current affiliation: Laboratori Agroalimentari, Departament d’Agricultura, Ramaderia, Pesca i Alimentació (DARP), C/Camí de Mataró Núm. 1, 08348 Cabrils, Spain.
Agriculture 2021, 11(8), 747; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080747
Submission received: 15 July 2021
/
Revised: 1 August 2021
/
Accepted: 2 August 2021
/
Published: 6 August 2021
(This article belongs to the Special Issue Breeding and Genetics to Shape Rice Production Systems in the New Climatic Scenario)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Rice is a staple food for more than half of the global population. Rice production is, however, severely affected by biotic and abiotic stresses. Fertilisers and pesticides are widely used in rice farming to maintain optimal yield and to prevent losses caused by environmental stress. However, the indiscriminate use of agrochemicals has adverse effects on the environment and human health. Stress caused by nutrient excess or deficiency has an impact on plant disease resistance. The interference of plant responses induced by nutrient stress can result in a positive or negative impact on resistance to pathogen infection. In this review, we explore the effects of combined stresses in rice, focusing on nutrient stress, such as nitrogen and phosphorous supply, and infection by fungal pathogen Magnaporthe oryzae, the causal agent of rice blast disease. Crosstalk between nutrient stress-induced and pathogen-induced signalling pathways in rice is discussed, as well, with particular emphasis on regulatory microRNAs. Understanding the interconnected regulations between nutrient stress and disease resistance will lay a foundation for rationally optimising fertiliser and pesticide use in rice production.
1. Introduction
Rice (Oryza sativa L.) is one of the most important crops in the world and a staple food for more than half of the global population. China is the largest rice producer in the world, followed by India, Indonesia, and Bangladesh [1]. Rice is grown in a wide range of environments characterised by different climatic conditions, different cultivation systems such as paddy fields and rainfed uplands, and different irrigation or flooding conditions. To meet the forecasted demand of rice, posed by the increasing human population, rice production must significantly increase. This needs to be achieved within the declining arable land, due to urbanisation, which is often polluted by agrochemicals and industrialisation. Additionally, rice farmers are prone to face climate change and water scarcity factors.
Rice, a monocotyledonous plant species, originated around 9000 years ago in the Yangtze Valley (China) and it has a long history of natural selection and domestication [2]. It belongs to the genus Oryza which includes the two cultivated species, the Asian rice (Oryza sativa) and the African rice (Oryza glaberrima). O. sativa comprises japonica and indica subspecies [3,4,5,6]. O. rufipogon and O. nivara are considered to be the progenitors of the Asian cultivated rice, whereas O. glaberrima originated from its wild ancestor O. barthii [7,8]. O. sativa was the first crop species which was genome sequenced, and reference genomes are available for japonica and indica subspecies [9,10]. More recently, the 3 K Rice Genome Project sequenced more than 3000 rice accessions which provided a tremendous resource for rice research [11]. Rice has been adopted as the model plant for functional genomics in cereal species. Progress in agricultural research is needed for the development of necessary management strategies that can capitalise on enormous variety of rice production systems in very different climates. Sustaining rice production in adverse environmental conditions is a big challenge in the coming years. To overcome problems caused by biotic and abiotic stresses, new rice varieties with higher yield and improved tolerances need to be developed, taking advantage of all the resources and technologies currently available.
In this review, we present an overview of the major biotic stresses that affect rice production, with special emphasis on the rice blast disease. As pathogen infection causes major problems in rice cultivation, and nutrient stress (deficiency or excess) can be a factor in disease resistance, in this review we describe problems associated with inadequate supply of nutrients in resistance to pathogen, focusing on blast resistance. Our current knowledge on the molecular mechanisms of the plant immune system, including microRNA (miRNA)-mediated regulation of gene expression, is summarised. Lastly, we discuss the relevance of miRNAs in crosstalk between nutrient stress and immune signalling in plants, particularly in rice.
2. Biotic and Abiotic Stresses in Rice
Rice is prone to various types of stresses, both biotic and abiotic. Biotic stresses, caused by fungi, bacteria, viruses, insect pests, or nematodes, represent a serious threat to sustainable rice production. To reduce losses caused by pathogen infections and to obtain maximum yields, high rates of pesticides and fertilisers continue to be applied in rice farming. This has been harming to the environment and human health. Furthermore, climate change is promoting the northerly movement of pests, which means that temperate rice agriculture should be concerned about tropical pests and diseases.
Abiotic stresses, such as drought, salinity, cold, high temperature, submergence, nutrient deficiencies, and heavy metal toxicity are also factors negatively affecting rice growth and productivity worldwide. As most of the global rice supply originates from flooded ecosystems, rice is one of the most affected crops by drought stress. High salinity is also a major stress factor that can seriously affect rice growth, development, and yield. Indeed, rice is rated as the most salt-sensitive cereal crop. Nutrient stress can be also a major cause of decline in rice production. Nutrient stress refers to deficiencies of essential plant nutrients as well as excess of nutrients caused by over-fertilisation. Under nutrient stress conditions, the plant must respond dynamically to maintain nutrient levels in the optimal range for normal growth and development. Physiological disorders, also called physiological diseases, are quite common in rice grown under poor soil conditions [12].
It is also true that, in nature, plants are simultaneously exposed to varying combinations of biotic and abiotic stresses. However, most studies to determine the effects of environmental stress have been performed on plants exposed to an individual stress. So far, there is limited research focusing on the impact of combined abiotic and biotic stresses in rice. Crosstalk between plant responses induced by one stress may result in positive or negative impact over the other stress [13].
2.1. Biotic Stress
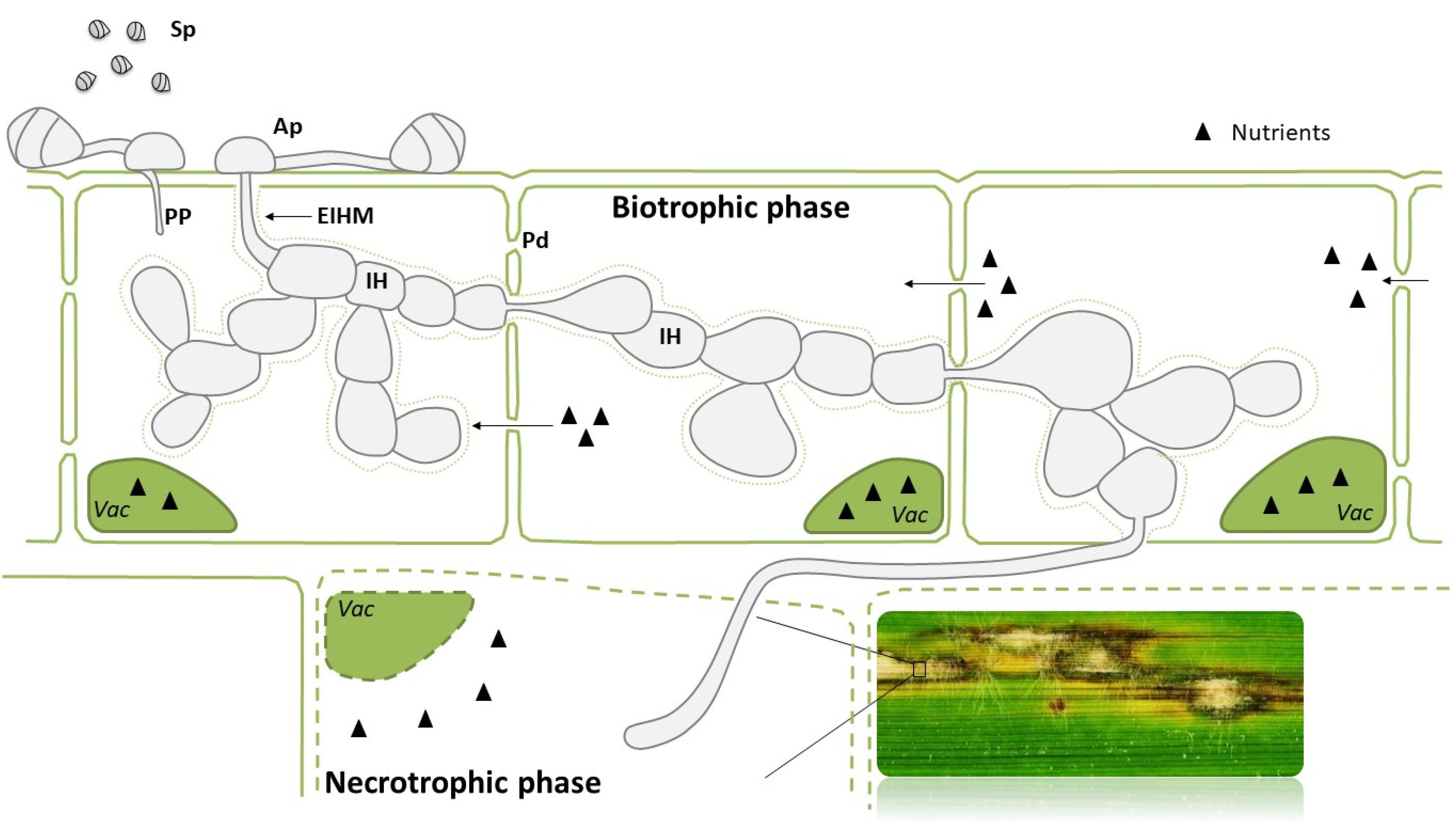
One of the most important pathogens affecting rice is the fungus Magnaporthe oryzae. This fungus can infect the leaves, leaf collars, necks, and panicles of rice plants and it causes rice blast. M. oryzae is an ascomycete fungus with a hemibiotrophic lifestyle that involves initial proliferation inside living host cells before switching to a destructive necrotrophic mode [14,15]. The blast disease cycle starts when a conidium reaches the host tissue and develops a germ tube, which grows on the leaf surface before differentiating into the dome-shaped appressorium (Figure 1). A penetration peg develops from the appressorium to enter into the epidermal cells, where it expands to become an infective hypha. During the biotrophic stage, the hyphae move from cell to cell through plasmodesmata [16]. At this stage of the infection process, the fungus redirects nutrients from living plant cells to facilitate nutrient acquisition from the host tissue (Figure 1) [15,17]. Several days after infection, the fungus switches to a necrotrophic lifestyle and blast lesions appear on the leaf surface, typically diamond-shaped with a grey or white centre and brown or reddish-brown border (Figure 1). The fungus sporulates profusely in these lesions, thus, facilitating the disease to spread rapidly to adjacent rice plants. Neck rot and panicle blast are particularly devastating diseases that cause major yield losses in severe epidemics. Furthermore, M. oryzae is capable of infecting different crops apart from rice, such as barley or wheat, which highlights its incredible ability to infect monocotyledonous plant species. The M. oryzae genome has been sequenced, and functional genomic approaches have led to the identification of hundreds of genes involved in its pathogenesis. M. oryzae has become a model fungus for studying host-pathogen interactions, at both cell biology and molecular levels [18].
Rice blast is a classical gene-for-gene interaction system, and numerous blast resistance (R) genes are identified in rice [19,20]. Furthermore, numerous avirulence (AVR) genes have been genetically and functionally described in M. oryzae [21]. However, traditional resistance conferred by R genes often breaks down in a few years due to the high variability and fast evolving fungal populations. Therefore, breeders usually prefer rice varieties that contain multiple R genes and have broad-spectrum blast resistance [22]. Chemical fungicides acting on the M. oryzae life cycle (e.g., inhibitors of melanin biosynthesis) are currently used for the control of the blast disease [23]. Chemical treatments aiming at the activation of plant defences, such as probenazole or spermidine, are also used for blast control [24,25]. The wide use of these chemicals might, however, provoke the development of resistance in the pathogen and/or cause side effects on non-target organisms and the environment. As there is a deep public concern about the side effects of chemical fungicides, it is critical to have effective methods to control the rice blast disease in an environmentally friendly way.
Bakanae (“foolish seedling” in Japanese) disease, caused by one or more seed-borne Fusarium species, mainly F. fujikuroi, is a disease of increasing economic importance in major rice-producing areas. The fungus infects rice plants from the seedling stage to the mature stage, with severe infection of rice seeds [28]. Symptoms of the disease are poor seedling emergence, abnormal elongation of internodes in the seedlings due to the ability of F. fujikuroi to produce gibberellic acids (GAs), and grain contamination. The most common management practices to limit bakanae are based on thermal seed treatment (hot water immersion) and application of fungicides. On the other hand, very few QTLs (Quantitative Trait Loci) for resistance to bakanae have been mapped in the rice genome, and only a small number of rice varieties showing resistance to the disease are identified [29]. Fusarium species also produce mycotoxins in rice grains that represent serious food safety concerns in animal and human health [30].
Another economically significant rice disease affecting rice worldwide is rice sheath blight, which is caused by the fungus Rhizoctonia solani. This fungus is a necrotrophic soil fungus that infects rice plants by vegetative mycelia and sclerotia that is able to survive for long periods in the soil. Its dispersal is facilitated by water management practices and infected plant debris. Protection of rice from sheath blight relies on the use of chemical fungicides and bacterial biocontrol agents (e.g., commercial formulations of Bacillus subtilis) [31].
Bacterial blight and bacterial leaf streak, caused by Xanthomonas oryzae pv. oryzae and X. oryzae pv. oryzicola, respectively, are considered as major bacterial diseases in rice [32]. X. oryzae enters into the host through wounds or natural openings on the surface of the plant. Once inside, this pathogen moves across the plant through the xylem and colonises different host tissues. Other bacterial pathogens of rice are Dickeya zeae that causes bacterial foot rot [33] and Burkholderia glumae that causes bacterial panicle blight of rice [34]. Fungicides are being used for the control of bacterial diseases in rice.
Rice is also vulnerable to a number of diseases caused by viruses that occur especially in the tropics [35]. The distribution of viruses affecting rice is generally restricted to one of the continents in which rice is grown, e.g., Rice hoja blanca virus (RHBH) in South America and Rice Yellow Mottle Virus (RYMV) in Africa. Rice stripe virus (RSV) and Rice tungro viruses (RTVB and RTSV) are found in Asia. Most of the rice viruses are transmitted by insect vectors in a persistent or non-persistent manner.
Insect pests are also important biological constraints limiting rice yield. Stem borers are generally considered the most damaging insect pest of rice. In particular, the striped stemborer (SSB) (Chilo suppressalis) is a major constrain on rice production. Stemborers are difficult to control with insecticides, as once they are inside the stem, they are protected from chemical sprays. In order to be successful with the chemical control, repeated applications of appropriate insecticide are needed. The insecticidal Cry proteins produced by Bacillus thuringiensis have provided a tool for the control of a wide diversity of insect pests, including the rice SSB, and rice varieties protected from SSB damage through the expression of Bt cry-endotoxin genes (Bt rice) have been developed [36]. The public controversy over genetically modified (GM) crops is, however, limiting the cultivation of Bt rice varieties. Conventional breeding for C. suppressalis resistance has proven to be difficult due to the poor knowledge of the genetic basis of this trait. It is also worthy to mention the problem with the herbivore Nilaparvata lugens, also known as the brown planthopper [37].
The root knot nematode Meloidogine graminicola is the most serious nematode pest of rice. This nematode is well-adapted to several scenarios where rice is grown, thus causing severe losses of this crop. Among the symptoms, it is possible to find stunted growth, yellowing, and gall formation on the roots of rice plants, symptoms that are greatly dependent on the environmental conditions [38].
Clearly, integrative strategies are required for successful control of rice diseases, including the use of advanced breeding lines and cultivars, forecasting distribution of the disease and pathogen races, examining wild Oryza species for novel sources of resistance, as well as using appropriated disease and water management systems. Modern biotechnology offers new opportunities for the development of rice varieties with enhanced disease resistance, which will be beneficial if rice yields are increased using sustainable strategies.
2.2. Plant Responses to Nutritional Stresses
Nutrients, macronutrients (nitrogen, phosphorus, potassium) and micronutrients (iron, copper, zinc, manganese, nickel, etc.) are taken from the soil and play a very important role in plant growth and development. Plants naturally evolved physiological mechanisms and adaptation strategies to cope with nutrient deprivation. Under limiting nutrient conditions, plants change the root system architecture by stimulating root branching and can also secrete small molecules into the soil to enhance the nutrient’s bioavailability. On a molecular level, nutrient limitation associates to the induction of genes encoding nutrient transporters to increase uptake, as well as genes involved in recycling or reallocation of nutrients to maintain nutrient levels in the optimal range for plant growth. Under nutrient excess, the plant responds through modification of nutrient acquisition programs, detoxification pathways, and sequestration into cellular compartments (e.g., vacuoles).
Nitrogen (N) is a component of important molecules, such as amino acids, proteins, or nucleic acids. The most available forms of N in the soil are nitrate and ammonium. Plants have developed different strategies to overcome low N in the soil, aiming to enhance N use efficiency through the regulation of N utilisation and acquisition. Alterations in root architecture and increased activity of influx transporters are the major determinants to increase N acquisition efficiency [39].
Phosphorus (P), like N, is a main macronutrient in plants. It is a component of nucleic acids and phospholipids, and also plays crucial roles in enzymatic reactions and signal transduction cascades (e.g., protein phosphorylation). P is an integral component of adenosine triphosphate (ATP), the universal cellular energy cofactor in biological processes. In plants, P is absorbed through the roots in the form of inorganic phosphate (Pi). Even though the overall content of Pi in the soil is generally high, its low bioavailability represents a limiting factor for plant growth in many agricultural ecosystems [40]. P deficiency is widespread in major rice ecosystems and is the major growth-limiting factor in acid upland soils where soil P-fixation capacity is often important. To explore the soil, the Pi-starved plants change their root architecture by increasing lateral roots and root hairs [41]. The most evident symptoms of Pi deficiency in plants are the reduction in plant growth and development of thinner leaves with darker appearance [42]. Pi-deprived plants experience important transcriptional reprogramming aiming to increase Pi acquisition in the root, translocation from the root to the shoot, and mobilisation of Pi stored in the vacuole, the so-called Phosphate Starvation Response (PSR) [43,44,45]. Another strategy developed by Pi-starved plants is an increase in RNAses and phosphatase activities, these enzymes being involved in metabolic Pi recycling [46,47]. Pi excess in the soil also has an effect on root development, as the plants show a reduction in primary root growth and less meristematic activity [48]. The excess of Pi also provokes alterations in the content of other nutrients such as iron or zinc [48]. Excessive Pi accumulation leads to toxicity at the tip of rice leaves (leaf tip necrosis), being more evident in older leaves.
Potassium (K) is another important inorganic nutrient for plant growth and production. It plays important roles in plant cell processes related with stomata closure, photosynthesis, turgor pressure, ion transport, or enzyme reactions as a cofactor. Although most soils are rich in K minerals, relatively little K+ is present in forms that are available to plants. When the plant senses the lack of this nutrient in the soil, it triggers the activation of channels and K transporters and, in some cases, is accompanied by the induction of other cellular processes which are also regulated by Ca2+, Reactive Oxygen Species (ROS), and hormones such as abscisic acid (ABA) [49].
Although required in smaller amounts than macronutrients (N, P, K), micronutrients are also essential for plant growth and play an important role in balanced crop nutrition. Iron (Fe), copper (Cu), zinc (Zn), manganese (Mn), molybdenum (Mo), and nickel (Ni) are metal micronutrients that function in a range of cell’s chemical reactions. Silicon (Si) and boron (B) mainly contribute to the plant’s structure, while the major role of the non-metal chlorine (Cl) serves in osmotic balance.
Fe performs a number of important functions in plant cellular processes, and like other elements, it functions as a cofactor of many enzymes required for a wide range of biological functions. Oxidation-reduction reactions are most commonly associated with Fe-containing enzymes, such as catalases and peroxidases. Fe is a component of ferredoxin, which is indispensable for photosynthesis, and cytochromes, which are necessary for the electron transport system in mitochondria and chloroplasts. Symptoms of Fe deficiency include pronounced interveinal chlorosis in which photosynthetic tissues turn yellow due to the lack of chlorophyll. Although Fe is generally present at high quantities in soils, its bioavailability is limited in aerobic conditions where iron is predominantly found in the Fe3+ form, mainly as a constituent of polymers with extremely low solubility [50]. Rice is, however, typically cultivated under anaerobic conditions (paddy fields) where abundant Fe2+ is readily available to the plant, especially in soils with a low pH in which Fe3+ is reduced to the more soluble ferrous ion Fe2+. Under such conditions, unlike other graminaceous plants, the absorption of Fe2+ by the rice roots might cause severe Fe toxicity. Excessive Fe accumulation in rice tissues causes overproduction of ROS, which, in turn, causes oxidative damage to macromolecules (nucleic acids, lipids, proteins, etc.) and damage to cellular structures [51]. Rice responds to Fe toxicity by means of different adaptive mechanisms. Firstly, rice can retain Fe in the root tissues and decrease Fe translocation from roots to shoots to avoid excessive Fe accumulation in leaves. Secondly, the Fe2+ ions, once in the cells, might associate with proteins (e.g., ferritins) or can be stored in cellular compartments (vacuoles). Under Fe excess, the rice plant activates the expression of genes associated with protection against oxidative stress [51]. High iron content in the soil also impairs the absorption of Cu in rice.
Cu serves primarily as a cofactor of metabolic enzymes and protein complexes in the electron transport chain. Cu is also an integral member of plastocyanin, which actively participates in the electron transport of chloroplast grana during photosynthesis. On the other hand, zinc is the cofactor for enzymes involved in N metabolism, and Zn deficiency declines anaerobic root metabolism and lessens the capacity of rice seedlings to cope with anaerobic soil conditions. Rice plants are more susceptible to Zn deficiency in early growth phases. On the other hand, B helps in the biosynthesis of cell walls, and thereby participates in cell division and elongation processes. B is critical for pollen tube elongation and B deficiency results in minimal pollen viability and lower grain yield [52].
Finally, rice is particularly susceptible to heavy metals contamination, in particular, cadmium (Cd) and arsenic (As) coming from contaminated soil, river, or groundwater. Upon exposure to heavy metals, plants experience oxidative stress that leads to cellular damage and disturbance of cellular ionic homeostasis. These toxic metals accumulate in rice grains and are significant risks for human and animal health. Water management practices significantly influence the Cd and As concentration in rice grains [53].
In the following sections, we present an introduction to the principles of plant immune responses. As nutrient stress can have an impact on resistance to pathogen infection in plants, we discuss interactions between nutrient responses and immunity in rice. Understanding how rice plants respond to pathogen infection in heterogeneous and fluctuating environments is crucial for the development of efficient means of disease control.
3. Plant Innate Immunity
Plants have evolved multiple mechanisms to sense and respond to pathogen infection. Resistance to pathogen infection relies on numerous changes at the biochemical, physiological, and molecular level, largely coordinated by a massive and fast reprogramming of the host transcriptome. Upon contact with the plant, conserved molecular signatures derived from microbes, known as pathogen-associated molecular patterns (PAMPs; or microbe-associated molecular patterns, MAMPs) are recognised by Pattern Recognition Receptors (PRRs) localised in the plasma membrane [13,54,55,56,57]. Examples of MAMPs include bacterial flagellin, lipopolysaccharide, EF-Tu, lipoproteins, peptidoglycans, and fungal chitin [58]. Plant PRRs fall into the receptor-like kinase (RLK) or receptor-like protein (RLP) families [59]. PAMP recognition triggers the activation of a general defence response referred to as PAMP-triggered immunity (PTI), in which multiple signal transduction pathways operate. Plants can also detect their own molecules such as oligogalacturonides released from plant cell walls as a consequence of physical damage caused by the pathogen infection, referred to as Damage-Associated Molecular Patterns (DAMPs), that are also able to initiate innate immune responses [58,60]. Extracellular ATP and endogenous elicitor peptides can also function as danger signals for the activation of the innate plant immune response [61]. Some pathogens are able to produce effector proteins that are delivered into the host cell to suppress PTI responses. Therefore, the pathogens carrying these effectors can cause diseases in their host plants [56]. During plant–pathogen coevolution, however, plants developed resistance (R) proteins, encoded by resistance (R) genes that recognise microbial effectors (or host proteins modified by effectors). This recognition triggers a rapid and robust host defence response, known as Effector-Triggered Immunity (ETI) [54,56]. Most R proteins belong to the nucleotide-binding site (NB)/leucine-rich-repeat (LRR) receptor family (NLR) [19]. ETI is often accompanied by a form of programmed cell death at the site of pathogen infection, the hypersensitive response (HR), which restricts the spread of infection. Accumulating evidence also supports that PTI and ETI share a number of signalling components [62]. Timing and intensity of the host responses, however, differ between PTI and ETI [63].
One of the early events in the plant response to pathogen infection is the increase in cytoplasmic Ca2+ for the activation of kinase cascades in which calcium-dependent protein kinases (CDPKs) and mitogen-activated protein kinases (MAPKs) participate [64]. Pathogen-induced defence responses include the production of ROS, such as hydrogen peroxide (H2O2), superoxide radicals (O2−), and hydroxyl radicals (−OH) [65]. ROS are reactive molecules with multiple functions in plant defence reactions to pathogen infection. In particular, H2O2 possess antimicrobial activity and contributes to cell wall reinforcement (e.g., lignification and oxidative cross-linking of cell-wall polymers) [66]. ROS also function as signalling molecules for the induction of defence responses. Another free radical that is produced in the plant during infection is nitric oxide (NO). NO seems to work together with ROS radicals in triggering cell death upon pathogen challenge and is also strongly related with Ca2+ and ABA signalling for regulation of the stomata closure [67].
Plants also produce proteins and secondary metabolites that can be toxic to phytopathogens. Perhaps the best-known examples of defensive metabolites are phytoalexins, which are synthesised de novo in response to infection. Major phytoalexins accumulating in rice are the diterpene phytoalexins momilactones, phytocassenes, and oryzalexins, and the flavanone phytoalexin sakuranetin [68,69]. However, Brassicaceae plant species accumulate camalexin, a tryptophan-derived phytoalexin with antifungal and antibacterial activity against phytopathogens [70]. Indole glucosinolates are also effective to limit colonisation by necrotrophic fungi in plants [71]. The accumulation of Pathogenesis-Related (PR) proteins is also a ubiquitous response of plants to pathogen infection, some of these proteins exhibiting antimicrobial activity [72].
Hormones play a crucial role in disease resistance by regulating plant immune responses [73]. Immune responses are coordinated mainly by the phytohormones salicylic acid (SA), ethylene (ET), jasmonic acid (JA and JA-derivatives), as well as by auxins and brassinosteroids. Plant hormones do not function independently, as synergistic and antagonistic interactions between hormone signalling pathways ultimately drive the fine-tuning of plant defence responses. Generally, the JA signalling pathway synergistically cross-talks with the ET signalling pathway, whereas the SA and JA pathways show negative crosstalk [73]. Hormone signalling networks in plant defence appear to be dependent on both the lifestyle of the pathogen in the host plant and the host plant itself. Whereas SA has been generally associated with plant defence against biotrophic pathogens, JA and ET are associated with protection against necrotrophic pathogens.
Historically, plant immune responses to fungal and bacterial pathogens have been considered to rely on the function of protein-coding genes, whereas RNA-based mechanisms largely regulate plant–virus interactions. Along with this, the transcriptional control of protein-coding defence genes has been studied extensively, but less attention has been paid to the post-transcriptional regulation of defence-related processes. Nowadays, the contribution of host endogenous small RNAs in the regulation of plant defence responses to infection by fungal and bacterial pathogens is well demonstrated [74,75,76,77]. Accordingly, the involvement of microRNAs (miRNAs), a class of small RNAs from plants, in post-transcriptional regulation of gene expression in the rice immune system is discussed below, with special emphasis on mechanisms involved in the rice response to infection by the blast fungus M. oryzae.
3.1. Regulatory Role of microRNAs in Rice Immunity
MiRNAs are short non-coding small RNAs that direct post-transcriptional gene silencing through cleavage or translational repression mechanisms [78,79]. In addition to miRNAs, plants possess another class of small RNAs, the small interfering RNAs (siRNAs) [80,81]. The two classes of small RNAs, miRNAs and siRNAs, are distinguished by their mode of biogenesis, function, and mechanisms of action. MiRNAs derive from long single-stranded RNA precursors with imperfect stem-loop structure (primary miRNAs, or pri-miRNAs) which are processed in a two-step process by RNAse III DICER-like proteins (DCL, typically DCL1) to produce a precursor miRNA (pre-miRNA) and then a double-stranded duplex, the miRNA-5p/miRNA-3p. The functional strand of the duplex is incorporated into an ARGONAUTE (AGO) protein in the RNA-induced silencing complex (RISC) where the miRNA recognises target transcripts by sequence complementarity [80]. Contrary to miRNAs, siRNAs derive from perfectly paired, long double-stranded RNA precursors that are produced by the activity of an RNA-dependent RNA polymerase (RDR). Readers interested in the details on miRNA biogenesis and mode of action are referred to specialised reviews [81,82,83].
According to their nucleotide sequence, miRNAs are categorised into different families, which comprise one or more members. The miRBase is a repository of miRNA sequences and annotations (www.mirbase.org (accessed on 3 June 2021) [84]. Currently, the miRBase contains 738 mature rice (O. sativa) miRNAs, following Medicago truncatula and Glycine max with 756 miRNAs in both species (miRBase release 22.1).
Plant miRNAs play a critical role in controlling diverse developmental processes and adaptation to environmental stress, both biotic and abiotic stresses [74,75,85,86,87,88]. Certain miRNAs have been shown to control traits of agronomic importance in rice, such as tillering, flowering, panicle and grain production [89,90]. Examples of miRNAs involved in tolerance to abiotic stress in rice are miR398 (drought, salt, and oxidative stresses), miR319 (cold stress), miR393 (salinity), or miR399 (nutrient homeostasis) [85,91].
The role of miRNAs in disease resistance was described for the first time in the model plant Arabidopsis thaliana [92]. Here, perception of the Pseudomonas syringae flagellin peptide flg2 induces miR393 accumulation, and down regulation of auxin receptors, which in turn results in bacterial resistance. Since then, other miRNAs have been reported to play a key role in Arabidopsis immunity, such as miR160, miR167, miR393, miR396, miR398b, miR400, miR472, miR773, miR844, miR858, and miR1916, among others [75,87,93,94,95,96]. Increasing evidence supports that plant miRNAs play a role in PTI and ETI. Depending on the target gene that they regulate, miRNAs might function as positive or negative regulators of immune responses [97]. Most of our current understanding of miRNAs in plant immunity comes from studies on the interaction of Arabidopsis plants with the bacterial pathogen P. syringae. Limited information is available on miRNAs involved in disease resistance in other plant species and/or during interaction with other types of pathogens.
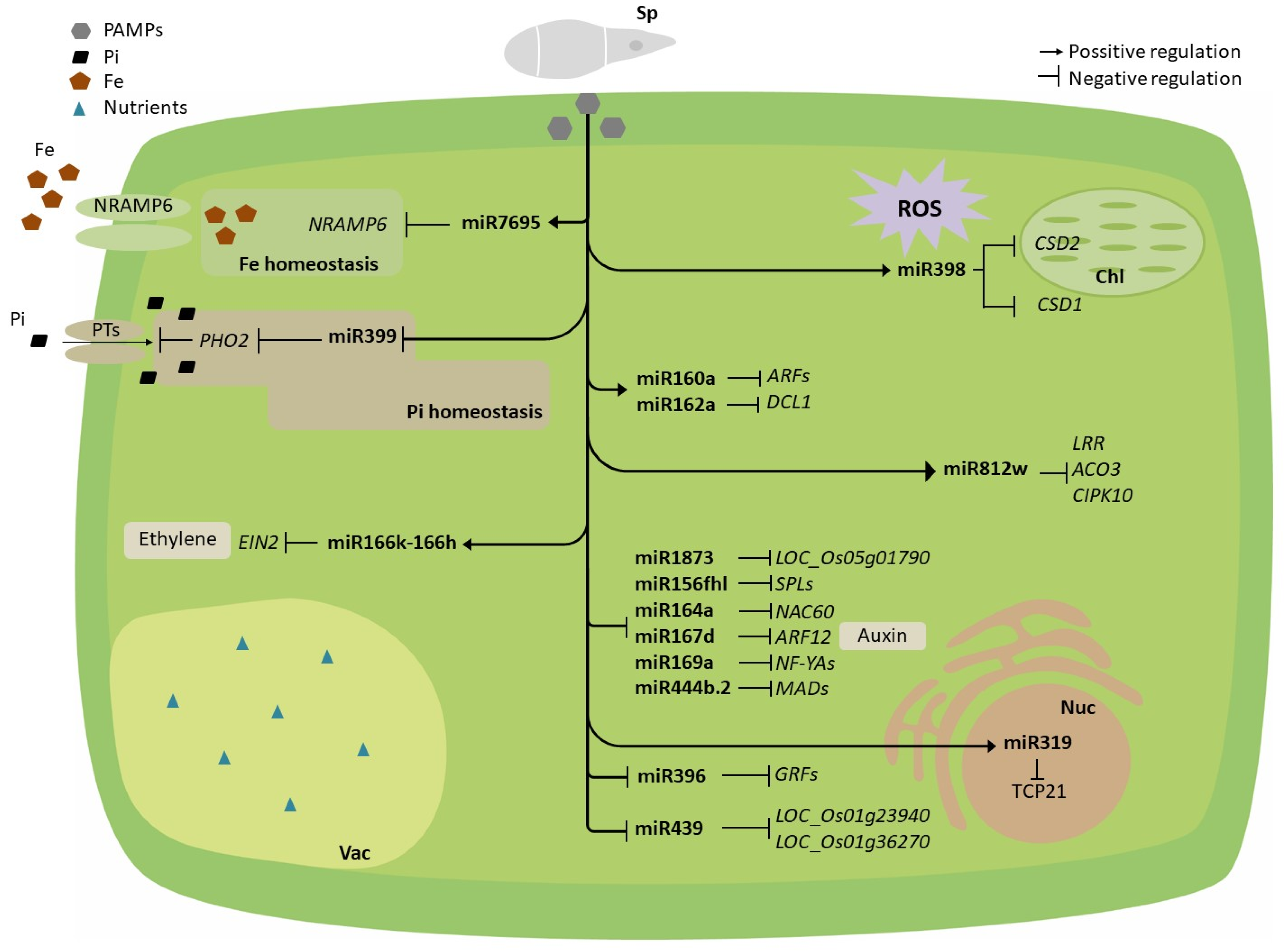
Even though a number of miRNAs have been shown to respond to pathogen infection in rice, the biological role of these pathogen-regulated miRNAs in rice immunity is not completely understood. Indeed, the biological role of only few miRNAs has been characterised in the rice/M. oryzae interaction (Figure 2). Some of these miRNAs function as positive regulators of immune responses (miR7695, miR160a, miR162a, miR398b, miR166k-166h and miR812w), whereas other miRNAs function as negative regulators of rice immune responses (miR156fhl, miR164a, miR167d, miR169a, miR319b, miR396, miR399, miR444b.2, miR439, and miR1873) [77,98,99,100,101,102,103,104,105,106,107,108,109]. To note, miR398 has been reported to function as a positive regulator of immune responses in rice against M. oryzae while negatively regulating immune responses against P. syringae in Arabidopsis [110].
In other studies, the activation of MIR166k-166h encoding a polycistronic miRNA from rice was found to enhance resistance to infection by M. oryzae and F. fujikuroi in rice [103]. The regulatory role of miR166k-166h relies on the activity of miR166k-5p (encoded by the miR166k-166h precursor) on EIN2 (ETHYLENE INSENSITIVE 2) transcripts, the EIN2 gene being a component of the ethylene signalling pathway in rice plants. Ethylene signalling is required for resistance against the blast fungus M. oryzae in rice [111].
Regarding miR812, a new member of the miR812 family in rice, miR812w, involved in blast resistance was recently described [99]. Whereas overexpression of miR812w increased blast resistance, CRISPR/Cs9-mediated MIR812w editing enhances disease susceptibility. Of interest, miR812 originated from a miniature-inverted repeat element (MITE) of the Stowaway family of DNA transposons from rice, and the incorporation of cognate MITEs into protein-coding genes would have created target sites for miR812w. Moreover, miR812w directs methylation in trans at its target genes, as well as in cis at the MIR812w locus [99].
Not only miRNAs themselves, but also components of the miRNA biogenesis machinery appear to play a role in blast resistance. Whereas silencing of OsDCL1 enhances resistance to the rice blast fungus, its activation compromises resistance against this fungus [112,113]. OsDCL1 activation represses pathogen-inducible rice defence responses, impairs diterpenoid phytoalexin production, and provokes important alterations in the population of rice miRNAs [113].
The functional characterisation of miRNAs, and components of the miRNA biogenesis and function machinery involved in disease resistance, also opens the possibility of using miRNAs as biomarkers in the management of disease resistance in rice breeding programs.
3.2. Impact of Nutrient Stress on Rice Immunity
Nutrient stress caused by excess or deficiency of nutrients might affect the plant’s response to pathogen infection, thus, determining the outcome of the interaction, resistance or susceptibility. The impact of nutrient stress on disease resistance is, however, difficult to predict, as different results are observed in the outcome of the interaction (resistance, susceptibility) depending on the identity of the interacting partners (host and pathogen) [114]. Although adaptation to nutrient stress and immunity are not independent processes, the molecular mechanisms involved in nutrient responses and innate immunity have been so far investigated separately from each other. To better understand how rice plants defend themselves during pathogen attack, it is essential to consider that they can be exposed to adverse environmental conditions, including improper nutrient supply and nutritional imbalances. Being a foliar pathogen, the fungus M. oryzae must acquire nutrients from the host, hence, the availability of host nutrients can be an important factor contributing to the establishment of the rice blast disease [17].
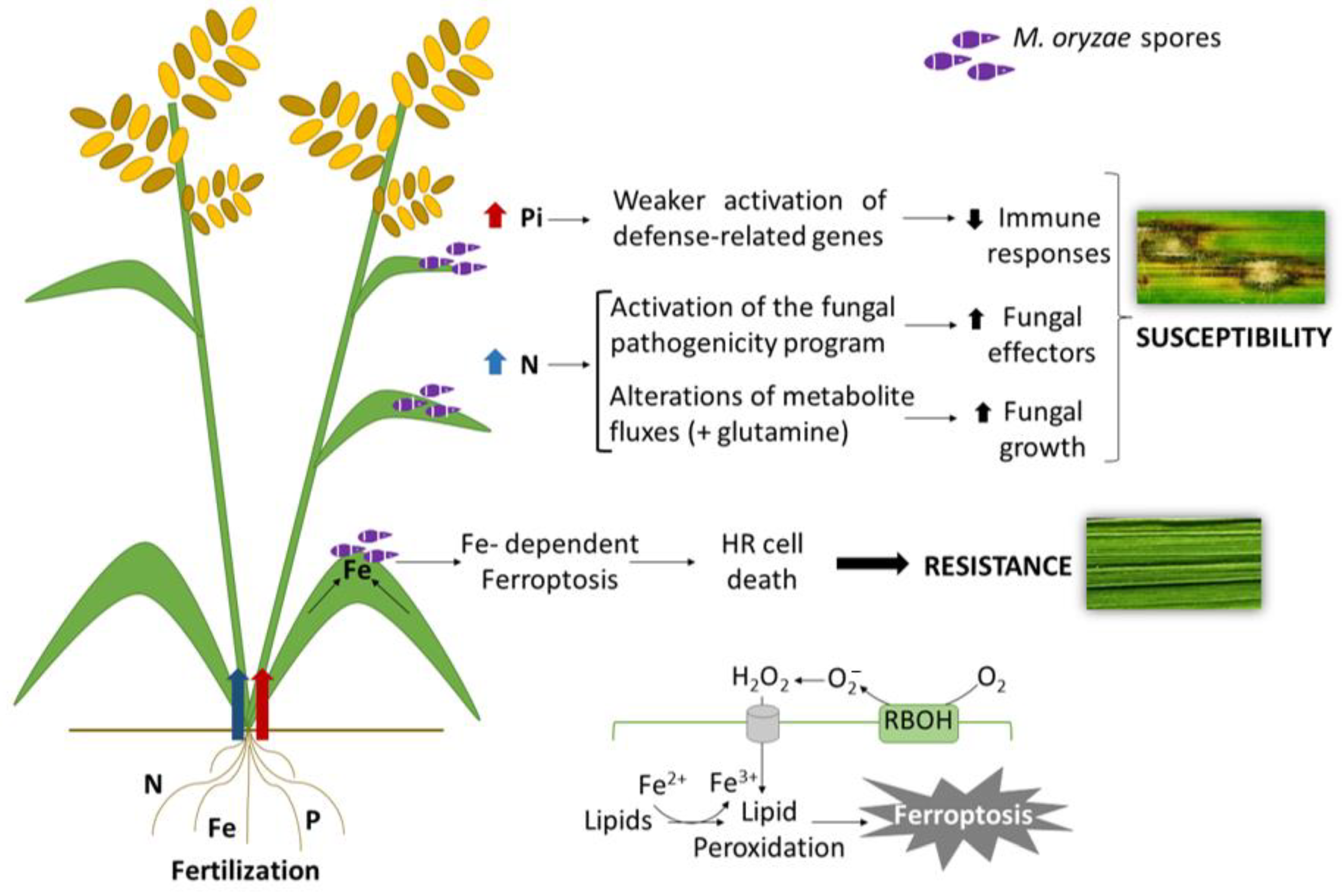
Plants can develop different nutritional-based defence mechanisms based on sequestration of essential nutrients away from the invading pathogen (withholding strategies), or localised accumulation of nutrients at the infection site that become toxic to the pathogen (i.e., hyper-accumulation of Fe for activation of an oxidative burst). The term “nutritional immunity” is used in human health as a non-specific host immune response and refers to mechanisms for withholding iron availability by the host that becomes inaccessible to the pathogen [115]. The term nutritional immunity can be also applied to restriction mechanisms of essential nutrients in plants to defend against pathogens. In plant/pathogen interactions, the host and the pathogen compete with each other for essential nutritional resources. This dynamic interaction is particularly relevant when plants are grown under nutrient limiting conditions. On the other hand, an increase in nutrient supply might create a more favourable environment for pathogen growth that would allow the pathogen to improve nutrient acquisition from the host plant, thus, promoting pathogenicity. However, the massive use of fertilisers might also have a negative impact on nutrient use efficiency in the plant and might be responsible for metabolic changes that jeopardise the plant defence responses. On the contrary, the accumulation of certain nutrients that can be toxic for the pathogen (e.g., iron) might protect the plant from infection. Figure 3 illustrates how interactions between nutrient and immune signalling pathways, particularly N, Pi, and Fe signalling, might lead to increased resistance or susceptibility to M. oryzae infection in rice.
It is generally assumed that N nutrition has an effect on disease incidence in different plant species [116]. During infection, pathogens require N sources, including NH4+ and NO3−, as well as amino acids. It has long been recognised that high N fertilisation enhances susceptibility to the blast fungus in rice plants [117]. It is also known that N supply has an impact on the two interacting partners: host and pathogen (Figure 3). From the perspective of the host plant, N availability has an effect on plant primary and secondary metabolism which, in turn, might affect host defence responses. While promoting plant growth, high N input might result in less formation of lignin and reduction in the thickness of the secondary cell wall, the plant’s physical barrier to pathogen infection [118]. From the perspective of the pathogen, fungal pathogenicity can be affected by N availability. The nitrogen-induced susceptibility (NIS) to rice blast has been shown to be associated with the induction of rice genes implicated in N recycling and an increase in M. oryzae pathogenicity [119]. Contrary to what is observed in rice, N fertilisation reduces disease severity caused by Verticillium spp. in Solanum species, indicating that no generic model can describe the role of N in a given interaction [114].
Regarding Pi nutrition, emerging evidence supports the existence of crosstalk between the Pi starvation signalling machinery and immune responses in plants [121,122]. However, our knowledge of interactions between plant adaptive mechanisms to Pi excess and immunity is still limited. Only recently, it was described that high Pi fertilisation compromises the expression of immune responses and enhances susceptibility to infection by M. oryzae in rice plants [98] (Figure 3). In other studies, overexpression of the OsPT8 phosphate transporter, and subsequent increase in Pi content, was reported to enhance susceptibility to M. oryzae infection [123]. Therefore, over-use of fertilisers in rice fields might have unintended consequences in rice production by facilitating blast infection, while contributing to water and soil pollution.
Rice plants are also able to establish symbiotic associations with arbuscular mycorrhizal (AM) fungi [124]. Root colonisation by AM fungi improves the uptake of mineral nutrients in the host plant, mainly P and N [125,126,127,128]. Even though root colonisation by AM fungi is reduced in rice plants grown under permanent flooded conditions, evidence supports that, once the AM symbiosis is established, its functional capacities are not affected by flooding [129]. During colonisation by AM fungi, the rice root system architecture undergoes drastic morphological and transcriptional alterations to accommodate fungal structures [130,131]. AM colonisation is, however, unevenly distributed within the rice root system: the crown roots are weakly colonised, the large lateral roots are strongly colonised, and fine lateral roots are not colonised [132,133].
Moreover, there is an important genetic variation within rice varieties for root colonisation by AM fungi [133]. Beneficial effects of root colonisation by AM fungi include growth stimulation, increased yield, and blast resistance, but differences are observed among cultivars in terms of blast resistance [134,135,136,137]. Recent studies in leaves of mycorrhizal rice plants revealed that the AM symbiosis is accompanied by a coordinated systemic regulation in the expression of genes involved in hormone signalling (JA, ET) and lipid signalling [136]. Then, systemic transcriptional responses occurring in leaves of mycorrhizal rice plants might be potentially important for defence against foliar pathogens. The AM symbiosis might as well contribute to the sustainability of rice production, an aspect that remains poorly explored.
As previously mentioned, Fe is an essential element for plants and a co-factor for a variety of proteins involved in redox reactions. Plants have developed strategies to mobilise Fe from the rhizosphere, referred to as Strategy I and Strategy II [138]. Typically, dicotyledonous species (such as Arabidopsis) and non-graminaceous monocotyledonous species use the strategy I, in which protons are released into the rhizosphere to acidify the soil and to increase ferric iron (Fe3+) solubility. Iron is then reduced to ferrous form (Fe2+) by a ferric reductase oxidase (FRO) and enters into the root cells by the activity of the Fe2+ transporters (IRTs). A different strategy, Strategy II, is used by graminaceous monocotyledonous plants, in which phytosiderophores are released into the rhizosphere and then reabsorption of the metal-phytosiderophore complex occurs by root membrane transporter proteins. Rice employs both strategies for iron uptake [138,139]. When in excess, however, iron becomes toxic to the plant cells due to the formation of ROS, which might cause oxidative damage to macromolecules and cellular structures, eventually leading to death. Therefore, iron homeostasis needs to be tightly controlled in the plant to avoid toxicity.
In plant–pathogen interactions, iron homeostasis must be carefully regulated to allow normal plant growth while providing a way to arrest pathogen growth. The molecular mechanisms underlying Fe homeostasis in the host plant during pathogen infection are, however, poorly understood. Evidence supports a relationship between iron content and expression of immune responses in plants, including rice [108,140,141,142]. In M. oryzae-infected rice leaves, Fe has been found to accumulate at the sites of attempted penetration by the fungus (appresoria), and in cells in the vicinity of the infection site, further supporting that Fe accumulation and redistribution in the host tissue might be important for blast resistance [108]. Iron- and ROS-dependent signalling cascades are known to be involved in the ferroptotic cell death pathway, a form of non-apoptotic, iron-dependent cell death that rice plants use to prevent M. oryzae infection [120] (Figure 3). Studies in wheat plants revealed that infection with Blumeria graminis f. sp. tritici elicits Fe accumulation in the apoplast, which in turn mediates apoplastic oxidative burst and induction of defence gene expression [143]. It has been proposed that disturbance of Fe homeostasis is sufficient to prime the plant immune system for enhanced defence [144].
Cu is an important element in a number of pesticides in agriculture because it interferes with numerous enzymatic reactions and blocks respiratory activity in bacterial, fungal, and oomycete pathogens. Accordingly, Cu is widely used in crop protection because of its antimicrobial activity against phytopathogens. In rice, Cu application is very effective to suppress bacterial blight, caused by Xanthomonas oryzae pv oryzae (Xoo). However, several bacterial pathogens have developed resistance to the metal, and Cu-resistant pathogenic bacteria have been reported because of the wide application of Cu-containing pesticides in agriculture worldwide [145].
In summary, evidence gathered over years indicates that nutrient stress has an impact on disease resistance in rice. Our understanding of the molecular mechanisms underlying the innate ability of rice plants to cope with pathogen infection in heterogeneous environments is, however, far from complete. Further investigation on these processes in different rice/pathogen/environment interactions would provide a solid foundation for the development of novel strategies for rationally optimising fertiliser and pesticide use in rice production and protection against diseases.
3.3. miRNAs in the Crosstalk between Nutrient Stress and Immune Signalling
Understanding how plants cope with pathogens under nutrient stress conditions and how nutrient signalling pathways interact with immune signalling is an issue of great importance in crop protection. Clearly, a dynamic regulation of gene expression is required for optimal plant responses to nutrient conditions that are diverse in time and space, as well as during pathogen infection. Being versatile regulators of gene expression, miRNAs are well suited for proper integration of these inputs.
Several miRNAs have been shown to play a key role in regulating nutrient homeostasis in plants [146,147]. Some of them are specifically responsive to a specific nutrient condition, but others can be regulated by different nutrient conditions. For instance, miR167, miR169, miR393, and miR827 have been described as nitrogen-responsive miRNAs, whereas miR395 (targeting ATP sulfurylase genes) functions in sulphur assimilation [146,147]. Nutrient-responsive miRNAs might then play a role in controlling nutrient homeostasis as well as crosstalk among nutrient stress responses. So far, most research on miRNAs involved in nutrient homeostasis focused on plants under nutrient-limiting conditions, and less is known about their implication in plants that encounter excess of nutrients in the environment. It is also true that, although a large number of miRNAs have been shown to be responsive to nutrient stress, how such alterations affect nutrient homeostasis remains to be determined.
The implication of miR399 in the phosphate starvation response of plants is well documented [43,91,148,149,150]. Most of our knowledge on this issue comes from studies in the model plant Arabidopsis thaliana, where Pi starvation is accompanied by an increase in miR399 accumulation in roots which, in turn, down-regulates the expression of PHO2 (PHOSPHATE 2). PHO2 encodes an ubiquitin E2 conjugating enzyme that mediates degradation of the Pi transporters. In this way, miR399 accumulation relieves negative post-transcriptional control of Pi transporters for an increase in Pi uptake. Moreover, miR399 moves from roots to shoots and serves as a signalling molecule in regulating Pi homeostasis during Pi deprivation at the whole-plant level. This allows the plant to increase uptake and root-to-shoot allocation of Pi in Arabidopsis plants under Pi-limiting conditions [91,148,151]. The miR399/PHO2 module has a conserved function in controlling phosphate homeostasis in Arabidopsis and rice plants.
Another miRNA controlling Pi homeostasis is miR827, which appears to function via different pathways in Arabidopsis and rice plants. In Arabidopsis, miR827 targets NITROGEN LIMITATION ADAPTATION (NLA) encoding an ubiquitin E3 ligase that mediates degradation of the plasma membrane-localised Pi transporter PHT1 [152,153], whereas in rice, miR827 targets two different OsSPX-MFS genes (OsSPX-MFS1 and OsSPX-MFS2) that localise at the tonoplast [154,155]. It has been proposed that in rice, miR827 functions in Pi compartmentalisation and Pi storage [154,155,156,157]. For additional information on Pi starvation responses in rice, we refer to [43,44,158].
Recent studies have demonstrated that the miR399 might function as a potential integrator of Pi starvation responses and immune-defence responses in rice plants. Thus, transgenic rice plants overexpressing miR399 showed increased phosphate content, weaker induction of defence-related genes, and enhanced susceptibility to M. oryzae, which also correlates with the susceptibility phenotype observed in rice plants grown in Pi excess conditions [98]. Other components of the Pi starvation signalling also regulate the expression of genes involved in immune responses [121,122].
On the other hand, miR398 functions in plant stress responses by regulating the expression of genes encoding copper superoxide dismutase (CSD1, CSD2) and Cu chaperones for SOD1 involved in protection against oxidative stress in plants [159,160]. Overexpression in rice of miR398 results in up-regulation of defence-related genes, which in turn enhances resistance to infection by M. oryzae [110].
In other studies, a rice miRNA, miR7695, has been reported to regulate iron homeostasis and rice immunity. MiR7695 negatively regulates the expression of OsNramp6 (Natural resistance-associated macrophage protein 6), encoding an iron and manganese transporter from rice [102,140]. miR7695 regulates not only Fe content, but also defence responses during infection of rice plants with M. oryzae. Both MIR7695 activation and growing rice plants under high Fe supply is accompanied by a superinduction of PR and diterpenoid phytoalexin biosynthesis genes which, in turn, results in enhanced resistance to infection by M. oryzae [108].
Thus, from what we have learned so far, distinct miRNAs can play a role in the crosstalk between nutrient stress signalling and immune signalling. MiRNAs might potentially work as molecular switches between these two intimately connected processes. As miRNAs are responsible for fine regulation of gene expression, rather than on-off regulations, these riboregulators are well suited to function in the crosstalk between stress-induced signalling pathways. At present, however, our knowledge on interconnected regulations between nutrient signalling and immune signalling, in which miRNAs participate, is still in its infancy.
4. Conclusions and Perspectives
With the predicted global demand for rice, in an ever-growing population, there is an increasing pressure to develop rice cultivars with improved stress responses and increased yield. The potential impact of climate change on rice production and the emergence of new diseases in a changing climate are also major issues that need to be addressed to ensure global food security. Towards this end, understanding the molecular mechanisms underlying disease resistance under nutrient stress conditions is essential for the development of effective strategies aiming at combating pathogen infection, an aspect that remains poorly explored. Fertilisers containing plant nutrients are commonly used to maintain optimal yield in rice cultivation. Ironically, excess nitrogen and phosphate fertilisation has been shown to increase susceptibility to infection with the blast fungus M. oryzae in rice [98,117]. Thus, the indiscriminate use of fertilisers might have adverse effects on the rice plant by increasing the likelihood of blast disease. There is the need to develop eco-friendly technologies to maintain rice productivity while reducing inputs of agrochemicals, fertilisers and pesticides, also in response to legislative and public pressure.
Regarding miRNA function, this is a revolutionary field of research, expanding in recent years. Distinct miRNAs are known to function in crosstalk between nutrient stress and immune responses. However, to fully exploit the potential of miRNAs as regulators of rice immune responses, a better knowledge of interconnected regulations between nutrient homeostasis and immune responses is needed. Here, it is worth mentioning that because of the small size of MIRNA genes, the functional analysis of MIR genes has historically been a challenge (e.g., lack of T-DNA insertional knock-out mutants). Alternative approaches have been used to repress endogenous miRNA activity which includes miRNA target mimicry, transcriptional gene silencing of miRNA promoters, and artificial miRNA-directed silencing of miRNA precursors. New technologies for precise, efficient gene targeting or genome editing such as the CRISPR/Cas9 system has been developed for genome editing and targeted mutagenesis of plant miRNAs, which have been successfully used for the functional analysis of rice miRNAs [99,161,162]. Using this methodology will enable us to expand our knowledge on the regulatory role of miRNAs in adaptive processes to environmental stress in rice plants, including pathogen resistance under nutrient stress conditions. Unravelling miRNA-mediated mechanisms governing pathogen resistance and nutrient homeostasis will be useful in designing novel strategies to improve not only disease resistance but also the nutritional status of rice plants with exclusion of use and release of agrochemicals. Since diseases caused by pathogens are major threats to rice production, high rates of pesticides and fertilisers are currently applied in rice production. As a consequence, environmental problems have arisen in rice growing areas due to excessive use of pesticides and fertilisers. The combination of novel molecular and biotechnological tools, screening technologies, advanced breeding, and cultural practices, also considering the plant microbiome, should become the main goal in agriculture in the 21st century.
Author Contributions
All authors contributed to and approved the submitted version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by FEDER/Ministerio de Ciencia, Innovación y Universidades—Agencia Estatal de Investigación (RTI2018-101275-B-I00). We acknowledge financial support from the Spanish Ministry of Science and Innovation—State Research Agency (AEI), through the “Severo Ochoa Programme for Centres of Excellence in R&D” CEX2019-000902-S, and the CERCA Programme from the “Generalitat de Catalunya”. B.V.-T. is a recipient of a Ph.D. grant from the Ministerio de Economia, Industria y Competitividad/Agencia Estatal de Investigación/Fondo Social Europeo (BES-2016-076289).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Mani Deepika Mallavarapu, Ferrán Sánchez-Sanuy, Héctor Martín-Cardoso, María Ribaya, Glòria Escolà, and Roberto Mateluna for critical reading of this work. We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAOSTAT. Crop Prospects and Food Situation in Rome: Food and Agriculture Organization of the United Nation. Available online: http://www.fao.org/faostat (accessed on 3 June 2021).
- Gutaker, R.M.; Groen, S.C.; Bellis, E.S.; Choi, J.Y.; Pires, I.S.; Bocinsky, R.K.; Slayton, E.R.; Wilkins, O.; Castillo, C.C.; Negrão, S.; et al. Genomic history and ecology of the greographic spread of rice. Nat. Plants 2020, 6, 492–502. [Google Scholar] [CrossRef]
- Sweeney, M.; McCoch, S. The complex history of the domestication of rice. Ann. Bot. 2007, 100, 951–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, B.L.; Zhao, Z. Archaeological and genetic insights into the origins of domesticated rice. Proc. Natl. Acad. Sci. USA 2014, 111, 6190–6197. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Yu, Y.; Haberer, G.; Marri, P.R.; Fan, C.; Goicoechea, J.L.; Zuccolo, A.; Song, X.; Kudrna, D.; Ammiraju, J.S.S.; et al. The genome sequence of african rice (Oryza glaberrima) and evidence for independent domestication. Nat. Genet. 2014, 46, 982–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linares, O.F. African Rice (Oryza glaberrima): History and future potential. Proc. Natl. Acad. Sci. USA 2002, 99, 16360–16365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londo, J.P.; Chiang, Y.C.; Hung, K.H.; Chiang, T.Y.; Schaal, B.A. Phylogeography of asian wild rice, Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. Proc. Natl. Acad. Sci. USA 2006, 103, 9578–9583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goff, S.A.; Ricke, D.; Lan, T.H.; Presting, G.; Wang, R.; Dunn, M.; Glazebrook, J.; Sessions, A.; Oeller, P.; Varma, H.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica). Science 2002, 296, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.S.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef]
- Li, Z.; Fu, B.Y.; Gao, Y.M.; Wang, W.S.; Xu, J.L.; Zhang, F.; Zhao, X.Q.; Zheng, T.Q.; Zhou, Y.L.; Zhang, G.; et al. The 3000 rice genomes project. Gigascience 2014, 3, 7. [Google Scholar]
- Mukkram Ali Tahir, R.; Noor-us-Sabah; Afzal, M.; Sarwar, G.; Rasool Noorka, I. Smart nutrition management of rice crop under climate change environment. In Protecting Rice Grains in the Post-Genomic Era; Jia, Y., Ed.; IntechOpen: London, UK, 2019. [Google Scholar]
- Saijo, Y.; Loo, E.P. Plant immunity in signal integration between biotic and abiotic stress responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.A.; Talbot, N.J. Under Pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.; Orth, K. Rise of a Cereal Killer: The biology of Magnaporthe oryzae biotrophic growth. Trends Microbiol. 2018, 26, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Kankanala, P.; Czymmek, K.; Valent, B. Roles for rice membrane dynamics and plasmodesmata during biotrophic invasion by the blast fungus. Plant Cell 2007, 19, 706–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.A.; Fernandez, J.; Quispe, C.F.; Gradnigo, J.; Seng, A.; Moriyama, E.; Wright, J.D. Towards defining nutrient conditions encountered by the rice blast fungus during host infection. PLoS ONE 2012, 7, e47392. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, F.; Nishimura, M.T. Structural, functional, and genomic diversity of plant NLR proteins: An evolved resource for rational engineering of plant immunity. Annu. Rev. Phytopathol. 2018, 56, 243–267. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Chern, M.; Yin, J.; Wang, J.; Chen, X. Recent advances in broad-spectrum resistance to the rice blast disease. Curr. Opin. Plant Biol. 2019, 50, 114–120. [Google Scholar] [CrossRef]
- Wang, B.H.; Ebbole, D.J.; Wang, Z.H. The arms race between Magnaporthe oryzae and rice: Diversity and interaction of Avr and R genes. J. Integr. Agric. 2017, 16, 2746–2760. [Google Scholar] [CrossRef]
- Wu, Y.; Xiao, N.; Chen, Y.; Yu, L.; Pan, C.; Li, Y.; Zhang, X.; Huang, N.; Ji, H.; Dai, Z.; et al. Comprehensive evaluation of resistance effects of pyramiding lines with different broad-spectrum resistance genes against Magnaporthe oryzae in rice (Oryza sativa L.). Rice 2019, 12, 11. [Google Scholar] [CrossRef]
- Takagaki, M. Melanin biosynthesis inhibitors. In Fungicide Resistance in Plant Pathogens; Ishii, H., Hollomon, D., Eds.; Springer: Tokyo, Japan, 2015; pp. 175–180. [Google Scholar]
- Iwai, T.; Seo, S.; Mitsuhara, I.; Ohashi, Y. Probenazole-induced accumulation of salicylic acid confers resistance to Magnaporthe grisea in adult rice plants. Plant Cell Physiol. 2007, 48, 915–924. [Google Scholar] [CrossRef] [Green Version]
- Moselhy, S.S.; Asami, T.; Abualnaja, K.O.; Al-Malki, A.L.; Yamano, H.; Akiyama, T.; Wada, R.; Yamagishi, T.; Hikosaka, M.; Iwakawa, J.; et al. Spermidine, a polyamine, confers resistance to rice blast. J. Pestic. Sci. 2016, 41, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Sakulkoo, W.; Osés-Ruiz, M.; Garcia, E.O.; Soanes, D.M.; Littlejohn, G.R.; Hacker, C.; Correia, A.; Valent, B.; Talbot, N.J. A Single fungal MAP kinase controls plant cell-to-cell invasion by the rice blast fungus. Science 2018, 359, 1399–1403. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, S.; Minami, E.; Nishizawa, Y. Live-cell imaging of rice cytological changes reveals the importance of host vacuole maintenance for biotrophic invasion by blast fungus, Magnaporthe oryzae. Microbiol. Open 2015, 4, 952–966. [Google Scholar] [CrossRef]
- Iqbal, M.; Javed, N.; Sahi, S.T.; Mehmood Cheema, N. Genetic management of bakanae disease of rice and evaluation of various fungicides against Fusarium moniliforme in vitro. J. Phytopathol. 2011, 23, 103–107. [Google Scholar]
- Volante, A.; Tondelli, A.; Aragona, M.; Valente, M.T.; Biselli, C.; Desiderio, F.; Bagnaresi, P.; Matic, S.; Gullino, M.L.; Infantino, A.; et al. Identification of bakanae disease resistance loci in japonica rice through genome wide association study. Rice 2017, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Munkvold, G.P. Fusarium species and their associated mycotoxins. Methods Mol. Biol. 2017, 1542, 51–106. [Google Scholar]
- Suthin Raj, T.; Muthukumar, A.; Renganathan, P.; Sudha Raja Kumar, R.; Ann Suji, H. Biological control of sheath blight of rice caused by Rhizoctonia solani kuhn using marine associated Bacillus subtilis. Int. Arch. Appl. Sci. Technol. 2019, 10, 148–153. [Google Scholar]
- Niño-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef]
- Pu, X.M.; Zhou, J.N.; Lin, B.R.; Shen, H.F. First report of bacterial foot rot of rice caused by a Dickeya zeae in china. Plant Dis. 2012, 96, 1820. [Google Scholar] [CrossRef]
- Cui, Z.-Q.; Zhu, B.; Xie, G.-L.; Li, B.; Huang, S.-W. Research status and prospect of Burkholderia glumae, the pathogen causing bacterial panicle blight. Rice Sci. 2016, 23, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Geering, A.D.W.; Randles, J.W. Virus Diseases of Tropical Crops; John Wiley and Sons: Chichester, UK, 2012. [Google Scholar]
- Breitler, J.C.; Cordero, M.J.; Royer, M.; Meynard, D.; San Segundo, B.; Guiderdoni, E. The −689/+197 region of the maize protease inhibitor gene directs high level, wound-inducible expression of the Cry1b gene which protects transgenic rice plants from stemborer attack. Mol. Breed. 2001, 7, 259–274. [Google Scholar] [CrossRef]
- Wei, Z.; Hu, W.; Lin, Q.; Cheng, X.; Tong, M.; Zhu, L.; Chen, R.; He, G. Understanding rice plant resistance to the brown planthopper (Nilaparvata lugens): A proteomic approach. Proteomics 2009, 9, 2798–2808. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, M.; Krishnappa, J. Rice root-knot nematode (Meloidogyne graminicola) an emerging problem integrataed management of pomegranate diseases. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3143–3171. [Google Scholar]
- Kiba, T.; Krapp, A. Plant nitrogen acquisition under low availability: Regulation of uptake and root architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Hinsinger, P.; Betencourt, E.; Bernard, L.; Brauman, A.; Plassard, C.; Shen, J.; Tang, X.; Zhang, F. P for two, sharing a scarce resource: Soil phosphorus acquisition in the rhizosphere of intercropped species. Plant Physiol. 2011, 156, 1078–1086. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Zhang, D. The plasticity of root systems in response to external phosphate. Int. J. Mol. Sci. 2020, 21, 5955. [Google Scholar] [CrossRef]
- Malhotra, H.; Vandana; Sharma, S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 171–190. [Google Scholar]
- Puga, M.I.; Rojas-Triana, M.; de Lorenzo, L.; Leyva, A.; Rubio, V.; Paz-Ares, J. Novel signals in the regulation of pi starvation responses in plants: Facts and promises. Curr. Opin. Plant Biol. 2017, 39, 40–49. [Google Scholar] [CrossRef]
- Chien, P.-S.; Chiang, C.-P.; Leong, S.J.; Chiou, T.-J. Sensing and signalling of phosphate starvation: From local to long distance. Plant Cell Physiol. 2018, 59, 1714–1722. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, F.; Lu, H.; Liu, Y.; Mao, C. Phosphate uptake and transport in plants: An elaborate regulatory system. Plant Cell Physiol. 2021. [Google Scholar] [CrossRef]
- Tadano, T.; Sakai, H. Soil science and plant nutrition secretion of acid phosphatase by the roots of several crop species under phosphorus-deficient conditions. Soil Sci. Plant Nutr. 2012, 37, 129–140. [Google Scholar] [CrossRef]
- Gho, Y.S.; Choi, H.; Moon, S.; Song, M.Y.; Park, H.E.; Kim, D.H.; Ha, S.H.; Jung, K.H. Phosphate-starvation-inducible S-like RNAse genes in rice are involved in phosphate source recycling by RNA decay. Front. Plant Sci. 2020, 11, 585561. [Google Scholar] [CrossRef]
- Shukla, D.; Rinehart, C.A.; Sahi, S.V. Comprehensive study of excess phosphate response reveals ethylene mediated signalling that negatively regulates plant growth and development. Sci. Rep. 2017, 7, 3074. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.K.; Shankar, A.; Chandran, A.K.N.; Sharma, M.; Jung, K.H.; Suprasanna, P.; Pandey, G.K. Emerging concepts of potassium homeostasis in plants. J. Exp. Bot. 2020, 71, 608–619. [Google Scholar] [CrossRef]
- Rout, G.R.; Sahoo, S. Role of iron in plant growth and metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Aung, M.S.; Masuda, H. How does rice defend against excess iron?: Physiological and molecular mechanisms. Front. Plant Sci. 2020, 11, 1102. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M.; Rashid, A.; Nadeem, F.; Stuerz, S.; Asch, F.; Bell, R.W.; Siddique, K.H.M. Boron nutrition of rice in different production systems. Agron. Sustain. Dev. 2018, 38, 25. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Ali, S.; Adrees, M.; Rizvi, H.; Zia-ur-Rehman, M.; Hannan, F.; Qayyum, M.F.; Hafeez, F.; Ok, Y.S. Cadmium stress in rice: Toxic effects, tolerance mechanisms, and management: A critical review. Environ. Sci. Pollut. Res. 2016, 23, 17859–17879. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defence. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol. 2016, 16, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, D.; Wang, G.; Zhou, J.M. Receptor kinases in plant-pathogen interactions: More than pattern recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, S.; Savatin, D.V.; Sicilia, F.; Gramegna, G.; Cervone, F.; De Lorenzo, G. Oligogalacturonides: Plant damage-associated molecular patterns and regulators of growth and development. Front. Plant Sci. 2013, 4, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Choi, J.; Cao, Y.; Stacey, G. Extracellular ATP acts as a Damage-Associated Molecular Pattern (DAMP) signal in plants. Front. Plant Sci. 2014, 5, 446. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Tsuda, K. Intimate Association of PRR- and NLR-mediated signalling in plant immunity. Mol. Plant Microbe Interact. 2021, 34, 3–14. [Google Scholar] [CrossRef]
- Peng, Y.; Van Wersch, R.; Zhang, Y. Convergent and divergent signalling in pamp-triggered immunity and effector-triggered immunity. Mol. Plant Microbe Interact. 2018, 31, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Seybold, H.; Trempel, F.; Ranf, S.; Scheel, D.; Romeis, T.; Lee, J. Ca2+ signalling in plant immune response: From pattern recognition receptors to Ca2+ decoding mechanisms. New Phytol. 2014, 204, 782–790. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signalling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, X.; Fan, B.; Zhu, C.; Chen, Z. Regulation and function of defence-related callose deposition in plants. Int. J. Mol. Sci. 2021, 22, 2393. [Google Scholar] [CrossRef]
- Van Meeteren, U.; Kaiser, E.; Malcolm Matamoros, P.; Verdonk, J.C.; Aliniaeifard, S. Is nitric oxide a critical key factor in ABA-induced stomatal closure? J. Exp. Bot. 2020, 71, 399–410. [Google Scholar] [CrossRef]
- Hasegawa, M.; Mitsuhara, I.; Seo, S.; Imai, T.; Koga, J.; Okada, K.; Yamane, H.; Ohashi, Y. Phytoalexin accumulation in the interaction between rice and the blast fungus. Mol. Plant Microbe Interact. 2010, 23, 1000–1011. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, M.; Mitsuhara, I.; Seo, S.; Okada, K.; Yamane, H.; Iwai, T.; Ohashi, Y. Analysis on blast fungus-responsive characters of a flavonoid phytoalexin sakuranetin; accumulation in infected rice leaves, antifungal activity and detoxification by fungus. Molecules 2014, 19, 11404–11418. [Google Scholar] [CrossRef]
- He, Y.; Xu, J.; Wang, X.; He, X.; Wang, Y.; Zhou, J.; Zhang, S.; Meng, X. The Arabidopsis pleiotropic drug resistance transporters PEN3 and PDR12 mediate camalexin secretion for resistance to Botrytis Cinerea. Plant Cell 2019, 31, 2206–2222. [Google Scholar] [CrossRef]
- Sanchez-Vallet, A.; Ramos, B.; Bednarek, P.; López, G.; Piślewska-Bednarek, M.; Schulze-Lefert, P.; Molina, A. Tryptophan-derived secondary metabolites in Arabidopsis thaliana confer non-host resistance to necrotrophic Plectosphaerella cucumerina fungi. Plant J. 2010, 63, 115–127. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Aerts, N.; Pereira Mendes, M.; Van Wees, S.C.M. Multiple levels of crosstalk in hormone networks regulating plant defence. Plant J. 2021, 105, 489–504. [Google Scholar] [CrossRef]
- Seo, J.K.; Wu, J.; Lii, Y.; Li, Y.; Jin, H. Contribution of small RNA pathway components in plant immunity. Mol. Plant Microbe Interact. 2013, 26, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Staiger, D.; Korneli, C.; Lummer, M.; Navarro, L. Emerging role for RNA-based regulation in plant immunity. New Phytol. 2013, 197, 394–404. [Google Scholar] [CrossRef]
- Islam, W.; Noman, A.; Qasim, M.; Wang, L. Plant responses to pathogen attack: Small rnas in focus. Int. J. Mol. Sci. 2018, 19, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bundó, M.; Campo, S.; San Segundo, B. Role of microRNAs in plant–fungus interactions. In Plant microRNAs; Miguel, C., Dalmay, T., Chaves, I., Eds.; Springer: Cham, Switzerland, 2020; pp. 199–220. [Google Scholar]
- Llave, C.; Xie, Z.; Kasschau, K.D.; Carrington, J.C. Cleavage of scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA. Science 2002, 297, 2053–2056. [Google Scholar] [CrossRef] [Green Version]
- Brodersen, P.; Sakvarelidze-Achard, L.; Bruun-Rasmussen, M.; Dunoyer, P.; Yamamoto, Y.Y.; Sieburth, L.; Voinnet, O. Widespread translational inhibition by plant miRNAs and siRNAs. Science 2008, 320, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Axtell, M.J. Classification and comparison of small RNAs from plants. Annu. Rev. Plant Biol. 2013, 64, 137–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell Biol. 2015, 16, 727–741. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Jia, T.; Chen, X. The ‘how’ and ‘where’ of plant microRNAs. New Phytol. 2017, 16, 1002–1017. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Mei, J.; Ren, G. Plant microRNAs: Biogenesis, homeostasis, and degradation. Front. Plant Sci. 2019, 27, 360. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, 155–162. [Google Scholar] [CrossRef]
- Jeong, D.-H.; Green, P.J. The Role of Rice microRNAs in abiotic stress responses. J. Plant Biol 2013, 56, 187–197. [Google Scholar] [CrossRef]
- Kumar, R. Role of microRNAs in biotic and abiotic stress responses in crop plants. Appl. Biochem. Biotechnol. 2014, 174, 93–115. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Bellinger, M.; Jin, H. Small RNAs: A new paradigm in plant-microbe interactions. Annu. Rev. Phytopathol. 2014, 52, 495–516. [Google Scholar] [CrossRef]
- Li, C.; Zhang, B. microRNAs in control of plant development. J. Cell. Physiol. 2016, 231, 303–313. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Yu, Y.; Wang, C.Y.; Li, Z.Y.; Liu, Q.; Xu, J.; Liao, J.Y.; Wang, X.J.; Qu, L.H.; Chen, F.; et al. Overexpression of microRNA OsmiR397 improves rice yield by increasing grain size and promoting panicle branching. Nat. Biotechnol. 2013, 31, 848–852. [Google Scholar] [CrossRef]
- Hu, J.; Zeng, T.; Xia, Q.; Qian, Q.; Yang, C.; Ding, Y.; Chen, L.; Wang, W. Unravelling miRNA regulation in yield of rice (Oryza sativa) based on differential network model. Sci. Rep. 2018, 8, 8498. [Google Scholar] [CrossRef]
- Chiou, T.-J. Regulation of phosphate homeostasis by microRNA in Arabidopsis. Plant Cell 2006, 18, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D.G. A plant miRNA contributes to antibacterial resistance by repressing auxin signalling. Science 2006, 312, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Baldrich, P.; Campo, S.; Wu, M.T.; Liu, T.T.; Hsing, Y.I.C.; San Segundo, B. microRNA-mediated regulation of gene expression in the response of rice plants to fungal elicitors. RNA Biol. 2015, 12, 847–863. [Google Scholar] [CrossRef] [Green Version]
- Camargo-Ramírez, R.; Val-Torregrosa, B.; San Segundo, B. MiR858-mediated regulation of flavonoid-specific MYB transcription factor genes controls resistance to pathogen infection in Arabidopsis. Plant Cell Physiol. 2017, 59, 190–204. [Google Scholar] [CrossRef]
- Soto-Suárez, M.; Baldrich, P.; Weigel, D.; Rubio-Somoza, I.; San Segundo, B. The Arabidopsis miR396 mediates pathogen-associated molecular pattern-triggered immune responses against fungal pathogens. Sci. Rep. 2017, 7, 44898. [Google Scholar] [CrossRef] [Green Version]
- Salvador-Guirao, R.; Baldrich, P.; Weigel, D.; RubioSo, B.S. The microRNA miR773 is involved in the Arabidopsis immune response to fungal pathogens. Mol. Plant Microbe Interact. 2018, 31, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Zheng, F.; Wei, S.; Zhang, S.; Li, G.; Cao, P.; Zhao, S. Evolution of disease defence genes and their regulators in plants. Int. J. Mol. Sci. 2019, 20, 335. [Google Scholar] [CrossRef] [Green Version]
- Campos-Soriano, L.; Bundó, M.; Bach-Pages, M.; Chiang, S.F.; Chiou, T.J.; San Segundo, B. Phosphate excess increases susceptibility to pathogen infection in rice. Mol. Plant Pathol. 2020, 21, 555–570. [Google Scholar] [CrossRef] [Green Version]
- Campo, S.; Sánchez-Sanuy, F.; Camargo-Ramírez, R.; Gómez-Ariza, J.; Baldrich, P.; Campos-Soriano, L.; Soto-Suárez, M.; San Segundo, B. A novel transposable element-derived microRNA participates in plant immunity to rice blast disease. Plant Biotechnol. J. 2021. [Google Scholar] [CrossRef]
- Feng, Q.; Li, Y.; Zhao, Z.-X.; Wang, W.-M. Contribution of small RNA Pathway to interactions of rice with pathogens and insect pests. Rice 2021, 14, 15. [Google Scholar] [CrossRef]
- Lu, J.; Yang, X.; Chen, J.; Li, T.; Hu, Z.; Xie, Y.; Li, J.; Zhao, J.; Pu, M.; Feng, H.; et al. Osa-miR439 negatively regulates rice immunity against Magnaporthe oryzae. Rice Sci. 2021, 28, 156–165. [Google Scholar] [CrossRef]
- Campo, S.; Peris-Peris, C.; Siré, C.; Moreno, A.B.; Donaire, L.; Zytnicki, M.; Notredame, C.; Llave, C.; San Segundo, B. Identification of a novel microRNA (miRNA) from rice that targets an alternatively spliced transcript of the Nramp6 (Natural Resistance-Associated Macrophage Protein 6) gene involved in pathogen resistance. New Phytol. 2013, 199, 212–227. [Google Scholar] [CrossRef]
- Salvador-Guirao, R.; Hsing, Y.I.; San Segundo, B. The Polycistronic miR166k-166h positively regulates rice immunity via post-transcriptional control of EIN2. Front. Plant Sci. 2018, 9, 337. [Google Scholar] [CrossRef]
- Wang, Z.; Xia, Y.; Lin, S.; Wang, Y.; Guo, B.; Song, X.; Ding, S.; Zheng, L.; Feng, R.; Chen, S.; et al. Osa-miR164a targets OsNAC60 and negatively regulates rice immunity against the blast fungus Magnaporthe oryzae. Plant J. 2018, 95, 584–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Bao, Y.; Shan, D.; Wang, Z.; Song, X.; Wang, Z.; Wang, J.; He, L.; Wu, L.; Zhang, Z.; et al. Magnaporthe oryzae defeats rice defence by inducing miR319b and suppressing jasmonic acid signalling. Plant Physiol. 2018, 184, 1665. [Google Scholar]
- Chandran, V.; Wang, H.; Gao, F.; Cao, X.-L.; Chen, Y.-P.; Li, G.-B.; Zhu, Y.; Yang, X.-M.; Zhang, L.-L.; Zhao, Z.-X.; et al. MiR396-OsGRFs module balances growth and rice blast disease-resistance. Front. Plant Sci. 2019, 9, 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Cao, X.L.; Zhu, Y.; Yang, X.M.; Zhang, K.N.; Xiao, Z.Y.; Wang, H.; Zhao, J.H.; Zhang, L.L.; Li, G.B.; et al. Osa-MiR398 boosts H2O2 production and rice blast disease-resistance via multiple superoxide dismutases. New Phytol. 2019, 222, 1507–1522. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Sanuy, F.; Peris-Peris, C.; Tomiyama, S.; Okada, K.; Hsing, Y.I.; San Segundo, B.; Campo, S. Osa-miR7695 enhances transcriptional priming in defence responses against the rice blast fungus. BMC Plant Biol. 2019, 19, 563. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.X.; Zhu, Y.; Wang, L.F.; Zheng, Y.P.; Chen, J.F.; Li, T.T.; Yang, X.M.; Wang, H.; Li, X.P.; Ma, X.C.; et al. Osa-miR1873 fine-tunes rice immunity against Magnaporthe oryzae and yield traits. J. Integr. Plant Biol. 2020, 62, 1213–1226. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lu, Y.G.; Shi, Y.; Wu, L.; Xu, Y.J.; Huang, F.; Guo, X.Y.; Zhang, Y.; Fan, J.; Zhao, J.Q.; et al. multiple rice microRNAs are involved in immunity against the blast fungus Magnaporthe oryzae. Plant Physiol. 2014, 164, 1077–1092. [Google Scholar] [CrossRef] [Green Version]
- Helliwell, E.E.; Wang, Q.; Yang, Y. Ethylene biosynthesis and signalling is required for rice immune response and basal resistance against Magnaporthe oryzae infection. Mol. Plant Microbe Interact. 2016, 29, 831–843. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Liu, M.; Tang, M.; Dong, B.; Wu, D.; Zhang, Z.; Zhou, B. Repression of microRNA biogenesis by silencing of OsDCL1 activates the basal resistance to Magnaporthe oryzae in rice. Plant Sci. 2015, 237, 24–32. [Google Scholar] [CrossRef]
- Salvador-Guirao, R.; Baldrich, P.; Tomiyama, S.; Hsing, Y.I.; Okada, K.; San Segundo, B. OsDCL1a activation impairs phytoalexin biosynthesis and compromises disease resistance in rice. Ann. Bot. 2019, 123, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Veresoglou, S.D.; Barto, E.K.; Menexes, G.; Rillig, M.C. Fertilization affects severity of disease caused by fungal plant pathogens. Plant Pathol. 2013, 62, 961–969. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 16, 525–537. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, M.; Mur, L.A.J.; Shen, Q.; Guo, S. Unravelling the roles of nitrogen nutrition in plant disease defences. Int. J. Mol. Sci. 2020, 21, 572. [Google Scholar] [CrossRef] [Green Version]
- Ballini, E.; Nguyen, T.T.T.; Morel, J.B. Diversity and genetics of nitrogen-induced susceptibility to the blast fungus in rice and wheat. Rice 2013, 6, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Wu, L.; Ding, Y.; Yao, X.; Wu, X.; Weng, F.; Li, G.; Liu, Z.; Tang, S.; Ding, C.; et al. Nitrogen fertilizer application affects lodging resistance by altering secondary cell wall synthesis in japonica rice (Oryza sativa). J. Plant Res. 2017, 130, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Nguyen Thi Thu, T.; He, X.; Gravot, A.; Bernillon, S.; Ballini, E.; Morel, J.B. Increase of fungal pathogenicity and role of plant glutamine in nitrogen-induced susceptibility (NIS) to rice blast. Front. Plant Sci. 2017, 8, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangol, S.; Chen, Y.; Hwang, B.K.; Jwa, N.S. Iron- and reactive oxygen species-dependent ferroptotic cell death in rice-Magnaporthe oryzae interactions. Plant Cell 2019, 31, 189–209. [Google Scholar] [CrossRef] [Green Version]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Law, T.F.; De Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, C.D.; et al. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef]
- Chan, C.; Liao, Y.-Y.; Chiou, T.-J. The impact of phosphorus on plant immunity. Plant Cell Physiol. 2021. [Google Scholar] [CrossRef]
- Dong, Z.; Li, W.; Liu, J.; Li, L.; Pan, S.; Liu, S.; Gao, J.; Liu, L.; Liu, X.; Wang, G.L.; et al. The rice phosphate transporter protein OsPT8 regulates disease resistance and plant growth. Sci. Rep. 2019, 9, 5408. [Google Scholar] [CrossRef]
- Vallino, M.; Greppi, D.; Novero, M.; Bonfante, P.; Lupotto, E. Rice root colonisation by mycorrhizal and endophytic fungi in aerobic soil. Ann. Appl. Biol. 2009, 154, 195–204. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef]
- Bonfante, P.; Genre, A. Mechanisms Underlying Beneficial Plant—Fungus Interactions in Mycorrhizal Symbiosis. Nat. Commun. 2010, 1, 48. [Google Scholar] [CrossRef] [Green Version]
- Maclean, A.M.; Bravo, A.; Harrison, M.J. Plant signalling and metabolic pathways enabling arbuscular mycorrhizal symbiosis. Plant Cell 2017, 29, 2319–2335. [Google Scholar] [CrossRef]
- Choi, J.; Summers, W.; Paszkowski, U. Mechanisms underlying establishment of arbuscular mycorrhizal symbioses. Annu. Rev. Phytopathol. 2018, 56, 135–160. [Google Scholar] [CrossRef]
- Vallino, M.; Fiorilli, V.; Bonfante, P. Rice flooding negatively impacts root branching and arbuscular mycorrhizal colonization, but not fungal viability. Plant Cell Environ. 2014, 37, 557–572. [Google Scholar] [CrossRef]
- Gutjahr, C.; Sawers, R.J.H.; Marti, G.; Andrés-Hernández, L.; Yang, S.Y.; Casieri, L.; Angliker, H.; Oakeley, E.J.; Wolfender, J.L.; Abreu-Goodger, C.; et al. Transcriptome diversity among rice root types during asymbiosis and interaction with arbuscular mycorrhizal fungi. Proc. Natl. Acad. Sci. USA 2015, 112, 6754–6759. [Google Scholar] [CrossRef] [Green Version]
- Pimprikar, P.; Gutjahr, C. Transcriptional regulation of arbuscular mycorrhiza development. Plant Cell Physiol. 2018, 59, 673–690. [Google Scholar] [CrossRef]
- Gutjahr, C.; Casieri, L.; Paszkowski, U. Glomus intraradices induces changes in root system architecture of rice independently of common symbiosis signalling. New Phytol. 2009, 182, 829–837. [Google Scholar] [CrossRef]
- Davidson, H.; Shrestha, R.; Cornulier, T.; Douglas, A.; Travis, T.; Johnson, D.; Price, A.H. Spatial effects and GWA mapping of root colonization assessed in the interaction between the rice diversity panel 1 and an arbuscular mycorrhizal fungus. Front. Plant Sci. 2019, 10, 633. [Google Scholar] [CrossRef] [Green Version]
- Campos-Soriano, L.; García-Garrido, J.M.; San Segundo, B. Activation of basal defence mechanisms of rice plants by Glomus intraradices does not affect the arbuscular mycorrhizal symbiosis. New Phytol. 2010, 188, 597–614. [Google Scholar] [CrossRef]
- Campos-Soriano, L.; García-Martínez, J.; San Segundo, B. The arbuscular mycorrhizal symbiosis promotes the systemic induction of regulatory defence-related genes in rice leaves and confers resistance to pathogen infection. Mol. Plant Pathol. 2012, 13, 579–592. [Google Scholar] [CrossRef]
- Campo, S.; San Segundo, B. Systemic induction of phosphatidylinositol-based signalling in leaves of arbuscular mycorrhizal rice plants. Sci. Rep. 2020, 10, 15896. [Google Scholar] [CrossRef]
- Campo, S.; Martín-Cardoso, H.; Olivé, M.; Pla, E.; Catala-Forner, M.; Martínez-Eixarch, M.; San Segundo, B. Effect of root colonization by arbuscular mycorrhizal fungi on growth, productivity and blast resistance in rice. Rice 2020, 13, 42. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Sun, W.; Wang, T. The adaptive mechanism of plants to iron deficiency via iron uptake, transport, and homeostasis. Int. J. Mol. Sci. 2019, 20, 2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimaru, Y.; Suzuki, M.; Tsukamoto, T.; Suzuki, K.; Nakazono, M.; Kobayashi, T.; Wada, Y.; Watanabe, S.; Matsuhashi, S.; Takahashi, M.; et al. Rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+. Plant J. 2006, 45, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Peris-Peris, C.; Serra-Cardona, A.; Śanchez-Sanuy, F.; Campo, S.; Ariño, J.; San Segundo, B. Two NRAMP6 isoforms function as iron and manganese transporters and contribute to disease resistance in rice. Mol. Plant Microbe Interact. 2017, 30, 385–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbon, E.H.; Trapet, P.L.; Stringlis, I.A.; Kruijs, S.; Bakker, P.A.H.M.; Pieterse, C.M.J. Iron and Immunity. Annu. Rev. Phytopathol. 2017, 55, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Herlihy, J.H.; Long, T.A.; McDowell, J.M. Iron homeostasis and plant immune responses: Recent insights and translational implications. J. Biol. Chem. 2020, 295, 13444–13457. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Greenshields, D.L.; Sammynaiken, R.; Hirji, R.N.; Selvaraj, G.; Wei, Y. Targeted alterations in iron homeostasis underlie plant defence responses. J. Cell Sci. 2007, 120, 596–605. [Google Scholar] [CrossRef] [Green Version]
- Trapet, P.L.; Verbon, E.H.; Bosma, R.R.; Voordendag, K.; Van Pelt, J.A.; Pieterse, C.M.J. Mechanisms underlying iron deficiency-induced resistance against pathogens with different lifestyles. J. Exp. Bot. 2020, 72, 2231–2241. [Google Scholar] [CrossRef]
- Yuan, M.; Wang, S.; Chu, Z.; Li, X.; Xu, C. The bacterial pathogen Xanthomonas oryzae overcomes rice defences by regulating host copper redistribution. Plant Cell 2010, 22, 3164–3176. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; Ai, Q.; Yu, D. Uncovering MiRNAs Involved in crosstalk between nutrient deficiencies in Arabidopsis. Sci. Rep. 2015, 5, 11813. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Datta, S.K.; Datta, K. MiRNA Regulation of nutrient homeostasis in plants. Front. Plant Sci. 2015, 6, 232. [Google Scholar] [CrossRef] [Green Version]
- Fujii, H.; Chiou, T.-J.; Lin, S.-I.; Aung, K.; Zhu, J.-K. A MiRNA involved in phosphate-starvation response in Arabidopsis. Curr. Biol. 2005, 15, 2038–2043. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, L.C.; Lin, S.I.; Shih, A.C.C.; Chen, J.W.; Lin, W.Y.; Tseng, C.Y.; Li, W.H.; Chiou, T.J. Uncovering small RNA-mediated responses to phosphate deficiency in Arabidopsis by deep sequencing. Plant Physiol. 2009, 151, 2120–2132. [Google Scholar] [CrossRef] [Green Version]
- Ham, B.K.; Chen, J.; Yan, Y.; Lucas, W.J. Insights into plant phosphate sensing and signalling. Curr. Opin. Biotechnol. 2018, 49, 1–9. [Google Scholar] [CrossRef]
- Aung, K. pho2, a Phosphate Overaccumulator, is caused by a nonsense mutation in a microRNA399 target gene. Plant Physiol. 2006, 141, 1000–1011. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.Y.; Huang, T.K.; Chiou, T.J. Nitrogen limitation adaptation, a target of microRNA827, mediates degradation of plasma membrane-localized phosphate transporters to maintain phosphate homeostasis in Arabidopsis. Plant Cell 2013, 25, 4061–4074. [Google Scholar] [CrossRef] [Green Version]
- Park, B.S.; Seo, J.S.; Chua, N.H. NITROGEN LIMITATION ADAPTATION recruits PHOSPHATE2 to target the phosphate transporter PT2 for degradation during the regulation of Arabidopsis phosphate homeostasis. Plant Cell 2014, 26, 454–464. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.I.; Santi, C.; Jobet, E.; Lacut, E.; El Kholti, N.; Karlowski, W.M.; Verdeil, J.L.; Breitler, J.C.; Périn, C.; Ko, S.S.; et al. Complex regulation of two target genes encoding SPX-MFS Proteins by rice miR827 in response to phosphate starvation. Plant Cell Physiol. 2010, 51, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.Y.; Lin, Y.Y.; Chiang, S.F.; Syu, C.; Hsieh, L.C.; Chiou, T.J. Evolution of microRNA827 targeting in the plant kingdom. New Phytol. 2018, 217, 1712–1725. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Huang, W.; Ying, Y.; Li, S.; Secco, D.; Tyerman, S.; Whelan, J.; Shou, H. Functional characterization of the rice SPX-MFS family reveals a key role of OsSPX-MFS1 in controlling phosphate homeostasis in leaves. New Phytol. 2012, 196, 139–148. [Google Scholar] [CrossRef]
- Liu, J.; Fu, S.; Yang, L.; Luan, M.; Zhao, F.; Luan, S.; Lan, W. Vacuolar SPX-MFS transporters are essential for phosphate adaptation in plants. Plant Signal. Behav. 2016, 11, 1213474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, B.X.; Lima-Melo, Y.; dos Santos Maraschin, F.; Margis-Pinheiro, M. Phosphate starvation responses in crop roots: From well-known players to novel candidates. Environ. Exp. Bot. 2020, 178, 104162. [Google Scholar] [CrossRef]
- Sunkar, R.; Kapoor, A.; Zhu, J.K. Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. Plant Cell 2006, 18, 2051–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauclair, L.; Yu, A.; Bouché, N. microRNA-directed cleavage and translational repression of the copper chaperone for superoxide dismutase mRNA in Arabidopsis. Plant J. 2010, 62, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Guo, D.; Zhang, J.; Huang, Q.; Qin, G.; Zhang, X.; Wan, J.; Gu, H.; Qu, L.J. Targeted mutagenesis in rice using CRISPR-Cas system. Cell Res. 2013, 23, 1233–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Deng, K.; Cheng, Y.; Zhong, Z.; Tian, L.; Tang, X.; Tang, A.; Zheng, X.; Zhang, T.; Qi, Y.; et al. CRISPR-Cas9 based genome editing reveals new insights into microRNA function and regulation in rice. Front. Plant Sci. 2017, 8, 1598. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Colonisation of rice leaves by the hemibiotroph fungus Magnaporthe oryzae. The fungus first establishes a biotrophic interaction with the rice plant and later switches to a necrotrophic lifestyle. During the biotrophic phase, the fungus grows inside living host cells. The hyphae are encased by the extra-invasive hyphal membrane (EIHM) which is continuous with the rice plasma membrane. At this stage of the infection process, the fungus uses plasmodesmata for cell-to-cell movement [16,26], and absorbs nutrients from the plant living cells. The host vacuolar integrity in the first invaded rice cells is important for M. oryzae early stages of infection [27]. In the necrotrophic phase, the fungus colonises the host tissue and obtains nutrients from the host dead cells. Ap, appresorium; EIHM, extra invasive hyphal membrane; IH, invasive hyphae; Pd, plasmodesmata; PP, penetration peg; Sp, spores; Vac, vacuole. Black triangles represent plant nutrients.
Figure 1.
Colonisation of rice leaves by the hemibiotroph fungus Magnaporthe oryzae. The fungus first establishes a biotrophic interaction with the rice plant and later switches to a necrotrophic lifestyle. During the biotrophic phase, the fungus grows inside living host cells. The hyphae are encased by the extra-invasive hyphal membrane (EIHM) which is continuous with the rice plasma membrane. At this stage of the infection process, the fungus uses plasmodesmata for cell-to-cell movement [16,26], and absorbs nutrients from the plant living cells. The host vacuolar integrity in the first invaded rice cells is important for M. oryzae early stages of infection [27]. In the necrotrophic phase, the fungus colonises the host tissue and obtains nutrients from the host dead cells. Ap, appresorium; EIHM, extra invasive hyphal membrane; IH, invasive hyphae; Pd, plasmodesmata; PP, penetration peg; Sp, spores; Vac, vacuole. Black triangles represent plant nutrients.

Figure 2.
miRNAs with a known function in the response of rice plants to infection by the blast fungus M. oryzae. The miRNA/target gene modules and the biological processes that they regulate are indicated. ACO, 1-aminocyclopropane-1-carboxylase oxidase3; ARF12, Auxin response factor; Chl, Chloroplast; CIPK, Calcineurin B-Like (CBL)-interacting serine– threonine protein kinase; CSD, Copper superoxide dismutase; DCL1, Dicer-like protein; EIN, Ethylene-insensitive protein 2; GRF, Growth regulating factors; LRR, Leucin-rich repeat; NAC, No apical meristem; NF-YA, Nuclear transcription factor Y subunit alpha; NRAMP, Natural resistance-associated macrophage protein; Nuc, Nucleus; PHO, Phosphate; PTs, Phosphate transporters; ROS, Reactive oxygen species; SPL, SQUAMOSA promoter-binding protein-like transcription factor; Sp, Spores; TCP, Teosinte branched/cycloidea/pcf; Vac, Vacuole.
Figure 2.
miRNAs with a known function in the response of rice plants to infection by the blast fungus M. oryzae. The miRNA/target gene modules and the biological processes that they regulate are indicated. ACO, 1-aminocyclopropane-1-carboxylase oxidase3; ARF12, Auxin response factor; Chl, Chloroplast; CIPK, Calcineurin B-Like (CBL)-interacting serine– threonine protein kinase; CSD, Copper superoxide dismutase; DCL1, Dicer-like protein; EIN, Ethylene-insensitive protein 2; GRF, Growth regulating factors; LRR, Leucin-rich repeat; NAC, No apical meristem; NF-YA, Nuclear transcription factor Y subunit alpha; NRAMP, Natural resistance-associated macrophage protein; Nuc, Nucleus; PHO, Phosphate; PTs, Phosphate transporters; ROS, Reactive oxygen species; SPL, SQUAMOSA promoter-binding protein-like transcription factor; Sp, Spores; TCP, Teosinte branched/cycloidea/pcf; Vac, Vacuole.

Figure 3.
Crosstalk between nutrient signalling pathways and immune response. Nutrients are essential for plant growth and development. They are obtained from the soil through the plant root system. However, insufficient or excessive amounts of certain nutrients can affect disease resistance in plants. In rice, high nitrogen (N) supply not only promotes M. oryzae growth but also induces the expression of fungal effector genes, thus making the plant more susceptible to infection [117]. Growing rice plants under high phosphate (Pi) conditions leads to an increase in leaf Pi content, which is associated with a weaker activation of defence responses and susceptibility against the rice blast fungus [98]. Ferroptosis, an iron (Fe)- and ROS-dependent cell death process, contributes to arrest M. oryzae infection in incompatible rice/M. oryzae interactions [120]. Thus, iron levels must be strictly controlled in the host plant to maintain the subtle balance between growth and defence.
Figure 3.
Crosstalk between nutrient signalling pathways and immune response. Nutrients are essential for plant growth and development. They are obtained from the soil through the plant root system. However, insufficient or excessive amounts of certain nutrients can affect disease resistance in plants. In rice, high nitrogen (N) supply not only promotes M. oryzae growth but also induces the expression of fungal effector genes, thus making the plant more susceptible to infection [117]. Growing rice plants under high phosphate (Pi) conditions leads to an increase in leaf Pi content, which is associated with a weaker activation of defence responses and susceptibility against the rice blast fungus [98]. Ferroptosis, an iron (Fe)- and ROS-dependent cell death process, contributes to arrest M. oryzae infection in incompatible rice/M. oryzae interactions [120]. Thus, iron levels must be strictly controlled in the host plant to maintain the subtle balance between growth and defence.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Val-Torregrosa, B.; Bundó, M.; San Segundo, B. Crosstalk between Nutrient Signalling Pathways and Immune Responses in Rice. Agriculture 2021, 11, 747. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080747
AMA Style
Val-Torregrosa B, Bundó M, San Segundo B. Crosstalk between Nutrient Signalling Pathways and Immune Responses in Rice. Agriculture. 2021; 11(8):747. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080747
Chicago/Turabian StyleVal-Torregrosa, Beatriz, Mireia Bundó, and Blanca San Segundo. 2021. "Crosstalk between Nutrient Signalling Pathways and Immune Responses in Rice" Agriculture 11, no. 8: 747. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11080747
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.