Effects of Soil pH and Fertilizers on Haskap (Lonicera caerulea L.) Vegetative Growth

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
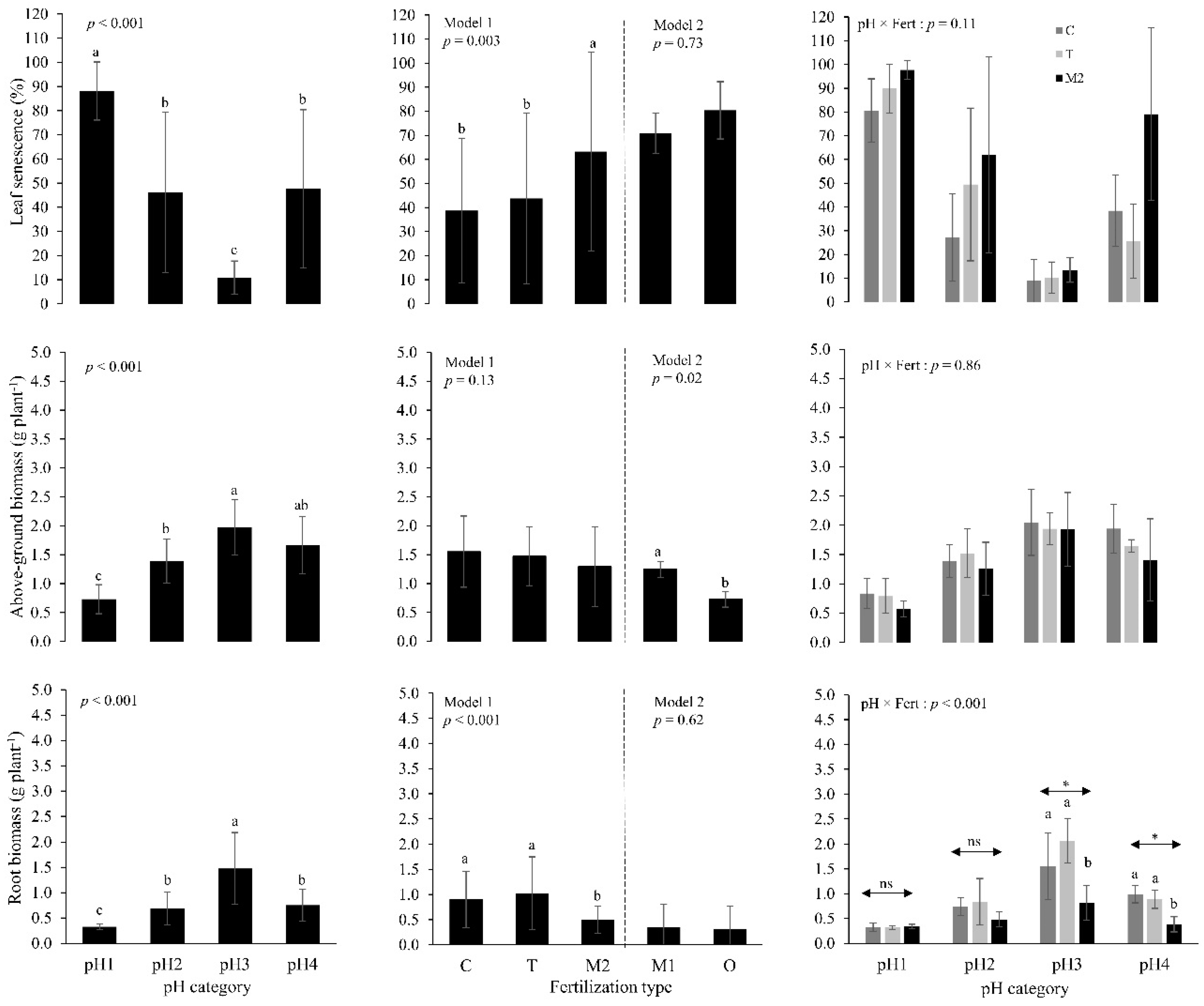
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vasantha Rupasinghe, H.; Yu, L.; Bhullar, K.; Bors, B. Haskap (Lonicera caerulea): A new berry crop with high antioxidant capacity. Can. J. Plant Sci. 2012, 92, 1311–1317. [Google Scholar] [CrossRef]
- Takahashi, A.; Okazaki, Y.; Nakamoto, A.; Watanabe, S.; Sakaguchi, H.; Tagashira, Y.; Kagii, A.; Nakagawara, S.; Higuchi, O.; Suzuki, T.; et al. Dietary Anthocyanin-rich Haskap Phytochemicals Inhibit Postprandial Hyperlipidemia and Hyperglycemia in Rats. J. Oleo Sci. 2014, 63, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummer, K.E. Blue honeysuckle: A new berry crop for North America. J. Am. Pomol. Soc. 2006, 60, 3–8. [Google Scholar]
- Lamoureux, D.; Sorokin, A.; Lefèvre, I.; Alexanian, S.; Eyzaguirre, P.; Hausman, J.F. Investigation of genetic diversity in Russian collections of raspberry and blue honeysuckle. Plant Genet. Resour. Charact. Util. 2011, 9, 202–205. [Google Scholar] [CrossRef]
- Thompson, M.M.; Barney, D.L. Evaluation and breeding of haskap in North America. J. Am. Pomol. Soc. 2007, 61, 25–33. [Google Scholar]
- Auzanneau, N.; Weber, P.; Kosińska-Cagnazzo, A.; Andlauer, W. Bioactive compounds and antioxidant capacity of Lonicera caerulea berries: Comparison of seven cultivars over three harvesting years. J. Food Compos. Anal. 2018, 66, 81–89. [Google Scholar] [CrossRef]
- Martel, P.-O. Faits saillants sur la camerise, Rencontre des intervenants des petits fruits émergents; Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ): Québec, QC, Canada, 2018.
- Gerbrandt, E. Propagation, management and adaptation of the blue honeysuckle. In Proceedings of the 2014 Annual Meeting of the International Plant Propagators Society, Bellefonte, PA, USA, 27 October 2014; Heuser, C.W., Ed.; International Society for Horticultural Science: Leuven, Belgium, 2015; Volume 1085, pp. 289–292. [Google Scholar]
- Gagnon, A. La Camerise: Guide de Production; Publication 15-0031; Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ): Québec, QC, Canada, 2015.
- Gerbrandt, E.M.; Bors, R.H.; Chibbar, R.N.; Baumann, T.E. Blue honeysuckle (Lonicera caerulea L.) vegetative growth cessation and leaf drop phenological adaptation to a temperate climate. Genet. Resour. Crop Evol. 2018, 65, 1471–1484. [Google Scholar] [CrossRef]
- Gerbrandt, E.M.; Bors, R.H.; Chibbar, R.N.; Baumann, T.E. Spring phenological adaptation of improved blue honeysuckle (Lonicera caerulea L.) germplasm to a temperate climate. Euphytica 2017, 213, 172. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Raymond, R.; Mailloux, A.; Dubé, A. Pédologie de la région du Lac-Saint-Jean; Bulletin technique no 11; Ministère de l’Agriculture et de la Colonisation du Québec: Québec, QC, Canada, 1965; p. 159. [Google Scholar]
- Cheng, Y.; Wang, J.; Mary, B.; Zhang, J.-b.; Cai, Z.-C.; Chang, S.X. Soil pH has contrasting effects on gross and net nitrogen mineralizations in adjacent forest and grassland soils in central Alberta, Canada. Soil Biol. Biochem. 2013, 57, 848–857. [Google Scholar] [CrossRef]
- Hendershot, W.H.; Lalande, H.; Duquette, M. Soil reaction and exchangeable acidity. In Soil Sampling and Methods of Analysis; Carter, M.R., Gregorich, E.G., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 173–178. [Google Scholar]
- Brennan, R.F.; Bolland, M.D.A. Relationship between pH measured in water and calcium chloride for soils of Southwestern Australia. Commun. Soil Sci. Plan. 1998, 29, 2683–2689. [Google Scholar] [CrossRef]
- Van Lierop, W. Conversion of organic soil pH values measured in water, 0.01M CaCl2 or 1N KCl. Can. J. Soil Sci. 1981, 61, 577–579. [Google Scholar] [CrossRef]
- Vanasse, A.; Hébert, M.; Khiari, L.; Marchand, S.; Badra, A.; Moore, H. La gestion du pH du sol. In Guide de référence en fertilisation, 2nd ed.; Parent, L.-E., Gagné, G., Eds.; Commission Chimie et Fertilité des sols, Centre de Référence en Agriculture et Agroalimentaire du Québec (CRAAQ): Québec, QC, Canada, 2010; pp. 71–100. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Islam, A.K.M.S.; Edwards, D.G.; Asher, C.J. pH optima for crop growth. Plant Soil 1980, 54, 339–357. [Google Scholar] [CrossRef]
- Walter, A.; Silk, W.K.; Schurr, U. Effect of soil pH on growth and cation deposition in the root tip of Zea mays L. J. Plant Growth Regul. 2000, 19, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Brunner, I.; Brodbeck, S.; Walthert, L. Fine root chemistry, starch concentration, and ‘vitality’ of subalpine conifer forests in relation to soil pH. Forest Ecol. Manag. 2002, 165, 75–84. [Google Scholar] [CrossRef]
- Hamid, F.S.; Ahmad, T.; Khan, B.M.; Waheed, A.; Ahmed, N. Effect of soil pH in rooting and growth of tea cuttings (Camellia sinensis L.) at nursery level. Pak. J. Bot. 2006, 38, 293–300. [Google Scholar]
- Abdulaha-Al Baquy, M.; Li, J.Y.; Xu, C.Y.; Mehmood, K.; Xu, R.K. Determination of critical pH and Al concentration of acidic Ultisols for wheat and canola crops. Solid Earth 2017, 8, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.Y.; Gerendas, J.; Hardter, R.; Sattelmacher, B. Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis) plants. Ann. Bot. 2007, 99, 301–310. [Google Scholar] [CrossRef]
- Kerley, S.J. The effect of soil liming on shoot development, root growth, and cluster root activity of white lupin. Biol. Fertil. Soils 2000, 32, 94–101. [Google Scholar] [CrossRef]
- Bernstein, L. Effects of salinity and sodicity on plant growth. Annu. Rev. Phytopathol. 1975, 13, 295–312. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Jan, A.; Osman, M.B.; Amanullah. Response of chickpea to nitrogen sources under salinity stress. J. Plant Nutr. 2013, 36, 1373–1382. [Google Scholar] [CrossRef]
- Farooq, N.; Kanwal, S.; Ditta, A.; Hussain, A.; Naveed, M.; Jamshaid, M.U.; Iqbal, M. Comparative efficacy of KCl blended composts and sole application of KCl or K2SO4 in improving K nutrition, photosynthetic capacity and growth of maize. Soil Environ. 2018, 37, 68–74. [Google Scholar] [CrossRef]
- Zhang, H.; Rong, H.; Pilbeam, D. Signalling mechanisms underlying the morphological responses of the root system to nitrogen in Arabidopsis thaliana. J. Exp. Bot. 2007, 58, 2329–2338. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D. The responses of plants to nonuniform supplies of nutrients. New Phytol. 1994, 127, 635–674. [Google Scholar] [CrossRef]


{kind=link}
| Soil Properties | Method | Value |
|---|---|---|
| Soil pH | In water (1:1) | 6.4 |
| Soil organic matter (%) | Combustion | 5.4 |
| Total nitrogen (g·kg−1) | Kjeldahl | 1.4 |
| P (mg·kg−1) | Mehlich 3-Extractable | 5.4 |
| K (mg·kg−1) | Mehlich 3-Extractable | 53.1 |
| Mg (mg·kg−1) | Mehlich 3-Extractable | 17.3 |
| Ca (mg·kg−1) | Mehlich 3-Extractable | 1284 |
| Al (mg·kg−1) | Mehlich 3-Extractable | 2097 |
| Fe (mg·kg−1) | Mehlich 3-Extractable | 67.8 |
| Factor | pH Category | Soil pH Class 1 | pHCaCl2 | pHwater 1 | |
|---|---|---|---|---|---|
| Primary factor | pH1 | Strongly acidic | 4.3–4.6 | 4.7–5.0 | |
| pH2 | Moderately acidic | 5.1–5.4 | 5.5–5.8 | ||
| pH3 | Slightly acidic | 5.5–6 | 5.9–6.5 | ||
| pH4 | Neutral | 6.3–7 | 6.8–7.5 | ||
| Fertilizer type | N–P–K 2 | N | P | K | |
| g plant−1 | Source | ||||
| Secondary factor | C | 0–0–0 | --- | --- | --- |
| T | 0–2.9–1.7 | --- | Ca(H2PO4)2 | KCl | |
| M1 | 4–2.9–1.7 | (NH4)2SO4 | Ca(H2PO4)2 | KCl | |
| M2 | 4–2.9–1.7 | Ca(NO3)2 | Ca(H2PO4)2 | KCl | |
| O | 4–2.9–1.7 | Granulated poultry manure | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tremblay, C.; Deslauriers, A.; Lafond, J.; Lajeunesse, J.; Paré, M.C. Effects of Soil pH and Fertilizers on Haskap (Lonicera caerulea L.) Vegetative Growth. Agriculture 2019, 9, 56. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9030056
Tremblay C, Deslauriers A, Lafond J, Lajeunesse J, Paré MC. Effects of Soil pH and Fertilizers on Haskap (Lonicera caerulea L.) Vegetative Growth. Agriculture. 2019; 9(3):56. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9030056
Chicago/Turabian StyleTremblay, Catherine, Annie Deslauriers, Jean Lafond, Julie Lajeunesse, and Maxime C. Paré. 2019. "Effects of Soil pH and Fertilizers on Haskap (Lonicera caerulea L.) Vegetative Growth" Agriculture 9, no. 3: 56. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9030056