Determination of Insecticide Susceptibility of Field Populations of Tomato Leaf Miner (Tuta absoluta) in Northern Nigeria


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
Chemicals and Reagents
2.2. Methods
2.2.1. Field Collection and Rearing of Insects
2.2.2. Morphological Identification of T. absoluta Life Stages
2.2.3. Molecular Identification of T. absoluta to Species Level
2.2.4. Insecticides (Leaf-Dip) Bioassay
2.2.5. Synergists Bioassay
2.2.6. Sequencing of Domain II of the Voltage-Gated Sodium Channel
2.3. Data Analysis
3. Results
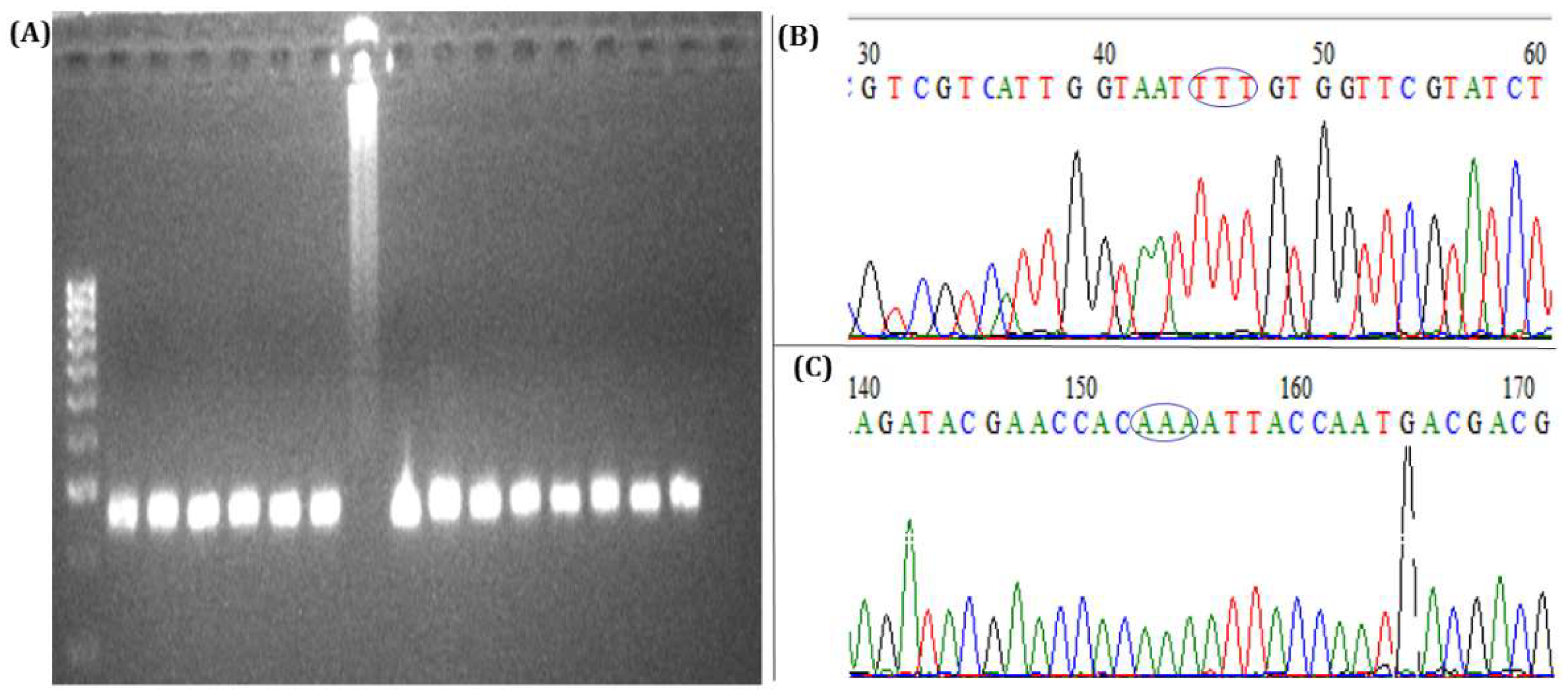
3.1. Morphological and Molecular Identification of T. absoluta to Species Level
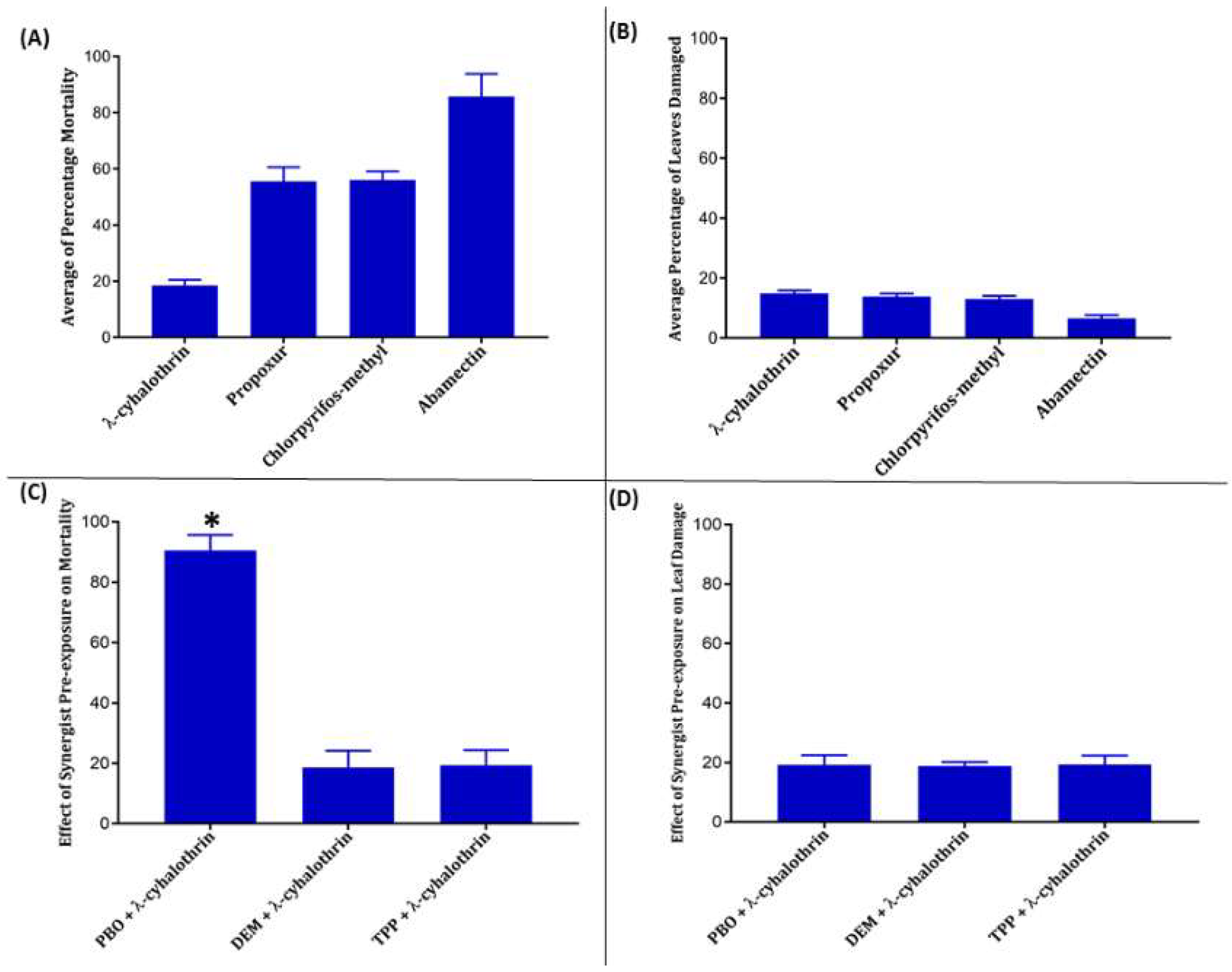
3.2. Insecticides Resistance Profile of T. absoluta Populations
3.3. Identification of Possible Mechanism of Resistance Using Synergists
3.4. Identification of 1014F kdr Mutation in Domain II of the Voltage-Gated Sodium Channel
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- García, M.F.; Espul, J.C. Bioecology of the tomato moth (Scrobipalpula absoluta) in Mendoza, Argentine Republic. Revista de Investigaciones Agropecuarias 1982, 17, 135–146. [Google Scholar]
- European and Mediterranean Plant Protection Organisation. Data sheets on quarantine pests. EPPO Bull. 2005, 35, 434–435. [Google Scholar] [CrossRef]
- García, A.U.; Vercher, R.; Llopis, V.N.; Porcuna y Col, J.L.; Marí, F.G. La polilla del tomate, “Tuta absoluta”. Phytoma Espana: La Revista Profesional de Sanidad Vegetal 2007, 194, 16–23. [Google Scholar]
- CABI. Invasive Species Compendium. Detailed Coverage of Invasive Species Threatening Livelihoods and the Environment Worldwide. Available online: https://www.cabi.org/ISC/tuta (accessed on 24 October 2018).
- Tropea, G.G.; Siscaro, G.; Biondi, A.; Zappalà, L. Tuta absoluta, a South American pest of tomato now in the EPPO region: Biology, distribution and damage. EPPO Bull. 2012, 42, 205–210. [Google Scholar] [CrossRef]
- Desneux, N.; Luna, M.G.; Guillemaud, T.; Urbaneja, A. The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: The new threat to tomato world production. J. Pest Sci. 2011, 84, 403–408. [Google Scholar] [CrossRef]
- Nayana, B.P.; Kalleshwaraswamy, C.M. Biology and external morphology of invasive tomato leaf miner, Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae). Pest Manag. Hortic. Ecosyst. 2015, 21, 169–174. [Google Scholar]
- Baniameri, V.; Cheraghian, A. The first report and control strategies of Tuta absoluta in Iran. EPPO Bull. 2012, 42, 322–324. [Google Scholar] [CrossRef]
- Gashawbeza, A.; Abiy, F. Occurrence of a new leaf mining and fruit boring moth of tomato (Tuta absoluta) in Ethiopia. Field Trip Rep. Ethiop. Inst. Agric. Res. 2013, 11, 2013. [Google Scholar]
- Haougui, A.; Adamou, B.; Garba, M.; Oumarou, S.; Gougari, B.; Abou, M.; Kimba, A.; Delmas, P. Confirmation of the presence of Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae) in Niger (West Africa). J. Sci. Environ. 2016, 5, 4481–4486. [Google Scholar]
- Pfeiffer, D.G.; Muniappan, R.; Sall, D.; Diatta, P.; Diongue, A.; Dieng, E.O. First record of Tuta absoluta (Lepidoptera: Gelechiidae) in Senegal. Fla. Entomol. 2013, 96, 661–662. [Google Scholar] [CrossRef]
- Maneno, C.; Al-zaidi, S.; Hassan, N.; Julie, A.; Kaaya, E.; Mrogoro, S. First record of tomato leaf miner Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) in Tanzania. Agric. Food Secur. 2016, 5, 17. [Google Scholar]
- Mutamiswa, R.; Machekano, H.; Nyamukondiwa, C. First report of tomato leaf miner, Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae), in Botswana. Agric. Food Secur. 2017, 6, 49. [Google Scholar] [CrossRef]
- NEPAD: Comprehensive Africa Agriculture Development Programme (CAADP). 2003. Available online: http://www.fao.org/nr/water/aquastat/sirte2008/NEPAD-CAADP%202003.pdf (accessed on 29 July 2018).
- FAOSTAT: CROPS Statistics for Countries. United Nations, NY. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 5 November 2018).
- Borisade, O.A.; Kolawole, A.O.; Adebo, G.M.; Uwaidem, Y.I. The tomato leafminer (Tuta absoluta)(Lepidoptera: Gelechiidae) attack in Nigeria: Effect of climate change on over-sighted pest or agro-bioterrorism? J. Agric. Ext. Rural Dev. 2017, 9, 163–171. [Google Scholar] [CrossRef]
- Sanda, N.B.; Sunusi, M.; Hamisu, H.; Wudil, B.; Sule, H.; Abdullahi, A. Biological Invasion of Tomato Leaf Miner, Tuta absoluta (Meyrick) in Nigeria: Problems and Management Strategies Optimization: A Review. Asian J. Agric. Hortic. Res. 2018, 1, 1–14. [Google Scholar] [CrossRef]
- Punch Newspaper Online. Tomato farmers in Kano lose N2bn to Tuta Absoluta. Punch Newspaper. Available online: https://punchng.com/tomato-farmers-in-kano-lose-n2bn-to-tuta-absoluta/ (accessed on 5 November 2018).
- Haddi, K.; Berger, M.; Bielza, P.; Rapisarda, C.; Williamson, M.; Moores, G.; Bass, C. Mutation in the ace-1 gene of the tomato leaf miner (Tuta absoluta) associated with organophosphates resistance. J. Appl. Entomol. 2017, 141, 612–619. [Google Scholar] [CrossRef]
- Roditakis, E.; Vasakis, E.; García-Vidal, L.; María del Rosario, M.; Rison, J.L.; Haxaire-Lutun, M.O.; Nauen, R.; Tsagkarakou, A.; Bielza, P. A four-year survey on insecticide resistance and likelihood of chemical control failure for tomato leaf miner Tuta absoluta in the European/Asian region. J. Pest Sci. 2018, 91, 421–435. [Google Scholar] [CrossRef]
- Salazar, E.R.; Araya, J.E. Respuesta de la polilla del tomate, Tuta absoluta (Meyrick), a insecticidas en Arica. Agric. Téc. 2001, 61, 429–435. [Google Scholar] [CrossRef]
- Silva, W.M.; Berger, M.; Bass, C.; Balbino, V.Q.; Amaral, M.H.; Campos, M.R.; Siqueira, H.A. Status of pyrethroid resistance and mechanisms in Brazilian populations of Tuta absoluta. Pestic. Biochem. Physiol. 2015, 122, 8–14. [Google Scholar] [CrossRef]
- Roditakis, E.; Skarmoutsou, C.; Staurakaki, M.; María del Rosario, M.; García-Vidal, L.; Bielza, P.; Haddi, K.; Rapisarda, C.; Rison, J.L.; Bassi, A.; et al. Determination of baseline susceptibility of European populations of Tuta absoluta (Meyrick) to indoxacarb and chlorantraniliprole using a novel dip bioassay method. Pest Manag. Sci. 2013, 69, 217–227. [Google Scholar] [CrossRef]
- Haddi, K.; Berger, M.; Bielza, P.; Cifuentes, D.; Field, L.M.; Gorman, K.; Rapisarda, C.; Williamson, M.S.; Bass, C. Identification of mutations associated with pyrethroid resistance in the voltage-gated sodium channel of the tomato leaf miner (Tuta absoluta). Insect Biochem. Mol. Biol. 2012, 42, 506–513. [Google Scholar] [CrossRef]
- Ayalew, G. Efficacy of selected insecticides against the South American tomato moth, Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) on tomato in the Central Rift Valley of Ethiopia. Afr. Entomol. 2015, 23, 410–417. [Google Scholar] [CrossRef]
- Shiberu, T.; Getu, E. Evaluation of Some Insecticides against Tomato Leaf Miner, Tuta absoluta (Meyrick) (Gelechiidae: Lepidoptera) Under Laboratory and Glasshouse Conditions. Agric. Res. Technol. 2017, 7. [Google Scholar] [CrossRef]
- Ranson, H.; Abdallah, H.; Badolo, A.; Guelbeogo, W.M.; Kerah-Hinzoumbe, C.; Yangalbe-Kalnone, E.; Sagnon, N.; Simard, F.; Coetzee, M. Insecticide resistance in Anopheles gambiae: Data from the first year of a multi-country study highlight the extent of the problem. Malar. J. 2009, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Bajonero, J.G.; Parra, J.R.P. Selection and suitability of an artificial diet for Tuta absoluta (Lepidoptera: Gelechiidae) based on physical and chemical characteristics. J. Insect Sci. 2017, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J. Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics 1984, 107, 611–634. [Google Scholar]
- Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Tonnang, H.E.; Mohamed, S.F.; Khamis, F.; Ekesi, S. Identification and risk assessment for worldwide invasion and spread of Tuta absoluta with a focus on Sub-Saharan Africa: Implications for phytosanitary measures and management. PLoS ONE 2015, 10, e0135283. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- IRAC. Tuta absoluta Susceptibility Test Method #022. Available online: http://www.irac-online.org/methods/tuta-absoluta-larvae/ (accessed on 4 March 2017).
- Nwane, P.; Etang, J.; Chouaїbou, M.; Toto, J.C.; Koffi, A.; Mimpfoundi, R.; Simard, F. Multiple insecticide resistance mechanisms in Anopheles gambiae sl populations from Cameroon, Central Africa. Parasites Vector 2013, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.M.; Raghavendra, K.; Verma, V.; Velamuri, P.S.; Pande, V. Esterases are responsible for malathion resistance in Anopheles stephensi: A proof using biochemical and insecticide inhibition studies. J. Vector Borne Dis. 2017, 54, 226. [Google Scholar]
- Sridhar, V.; Chakravarthy, A.; Asokan, R. New record of the invasive South American tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) in India. Pest Manag. Hortic. Ecosyst. 2014, 20, 148–154. [Google Scholar]
- Hebert, P.D.; Cywinska, A.; Ball, S.L. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sint, D.; Sporleder, M.; Wallinger, C.; Zegarra, O.; Oehm, J.; Dangi, N.; Giri, Y.P.; Kroschel, J.; Traugott, M. A two-dimensional pooling approach towards efficient detection of parasitoid and pathogen DNA at low infestation rates. Methods Ecol. Evol. 2016, 7, 1548–1557. [Google Scholar] [CrossRef]
- Davies, T.; Williamson, M. Interactions of pyrethroids with the voltage-gated sodium channel. Bayer CropSci. J. 2009, 62, 159–178. [Google Scholar]
- Son, D.; Bonzi, S.; Somda, I.; Bawin, T.; Boukraa, S.; Verheggen, F.; Francis, F.; Legrève, A.; Schiffers, B. First Record of Tuta absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae) in Burkina Faso. Afr. Entomol. 2017, 25, 259–263. [Google Scholar] [CrossRef]
- Chidege, M.; Abel, J.; Afonso, Z.; Tonini, M.; Fernandez, B. Tomato Leaf Miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) Detected in Namibe Province Angola. J. Appl. Life Sci. Int. 2017, 12, 1–5. [Google Scholar] [CrossRef]
- Zibaee, I.; Mahmood, K.; Esmaeily, M.; Bandani, A.; Kristensen, M. Organophosphate and pyrethroid resistances in the tomato leaf miner Tuta absoluta (Lepidoptera: Gelechiidae) from Iran. J. Appl. Entomol. 2018, 142, 181–191. [Google Scholar] [CrossRef]
- Siqueira, H.A.; Guedes, R.N.; Picanço, M.C. Insecticide resistance in populations of Tuta absoluta (Lepidoptera: Gelechiidae). Agric. For. Entomol. 2000, 2, 147–153. [Google Scholar] [CrossRef]
- Lietti, M.M.; Botto, E.; Alzogaray, R.A. Insecticide resistance in Argentine populations of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Néotrop. Entomol. 2005, 34, 113–119. [Google Scholar] [CrossRef]
- Siqueira, H.A.; Guedes, R.; Fragoso, D.B.; Magalhaes, L. Abamectin resistance and synergism in Brazilian populations of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Int. J. Pest Manag. 2001, 47, 247–251. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (ppm) | Population before Treatment | Population after Treatment | Mortality (%) | Leaf Damage (%) |
|---|---|---|---|---|
| Abamectin | ||||
| 1000 | 9 | 0 (9 dead) | 100 | 0.22 |
| 166.67 | 9 | 0 (9 dead) | 100 | 2.17 |
| 27.78 | 9 | 0 (9 dead) | 100 | 4.78 |
| 4.630 | 9 | 0 (9 dead) | 100 | 6.00 |
| 0.7716 | 9 | 0 (9 dead) | 100 | 7.22 |
| 0.1286 | 9 | 1 alive (8 dead) | 85.71 | 6.78 |
| 0.0214 | 9 | 4 alive (5 dead) | 42.86 | 12.00 |
| 0.0036 | 9 | 3 alive (6 dead) | 57.14 | 13.44 |
| 0.00 (control) | 9 | 9 alive (0 dead) | 0.00 | 16.61 |
| Propoxur | ||||
| 5000 | 9 | 2 alive (7 dead) | 77.77 | 5.78 |
| 833.33 | 9 | 3 alive (6 dead) | 66.66 | 8.67 |
| 138.89 | 9 | 4 alive (5 dead) | 55.55 | 9.67 |
| 23.15 | 9 | 4 alive (5 dead) | 55.55 | 14.22 |
| 3.858 | 9 | 5 alive (4 dead) | 44.44 | 19.78 |
| 0.643 | 9 | 6 alive (3 dead) | 33.33 | 20.22 |
| 0.00 (control) | 9 | 9 alive (0 dead) | 0.00 | 25.56 |
| λ-cyhalothrin | ||||
| 5000 | 9 | 6 alive (3 dead) | 33.33 | 7.89 |
| 833.33 | 9 | 7 alive (2 dead) | 22.22 | 9.17 |
| 138.89 | 9 | 8 alive (1 dead) | 11.11 | 13.33 |
| 23.15 | 9 | 7 alive (2 dead) | 22.22 | 16.89 |
| 3.858 | 9 | 8 alive (1 dead) | 11.11 | 19.22 |
| 0.643 | 9 | 8 alive (1dead) | 11.11 | 22.78 |
| 0.00 (control) | 9 | 9 alive (0 dead) | 0.00 | 25.56 |
| Chlorpyrifos-Methyl | ||||
| 5000 | 9 | 0 alive (9 dead) | 100 | 4.94 |
| 833.33 | 9 | 0 alive (9 dead) | 100 | 8.72 |
| 138.89 | 9 | 4 alive (5 dead) | 50 | 17.78 |
| 23.15 | 9 | 5 alive (4 dead) | 37.5 | 17.67 |
| 3.858 | 9 | 5 alive (4 dead) | 37.5 | 16.11 |
| 0.643 | 9 | 8 alive (1 dead) | 11.11 | 17.78 |
| 0.00 (control) | 9 | 9 alive (0 dead) | 11.11 | 20.44 |
| Piperonyl Butoxide (PBO) + λ-cyhalothrin | ||||
| 138.89 | 9 | 0 alive (9 dead) | 100 | 1 |
| 23.15 | 9 | 0 alive (9 dead) | 100 | 1 |
| 3.858 | 9 | 1 alive (6 dead) | 87.50 | 3 |
| 0.643 | 9 | 2 alive (7 dead) | 75.00 | 5 |
| 0.00 (control) | 27 | 27alive (0 dead) | 0.00 | - |
| Diethyl Maleate (DEM) + λ-cyhalothrin | ||||
| 138.89 | 9 | 7 alive (2 dead) | 22.22 | 13.33 |
| 23.15 | 9 | 8 alive (1 dead) | 11.11 | 16.89 |
| 3.858 | 9 | 8 alive (1 dead) | 11.11 | 19.22 |
| 0.643 | 9 | 8 alive (1 dead) | 11.11 | 22.75 |
| Triphenyl Phosphate (TPP) + λ-cyhalothrin | ||||
| 138.89 | 9 | 7 alive (2 dead) | 22.22 | 12.66 |
| 23.15 | 9 | 7 alive (2 dead) | 22.22 | 17.47 |
| 3.858 | 9 | 7 alive (2 dead) | 22.22 | 23.10 |
| 0.643 | 9 | 8 alive (1 dead) | 11.11 | 20.85 |
| Insecticides | n | Leaf Damage ± S.E.M. | % Mortality ± S.E.M. | LD50 (ppm) | LD50 (S.E.M) | LD50 (FL 95%) |
|---|---|---|---|---|---|---|
| λ-cyhalothrin | 54 | 14.88 ± 1.05 | 18.51 ± 5.70 | 7461.47 | 1213.79 | 5082.47–9840.89 |
| Propoxur | 54 | 13.83 ± 1.00 | 55.55 ± 5.02 | 1023.35 | 218.68 | 594.08–1452.04 |
| Chlorpyrifos-methyl | 54 | 13.06 ± 1.06 | 56.01 ± 3.10 | 106.30 | 13.09 | 80.75–132.15 |
| Abamectin | 72 | 6.58 ± 1.10 | 85.71 ± 8.10 | 0.034 | 0.0036 | 0.01–0.09 |
| Control | 36 | 22.04 ± 3.05 | 0.00 ± 0.00 | ND | ND | ND |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bala, I.; Mukhtar, M.M.; Saka, H.K.; Abdullahi, N.; Ibrahim, S.S. Determination of Insecticide Susceptibility of Field Populations of Tomato Leaf Miner (Tuta absoluta) in Northern Nigeria. Agriculture 2019, 9, 7. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9010007
Bala I, Mukhtar MM, Saka HK, Abdullahi N, Ibrahim SS. Determination of Insecticide Susceptibility of Field Populations of Tomato Leaf Miner (Tuta absoluta) in Northern Nigeria. Agriculture. 2019; 9(1):7. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9010007
Chicago/Turabian StyleBala, Ibrahim, Muhammad M. Mukhtar, Habeeb K. Saka, Nasiru Abdullahi, and Sulaiman S. Ibrahim. 2019. "Determination of Insecticide Susceptibility of Field Populations of Tomato Leaf Miner (Tuta absoluta) in Northern Nigeria" Agriculture 9, no. 1: 7. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9010007