Evaluation of Soil-Applied Chemical Fungicide and Biofungicide for Control of the Fusarium Wilt of Chrysanthemum and Their Effects on Rhizosphere Soil Microbiota

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Plant Material
2.2. Fungicide Preparation and Preassessment of the Inhibition of F. Oxysporum Growth
2.3. Field Experimental Design
2.4. Experimental Methods
2.4.1. Measurement of Chrysanthemum Growth and Quality
2.4.2. Disease Incidence
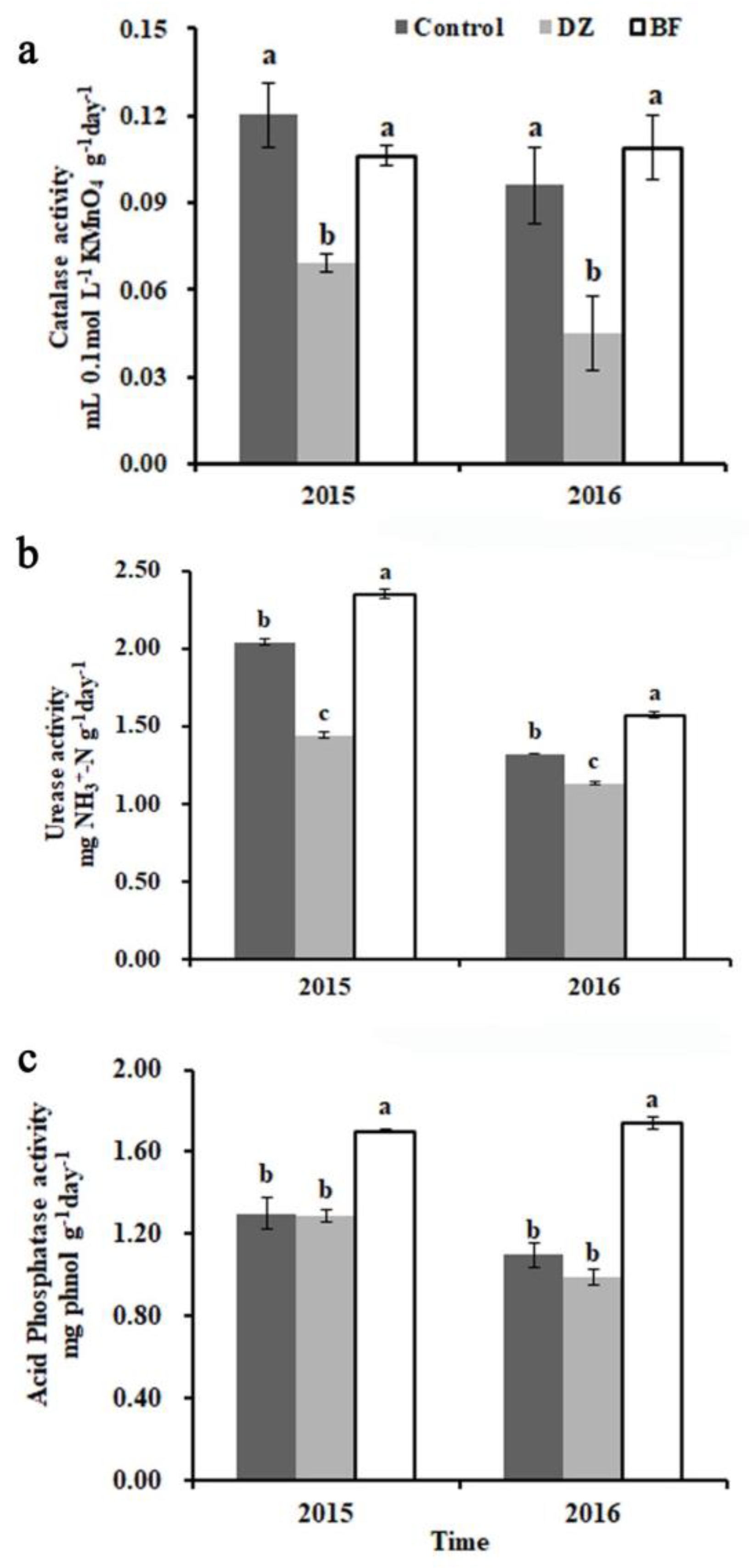
2.4.3. Soil-Enzyme Activity Analysis
2.4.4. Soil DNA Extraction
2.4.5. Real-Time Polymerase Chain Reaction (PCR) Amplification
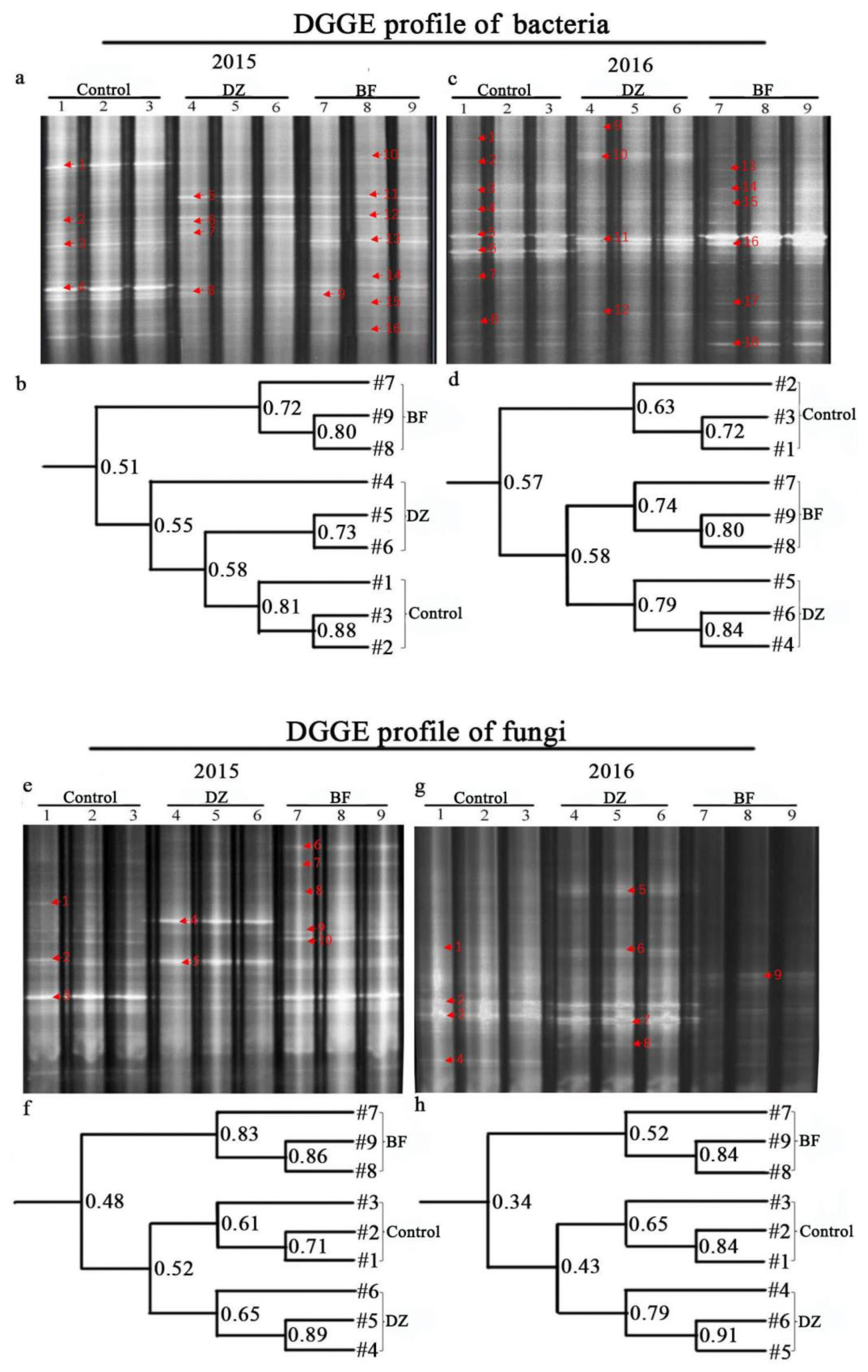
2.4.6. DGGE Profiling
2.5. Statistical Analyses
3. Results
3.1. Fungicide Preassessment
3.2. Effects of Fungicides on Chrysanthemum Fusarium Wilt
3.3. Effects of Fungicides on the Quality of Chrysanthemum
3.4. Effects of Fungicides on Chrysanthemum Soil Enzymatic Activities
3.5. Effects of Fungicides on Soil Microbiota Detected by Real-Time PCR
3.6. DGGE Fingerprints of DZ- and BF-Treated Rhizosphere Soil 16s and 18s Fragment
4. Discussion
5. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Song, A.; Zhao, S.; Chen, S.; Jiang, J.; Chen, S.; Li, H.; Chen, Y.; Chen, X.; Fang, W.; Chen, F. The abundance and diversity of soil fungi in continuously monocropped chrysanthemum. Sci. World J. 2013. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Chen, X.; Deng, S.; Dong, X.; Song, A.; Yao, J.; Fang, W.; Chen, F. The effects of fungicide, soil fumigant, bio-organic fertilizer and their combined application on chrysanthemum Fusarium wilt controlling, soil enzyme activities and microbial properties. Molecules 2016, 21, 526. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhong, M.; Zhang, X.; Xue, C.; Zhang, C.; Ma, T.; Zhang, S. Suppression of Fusarium wilt of cucumber by ammonia gas fumigation via reduction of Fusarium population in the field. Sci. Rep. 2017, 7, 43103. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Xu, Z.; Ling, N.; Yuan, Y.; Yang, X.; Chen, L.; Shen, B.; Shen, Q. Isolation and identification of lipopeptides produced by B. subtilis SQR 9 for suppressing Fusarium wilt of cucumber. Sci. Hortic. 2012, 135, 32–39. [Google Scholar] [CrossRef]
- Chen, F.; Wang, M.; Zheng, Y.; Luo, J.; Yang, X.; Wang, X. Quantitative changes of plant defense enzymes and phytohormone in biocontrol of cucumber Fusarium wilt by Bacillus subtilis B579. World J. Microbiol. Biotechnol. 2010, 26, 675–684. [Google Scholar] [CrossRef]
- Wu, H.; Yang, X.; Fan, J.; Miao, W.; Ling, N.; Xu, Y.; Huang, Q.; Shen, Q. Suppression of Fusarium wilt of watermelon by a bio-organic fertilizer containing combinations of antagonistic microorganisms. Biocontrol 2009, 54, 287–300. [Google Scholar] [CrossRef]
- Ling, N.; Zhang, W.; Tan, S.; Huang, Q.; Shen, Q. Effect of the nursery application of bioorganic fertilizer on spatial distribution of Fusarium oxysporum f. sp. niveum and its antagonistic bacterium in the rhizosphere of watermelon. Appl. Soil Ecol. 2012, 59, 13–19. [Google Scholar] [CrossRef]
- Chen, C.; Bauske, E.; Musson, G.; Rodriguezkabana, R.; Kloepper, J. Biological control of Fusarium wilt on cotton by use of endophytic bacteria. Biol. Control 1995, 5, 83–91. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y.; Shen, Z.; Xue, C.; Li, R.; Shen, Q. Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil Biol. Bioch. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Zhang, N.; He, X.; Zhang, J.; Raza, W.; Yang, X.; Ruan, Y.; Shen, Q.; Huang, Q. Suppression of Fusarium Wilt of Banana with application of bio-organic fertilizers. Pedosphere 2014, 24, 613–624. [Google Scholar] [CrossRef]
- Dean, R.A.; Lichens-Park, A.; Kole, C. Genomics of plant-associated fungi and oomycetes. Dicot Pathogl. 2014. [Google Scholar] [CrossRef]
- Gamliel, A.; Grinstein, A.; Peretz, Y.; Klein, L.; Nachmias, A.; Tsror, L.; Livescu, L.; Katan, J. Reduced dosage of methyl bromide for controlling verticillium wilt of potato in experimental and commercial plots. Plant Dis. 1997, 81, 469–474. [Google Scholar] [CrossRef]
- Barhate, B.G.; Musmade, N.A.; Nikhate, T.A. Management of Fusarium wilt of tomato by bioagents, fungicides and varietal resistance. Int. J. Plant Prot. 2015, 8, 49–52. [Google Scholar] [CrossRef]
- Nicola, L.; Turco, E.; Albanese, D.; Donati, C.; Thalheimer, M.; Pindo, M.; Insam, H.; Cavalieri, D.; Pertot, I. Fumigation with dazomet modifies soil microbiota in apple orchards affected by replant disease. Appl. Soil Ecol. 2017, 113, 71–79. [Google Scholar] [CrossRef]
- Wan, T.; Zhao, H.; Wang, W. Effect of biocontrol agent Bacillus amyloliquefaciens SN16-1 and plant pathogen Fusarium oxysporum on tomato rhizosphere bacterial community composition. Biol. Control 2017, 112, 1–9. [Google Scholar] [CrossRef]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Bioch. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P.; Crowley, D.; Lieberei, R. Arbuscular mycorrhizal infection changes the bacterial 16 S rDNA community composition in the rhizosphere of maize. Mycorrhiza 2001, 11, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Pérez, V.; Batlle, V.; Chacón, B.; Montenegro, M. Efficacy of Trichoderma harzianum A34 in the biocontrol of Fusarium oxysporum f.sp. cubense, the causal agent of Fusarium wilt or Panama disease of bananas in Cuba. Fitosanidad 2009, 13, 259–264. [Google Scholar]
- Mejri, S.; Siah, A.; Coutte, F.; Magnin-Robert, M.; Randoux, B.; Tisserant, B.; Krier, F.; Jacques, P.; Reignault, P.; Halama, P. Biocontrol of the wheat pathogen Zymoseptoria tritici using cyclic lipopeptides from Bacillus subtilis. Environ. Sci. Pollut. Res. Int. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ling, N.; Deng, K.; Song, Y.; Wu, Y.; Zhao, J.; Raza, W.; Huang, Q.; Shen, Q. Variation of rhizosphere bacterial community in watermelon continuous mono-cropping soil by long-term application of a novel bioorganic fertilizer. Microbiol. Res. 2014, 169, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Chen, X.H.; Budiharjo, A.; Bleiss, W.; Vater, J.; Borriss, R. Efficient colonization of plant roots by the plant growth promoting bacterium Bacillus amyloliquefaciens FZB42, engineered to express green fluorescent protein. J. Biotechnol. 2011, 151, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Lalloo, R.; Maharajh, D.; Görgens, J.; Gardiner, N. A downstream process for production of a viable and stable Bacillus cereus aquaculture biological agent. Appl. Microbiol. Biotechnol. 2010, 86, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Ramesh, A.; Sharma, M.P.; Joshi, O.P.; Govaerts, B.; Steenwerth, K.L.; Karlen, D.L. Microbial community structure and diversity as indicators for evaluating soil quality. Springer Neth. 2010. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, X.; Wang, X.; Shao, H.; Yang, J.; Wang, X. Soil enzymes as indicators of saline soil fertility under various soil amendments. Agric. Ecosyst. Environ. 2017, 237, 274–279. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Martínez, V.; Zobeck, T.M.; Gill, T.E.; Kennedy, A.C. Enzyme activities and microbial community structure in semiarid agricultural soils. Biol. Fertil. Soil 2003, 38, 216–227. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R.; González-Arenzana, L. Impact of chemical and biological fungicides applied to grapevine on grape biofilm, must, and wine microbial diversity. Front. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Shen, Z.; Zhang, F.; Raza, W.; Yuan, J.; Huang, R.; Ruan, Y.; Li, R.; Shen, Q. Bacillus amyloliquefaciens strain W19 can promote growth and yield and suppress Fusarium wilt in banana under greenhouse and field conditions. Pedosphere 2016, 26, 733–744. [Google Scholar] [CrossRef]
- Ge, Y.; Wang, Q.; Wang, L.; Liu, W.; Liu, X.; Huang, Y. Response of soil enzymes and microbial communities to root extracts of the alien Alternanthera philoxeroides. Arch. Agron. Soil Sci. 2017, 64, 708–717. [Google Scholar] [CrossRef]
- Garcı́a-Gil, J.C.; Plaza, C.; Soler-Rovira, P.; Polo, A. Long-term effects of municipal solid waste compost application on soil enzyme activities and microbial biomass. Soil Biol. Bioch. 2000, 32, 1907–1913. [Google Scholar] [CrossRef]
- Zhou, X.; Li, S.; Li, W.; Jiang, D.; Han, K.; Wu, Z.; Li, Y. Myxobacterial community is a predominant and highly diverse bacterial group in soil niches. Environ. Microbiol. Rep. 2014, 6, 45–56. [Google Scholar] [CrossRef] [PubMed]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Suppl. Mycol. 1990, 315–322. [Google Scholar]
- Lievens, B.; Claes, L.; Vanachter, A.C.R.C.; Cammue, B.P.A.; Thomma, B.P.H.J. Detecting single nucleotide polymorphisms using DNA arrays for plant pathogen diagnosis. Fems Microbiol. Lett. 2006, 255, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, X.; Xing, S.; Ma, Z.; Hu, S.; Tu, C. Bio-organic fertilizer promotes plant growth and yield and improves soil microbial community in continuous monoculture system of chrysanthemum morifolium cv. Chuju. Int. J. Agric. Biol. 2017, 19, 563–568. [Google Scholar] [CrossRef]
- Guo, Q.; Dong, W.; Li, S.; Lu, X.; Wang, P.; Zhang, X.; Wang, Y.; Ma, P. Fengycin produced by Bacillus subtilis NCD-2 plays a major role in biocontrol of cotton seedling damping-off disease. Microbiol. Res. 2014, 169, 533. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Zhang, R.; Xue, C.; Zhang, S.; Li, S.; Zhang, N.; Shen, Q. Application of bio-organic fertilizer can control Fusarium wilt of cucumber plants by regulating microbial community of rhizosphere soil. Biol. Fertil. Soil 2012, 48, 807–816. [Google Scholar] [CrossRef]
- Khezri, M.; Ahmadzadeh, M.; Jouzani, G.S.; Behboudi, K.; Ahangaran, A.; Mousivand, M.; Rahimian, H. Characterization of some biofilm-forming Bacillus subtilis strains and evaluation of their biocontrol potential against Fusarium culmorum. J. Plant Pathol. 2011, 93, 373–382. [Google Scholar] [CrossRef]
- Xiong, W.; Guo, S.; Jousset, A.; Zhao, Q.; Wu, H.; Li, R.; Kowalchuk, G.A.; Shen, Q. Bio-fertilizer application induces soil suppressiveness against Fusarium wilt disease by reshaping the soil microbiome. Soil Biol. Bioch. 2017, 114, 238–247. [Google Scholar] [CrossRef]
- Tian, T.; Li, S.; Sun, M. Synergistic effect of dazomet soil fumigation and clonostachys rosea against cucumber Fusarium wilt. Phytopathology 2014, 104, 1314. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Huang, X.; Chen, Q.; Liang, B.; Qiu, J.; Ali, S.W.; Li, S. Isolation and characterization of three Sphingobium sp. strains capable of degrading isoproturon and cloning of the catechol 1,2-dioxygenase gene from these strains. World J. Microbiol. Biotechnol. 2009, 25, 259–268. [Google Scholar] [CrossRef]
- Zhong, J.; Luo, L. Degradation pathways of 1-methylphenanthrene in bacterial Sphingobium sp. MP9-4 isolated from petroleum-contaminated soil. Mar. Pollut. Bull. 2016, 114, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Diagne, N.; Swanson, E.; Pesce, C.; Fall, F.; Diouf, F.; Bakhoum, N.; Fall, D.; Faye, M.N.; Oshone, R.; Simpson, S.; et al. Permanent draft genome sequence of Rhizobium sp. Strain LCM 4573, a salt-tolerant, nitrogen-fixing bacterium isolated from senegalese soils. Genome Announc 2017. [Google Scholar] [CrossRef] [PubMed]
- Marra, L.M.; Soares, C.R.F.S.; de Oliveira, S.M.; Ferreira, P.A.A.; Soares, B.L.; de Fráguas Carvalho, R.; de Lima, J.M.; de Souza Moreira, F.M. Biological nitrogen fixation and phosphate solubilization by bacteria isolated from tropical soils. Plant Soil 2012, 357, 289–307. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Index | 2015 | 2016 | |||||
|---|---|---|---|---|---|---|---|
| Control | DZ | BF | Control | DZ | BF | ||
| Shoot | Height (cm) | 50.92 ± 2.39 c | 71.25 ± 1.54 a | 55.71 ± 1.15 b | 58.63 ± 4.92 a | 48.82 ± 3.73 b | 61.50 ± 2.18 a |
| Diameter (cm) | 4.04 ± 0.17 a | 4.33 ± 0.31 a | 4.16 ± 0.03 a | 4.00 ± 0.26 a | 3.82 ± 0.24 b | 4.09 ± 0.07 a | |
| Dry wt (g) | 3.56 ± 0.26 b | 7.20 ± 0.89 a | 4.60 ± 0.21 b | 4.39 ± 0.07 b | 4.29 ± 0.07 b | 4.63 ± 0.06 a | |
| Leaf | Width (cm) | 2.53 ± 0.10 b | 2.88 ± 0.14 a | 2.65 ± 0.03 b | 2.10 ± 0.07 a | 2.23 ± 0.06 a | 2.24 ± 0.14 a |
| Length (cm) | 4.22 ± 0.04 ab | 4.48 ± 0.09 b | 4.57 ± 0.25 a | 3.50 ± 0.20 a | 3.53 ± 0.34 a | 3.78 ± 0.24 a | |
| SPAD Value (%) | 17.24 ± 0.42 b | 21.60 ± 0.32 a | 21.25 ± 0.29 a | 23.02 ± 0.21 b | 22.74 ± 0.12 b | 23.58 ± 0.24 a | |
| Nitrogen contents (mg/g) | 1.16 ± 0.02 b | 1.47 ± 0.02 a | 1.45 ± 0.02 a | 1.58 ± 0.02 ab | 1.56 ± 0.01 b | 1.62 ± 0.01 a | |
| Root | Dry wt (g) | 0.13 ± 0.05 a | 0.22 ± 0.13 a | 0.25 ± 0.04 a | 0.15 ± 0.03 c | 0.23 ± 0.01 b | 0.29 ± 0.03 a |
| Flower | Flower diameter (cm) | 10.01 ± 0.02 c | 12.82 ± 0.20 a | 11.89 ± 0.08 b | 10.03 ± 0.19 b | 9.93 ± 0.26 b | 12.36 ± 0.46 a |
| Dry wt (g) | 3.56 ± 0.26 b | 7.20 ± 0.89 a | 4.60 ± 0.21 b | 4.39 ± 0.07 b | 4.29 ± 0.07 b | 4.63 ± 0.06 a | |
| Time | Treatment | F. oxysporum (103 cfu·g−1 soil) | Bacteria (106 cfu·g−1 soil) | Fungi (105 cfu·g−1 soil) | Bacteria/Fungi Ratio |
|---|---|---|---|---|---|
| 2015 | Control | 5.69 ± 0.09 a | 2.66 ± 0.16 a | 12.92 ± 0.25 a | 2.06 ± 0.59 b |
| DZ | 1.49 ± 0.21 c | 2.03 ± 0.18 b | 2.23 ± 0.61 b | 9.10 ± 0.93 a | |
| BF | 2.49 ± 0.02 b | 2.15 ± 0.12 b | 2.96 ± 0.20 b | 7.26 ± 0.76 a | |
| 2016 | Control | 6.61 ± 0.62 a | 2.50 ± 0.21 a | 15.10 ± 0.06 a | 1.65 ± 0.36 b |
| DZ | 6.03 ± 0.06 a | 1.64 ± 0.09 b | 0.61 ± 0.16 b | 1.51 ± 0.21 b | |
| BF | 1.69 ± 0.07 b | 2.56 ± 0.16 a | 0.20 ± 0.54 c | 4.42 ± 0.76 a |
| Soil microbiota | Treatment | 2015 | 2016 | ||||
|---|---|---|---|---|---|---|---|
| H′ | J | S | H′ | J | S | ||
| Bacteria | Control | 3.42 ± 0.05 a | 0.97 ± 0.05 a | 30.33 ± 0.58 a | 3.10 ± 0.01 ab | 0.95 ± 0.04 a | 22.67 ± 0.67 a |
| DZ | 3.12 ± 0.03 b | 0.96 ± 0.18 a | 22.67 ± 0.58 b | 3.06 ± 0.03 b | 0.95 ± 0.14 a | 21.33 ± 0.67 a | |
| BF | 3.12 ± 0.05 b | 0.96 ± 0.05 a | 22.67 ± 1.15 b | 3.14 ± 0.02 a | 0.94 ± 0.07 a | 23.33 ± 0.33 a | |
| Fungi | Control | 2.95 ± 0.02 a | 0.95 ± 0.13 a | 18.67 ± 0.33 a | 2.91 ± 0.05 a | 0.94 ± 0.13 a | 17.33 ± 0.33 a |
| DZ | 2.81 ± 0.02 b | 0.94 ± 0.06 a | 16.67 ± 0.33 b | 2.85 ± 0.04 a | 0.94 ± 0.06 a | 17.67 ± 0.67 a | |
| BF | 2.84 ± 0.03 b | 0.94 ± 0.08 a | 17.33 ± 0.33 b | 2.45 ± 0.05 b | 0.91 ± 0.11 a | 11.67 ± 0.67 b | |
| Years | DGGE Band | Closest Relatives Micro-organisms | Treatment | Similarity (%) | Genebank Accession No. | ||
|---|---|---|---|---|---|---|---|
| Control | DZ | BF | |||||
| 2015 | 1 | Uncultured bacterium | + | + | + | 97 | JN802358.1 |
| 2 | Uncultured Chroococcidiopsis sp. | + | + | + | 99 | LN878320.1 | |
| 3 | Rhizobium sp. | + | − | + | 98 | KP299211.1 | |
| 4 | Halomonas sp. | + | − | − | 95 | KX001824.1 | |
| 5 | Roseospira sp. | - | + | − | 96 | LT158757.1 | |
| 6 | Uncultured sphingobium | - | + | − | 99 | HM371404.1 | |
| 7 | Rhizobium sp. | - | + | − | 100 | EU689093.1 | |
| 8 | Uncultured bacterium | + | + | + | 97 | GQ289422.1 | |
| 9 | Uncultured alpha proteobacterium | - | − | + | 95 | LC017178.1 | |
| 10 | Rhizobiales bacterium | - | − | + | 92 | KU531579.1 | |
| 11 | Uncultured bacterium | - | − | + | 95 | KU928400.1 | |
| 12 | Uncultured bacterium | - | − | + | 97 | JX673167.1 | |
| 13 | Uncultured rhizobiales bacterium | - | − | + | 98 | JF731367.1 | |
| 14 | Mesorhizobium sp. | - | − | + | 94 | DQ659080.1 | |
| 15 | Uncultured bacterium | + | + | + | 98 | KT791490.1 | |
| 16 | Uncultured Mariniflexile sp. | + | + | + | 92 | KM108655.1 | |
| 2016 | 1 | Uncultured Mitsuaria | + | − | − | 97 | KM052508.1 |
| 2 | Halomonas sp. | + | − | − | 97 | KX001824.1 | |
| 3 | Uncultured flexibacter sp. | + | − | − | 98 | EU926966.1 | |
| 4 | Uncultured bacterium | + | − | − | 96 | KY970042.1 | |
| 5 | Uncultured Mariniflexible sp. | + | − | − | 92 | KM108655.1 | |
| 6 | Uncultured Microbulbifer sp. | + | + | + | 99 | LN846107.1 | |
| 7 | Uncultured Agrobacterium sp. | + | + | - | 100 | KU375567.1 | |
| 8 | Uncultured bacterium | + | − | + | 98 | AY649352.1 | |
| 9 | Micromonospora sp. | - | + | − | 98 | KP900783.1 | |
| 10 | Uncultured bacterium | - | + | + | 92 | MF079697.1 | |
| 11 | Uncultured alpha proteobacterium | - | + | + | 97 | LC017178.1 | |
| 12 | Uncultured actinobacterium | - | + | − | 91 | FJ620849.1 | |
| 13 | Uncultured Micromonospora sp. | - | − | + | 92 | KP900783.1 | |
| 14 | Uncultured bacterium | - | − | + | 100 | JF261711.1 | |
| 15 | Uncultured Sporanaerobacter sp. | - | − | + | 100 | KR064311.1 | |
| 16 | Uncultured Rhizobium sp. | - | − | + | 99 | GQ365753.1 | |
| 17 | Uncultured bacterium | - | − | + | 93 | FJ753412.1 | |
| 18 | Uncultured Luteibacter sp. | + | − | + | 98 | JQ027706.1 | |
| Years | DGGE Band | Closest Relatives Microorganisms | Treatment | Similarity (%) | Genebank Accession No. | ||
|---|---|---|---|---|---|---|---|
| Control | DZ | BF | |||||
| 2015 | 1 | Uncultured fungus | + | − | − | 95 | HM246432.1 |
| 2 | Fusarium oxysporum | + | − | − | 98 | KR063173.1 | |
| 3 | Rhizoctonia zeae | + | + | + | 100 | KT347101.1 | |
| 4 | Paraphoma sp. | − | + | − | 94 | LC126020.1 | |
| 5 | Uncultured Orbiliaceae | + | + | + | 96 | KF258906.1 | |
| 6 | Uncultured eukaryote | − | − | + | 97 | KF357450.1 | |
| 7 | Uncultured eukaryote | − | − | + | 97 | AB627976.2 | |
| 8 | Malbranchea cinnamomea | + | − | + | 96 | JX268593.1 | |
| 9 | Monacrosporium sp. | + | + | + | 94 | AJ001998.1 | |
| 10 | Orbilia fimicola | - | − | + | 92 | AF006307.1 | |
| 2016 | 1 | Anguillospora furtiva | + | − | − | 100 | AY357262.1 |
| 2 | Rhizoctonia zeae | + | + | − | 100 | KT347101.1 | |
| 3 | Uncultured Ascomycota | + | + | − | 99 | FJ889079.1 | |
| 4 | Rhizoctonia solani | + | − | − | 98 | JF499071.1 | |
| 5 | Tiarosporella graminis | - | + | − | 99 | KF531827.1 | |
| 6 | Uncultured eukaryote | - | + | − | 96 | KU657636.1 | |
| 7 | Deconica sp. | - | + | − | 99 | KJ137262.1 | |
| 8 | Uncultured fungus | - | + | + | 98 | KJ755398.1 | |
| 9 | Uncultured eukaryote | + | + | + | 99 | KF357450.1 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Zhao, S.; Zhang, K.; Zhao, J.; Jiang, J.; Chen, F.; Fang, W. Evaluation of Soil-Applied Chemical Fungicide and Biofungicide for Control of the Fusarium Wilt of Chrysanthemum and Their Effects on Rhizosphere Soil Microbiota. Agriculture 2018, 8, 184. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture8120184
Chen H, Zhao S, Zhang K, Zhao J, Jiang J, Chen F, Fang W. Evaluation of Soil-Applied Chemical Fungicide and Biofungicide for Control of the Fusarium Wilt of Chrysanthemum and Their Effects on Rhizosphere Soil Microbiota. Agriculture. 2018; 8(12):184. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture8120184
Chicago/Turabian StyleChen, Huijie, Shuang Zhao, Kaikai Zhang, Jiamiao Zhao, Jing Jiang, Fadi Chen, and Weimin Fang. 2018. "Evaluation of Soil-Applied Chemical Fungicide and Biofungicide for Control of the Fusarium Wilt of Chrysanthemum and Their Effects on Rhizosphere Soil Microbiota" Agriculture 8, no. 12: 184. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture8120184