
Seaweed Beds and Community Structure in the East and South Coast of Korea


1
Department of Ecological Engineering, Pukyong National University, Busan 48513, Korea
2
East Sea Environment Research Center, East Sea Research Institute, Korea Institute of Ocean Science and Technology, Uljin 36315, Korea
*
Author to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2022, 10(5), 689; https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050689
Submission received: 20 April 2022
/
Revised: 11 May 2022
/
Accepted: 12 May 2022
/
Published: 19 May 2022
(This article belongs to the Section Marine Ecology)
Abstract
:This study conducted a community investigation via scuba-diving excursions into the subtidal regions of seven sea areas on the eastern coasts and three sea areas on the southern coasts, from October to December 2017, to determine the characteristics of seaweed communities and the current status of barren ground in natural seaweed beds in Korea. The results showed that species composition and average biomass in the sea area were 5–48 species and an average of 114.42 g/m2 (0.29–273.60 g/m2) in the eastern coasts, where red algae—an annual opportunity species—were dominant, and 21–48 species and an average of 1056.84 g/m2 (53.03–2683.02 g/m2) in the southern coasts, where perennial large brown algae were dominant. Using Orfanidis’ EEI-c model, evaluations of the community states showed they varied significantly depending on the inclusion of melobesidean algae, and this model was determined to be inappropriate for direct application in sea areas with a low coverage of all macroalgae. A comprehensive review of the seaweed community characteristics of seaweed beds, the marine environment, the coverage of melobesidean algae, and the analysis results regarding the density of grazers showed that a decrease in the seaweed community, according to the barren ground phenomenon, was more severe in the eastern than southern coasts. Furthermore, there were also significant differences in seaweed community characteristics according to sea area and barren ground. Therefore, suitable countermeasures corresponding to the characteristics of each sea area are necessary; for example, the creation of growth substrates for the colonization of macroalgae in Deoksin and Saido Is., the transplantation of large brown algae in Gangyang and Daedurado Is., action plans for marine forest monitoring in Geomundo Is., and an improvement in substrates in Yeongjin, Mangsang, Daejin, Chogok, and Geundeok are possible countermeasures.
1. Introduction
Natural seaweed beds, also referred to as “sea forest,” “kelp forest,” and “marine forest,” refer to communities of macroalgae that have been naturally formed [1,2,3]. Seaweed beds greatly contribute to coastal ecosystems as a habitat, spawning place, and food source for various marine organisms. However, the distribution of macroalgae communities around the world has recently been decreasing due to various causes, including climate change and ecological environment change [4,5].
In Korea, domestic barren ground phenomena started locally, occurring in the 1960s and 1970s [6,7], and the area of occurrence is continuously expanding. Since the report of a spreading area of 370 ha in 1997, it has expanded to 2413 ha in 2003–2004 and 10,518 ha in 2014 in the eastern coast. As a result, barren ground is becoming an increasingly serious problem, occurring across 62% of the entire rock mass area of 170,054 ha [8]. In the southern coasts, barren ground in the Yeosu sea area was reportedly 70 ha in 2006 [9], expanding to 962 ha across the entire southern coast in 2015, indicating that 30% of the entire rock mass area of 8234 ha is barren ground [10]. Thus, the spread of barren ground is expected to accelerate in both the eastern and southern coasts. This will result in increased damage, such as the destruction of coastal ecosystems and a reduction in fishery resources.
Ample quantitative research on the seaweed community in the eastern and southern coasts has been performed. This body of research began with Lee and Kang; Lee [11,12] studied the seaweed flora of Dongbaeksum, and this research continued with recent studies by Choi and Rho, Choi et al., Kim et al., Jeong et al., Kwon and Choi, Shin et al., and Han et al. [13,14,15,16,17,18,19,20], in the case of the eastern coasts. Studies on the southern coasts began with research by Song et al. [21], on the seaweed flora of Odongdo, and extends to recent studies by Park et al., Song et al., Oh et al., and Heo et al. [22,23,24,25,26]. However, most of these studies addressed seasonal changes in communities across certain sea areas, which is insufficient for handling the current status of barren ground phenomena.
Therefore, this study proposes a method for monitoring and managing seaweed beds suitable for the specific characteristics of seaweed communities in each sea area. This method was created by analyzing the structure of seaweed communities, evaluating the conditions of the communities, and identifying the current status of seaweed communities in natural seaweed beds among the seaweed habitats distributed in the eastern and southern coasts of Korea.
2. Materials and Methods
2.1. Marine Environment Survey
Marine environment data gathered from the nearby Marine Environmental Monitoring System were utilized for all sea areas except Geomundo Is. Since there are no Marine Environmental Monitoring System data on Geomundo Is., data, such as water temperatures, were collected at sites using a comprehensive water-quality measuring device (JFE-ADVANTECH, AAQ-RINKO, Victoria, Canada). Water temperature, salinity, pH, dissolved oxygen (DO), Chl-a, dissolved inorganic nitrogen (DIN), and dissolved inorganic phosphate (DIP) were analyzed according to the maritime environment pollutant testing method after transporting the water collected from the site in a refrigerator.
2.2. Collection and Analysis
Macroalgae were collected from October–December 2017 from the eastern coasts, including Yeongjin, Mangsang, Daejin, Geundeok, Chogok, Deoksin, and Gangyang, and southern coasts, including Saido Is., Daedurado Is., and Geomundo Is., to identify the characteristics of natural seaweed beds and the current status of barren ground distributed across 10 sea areas. After selecting stations representative of seaweed communities within a depth of 7–12 m for each sea area, qualitative and quantitative surveys were conducted through underwater scuba diving (Table 1). A quantitative survey was performed after taking photographs using a 50 × 50 cm quadrat, which was subdivided into 10 × 10 cm portions. The macroalgae were collected entirely using a scraper, and the coverage was simultaneously recorded at the sites. A 1 × 1 m quadrat was further utilized to record the density of grazers (sea urchins, turban shell, abalone, and aplysia) at the sites.
After the collected macroalgae were placed in an icebox at the site and transported to the laboratory, biological identification was performed with the naked eye and a microscope (Olympus SZX9, Olympus BX50, Tokyo, Japan). The wet weights of the quantitatively collected samples were measured with an accuracy up to 0.01 g for each species using an electronic balance (CAS, CBL3200H, Seoul, Korea). The names of each species and its list were identified following the method provided by Kim et al. [27]. The quantitative samples of macroalgae and the density of grazers were converted into biomass per unit area (g wet wt./m2) and number of individuals per unit area (ind./m2), respectively.
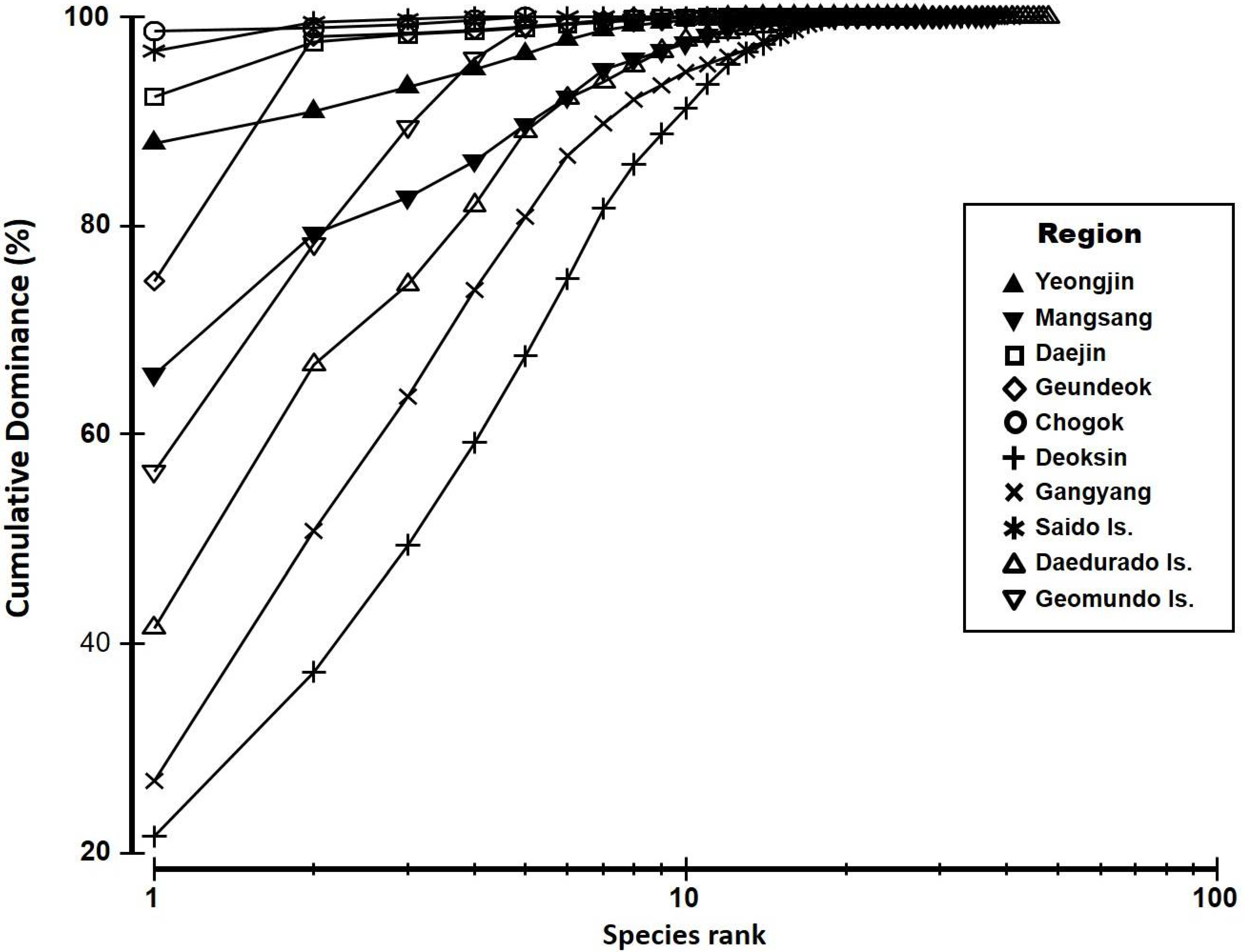
Samples were compared in terms of the diversity index [28], richness index [29], evenness index [30], and dominance index [31] to understand the community characteristics. The dominance depending on the biomass of the emerged macroalgae was compared with K-dominance curves. The community index was calculated, and K-dominance curves were plotted using Plymouth Routines Multivariate Ecological Research (PRIMER) version 6.0 and PERMANOVA+ [32].
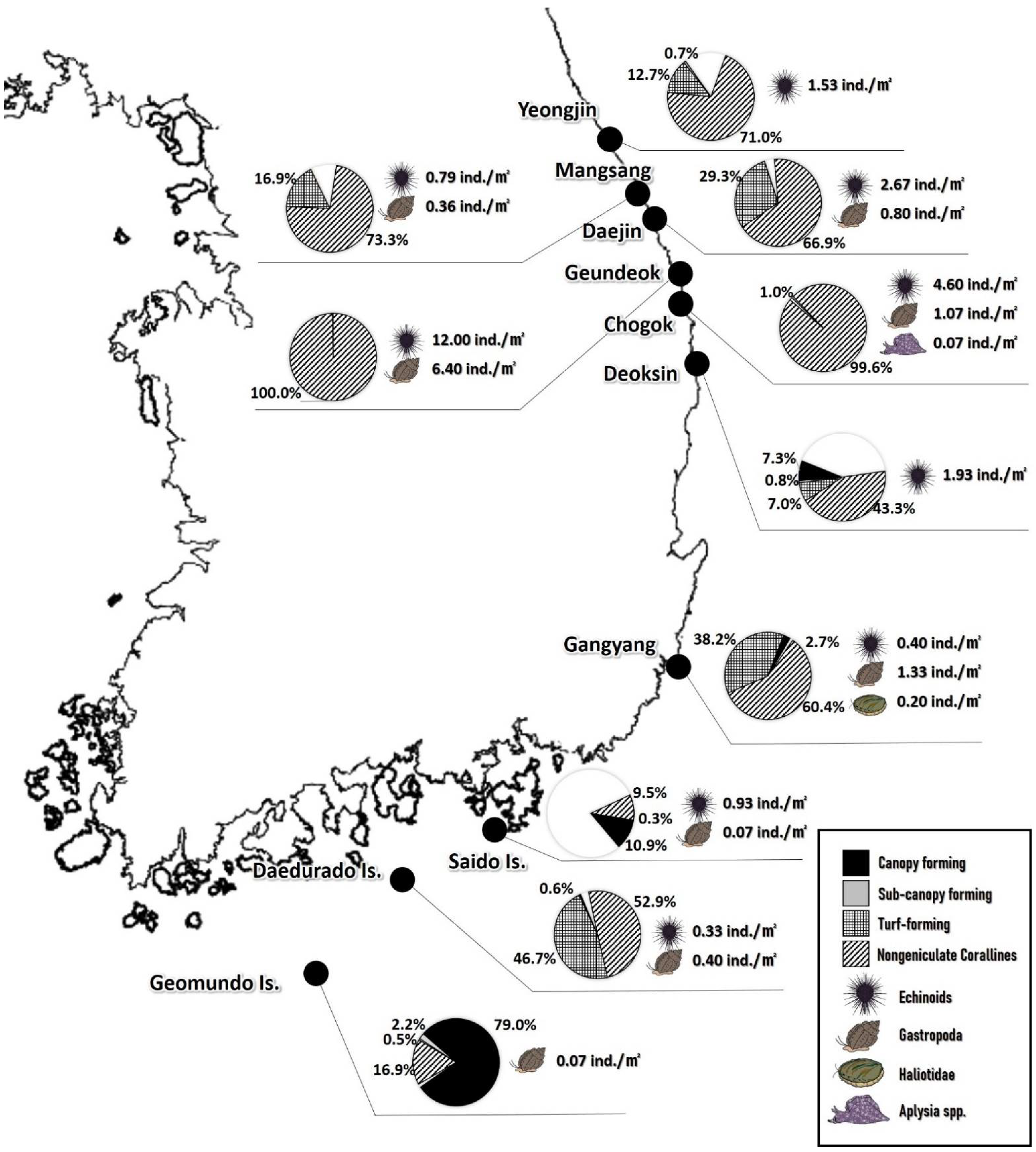
Comparisons of macroalgae coverage between sea areas showed that most of the species that appeared were composed of large brown algae, melobesidean algae, and macroalgae < 10 cm in body, which were length classified into four groups: canopy forming, sub-canopy forming, turf forming, and crustose corallines, based on their compositional characteristics [33,34,35]. Large brown algae were classified into either canopy-forming or sub-canopy-forming groups according to the body length of each species; all macroalgae except melobesidean algae were classified into the turf-forming group and the coverage of each group was schematically compared (Figure 1).
The EEI-c model from Orfanidis et al. [36] was applied to evaluate the community status. After the samples were classified into ESGⅠ (subgroups: ⅠA, ⅠB, ⅠC) and ESG Ⅱ (subgroups: ⅡA, ⅡB) (comprising five groups) by species according to the macroalgae classification and ecological characteristics, the EEI-c was calculated using the sum of the coverage for each group.
3. Results
3.1. Marine Environment and Substrate Characteristics
The substrate characteristics for each sea area showed that although rock masses were predominant in most of the sea areas, such as Yeongjin, Mangsang, Daejin, Geundeok, Chogok, and Geomundo Is., sandy substrates were somewhat mixed, and Gangyang was composed of large and small rock masses and gravels. However, the surroundings of Deoksin, Saido Is., and Daedurado Is. were composed of sand or mudflats except for bedrock near the shorelines.
The survey of marine environmental data showed that transparency was ≥10 m in the central and northern parts of the eastern coasts. This figure lowered in the order of the southern part of the eastern coasts and the southern coasts. The sea surface temperature was 15.13–18.67 °C, salinity was 31.81–33.75 psu, and pH was 8.08–8.35, showing no significant differences. DO was generally high on the eastern coasts, showing 7.57–8.22 mg/L on the eastern coasts and 6.61–7.88 mg/L on the southern coasts. DIN, DIP, and chlorophyll a (Chl-a) values varied depending on the sea area. The water quality index (WQI) calculated by combining the water quality items was determined as Grade 1 in Youngjin, Mangsang, and Daejin, Grade 2 in Deoksin, Gangyang, Saido Is., and Geomundo Is., and Grade 3 in Geundeok, Chogok, and Daedurado Is. (Table 2).
3.2. Species Composition
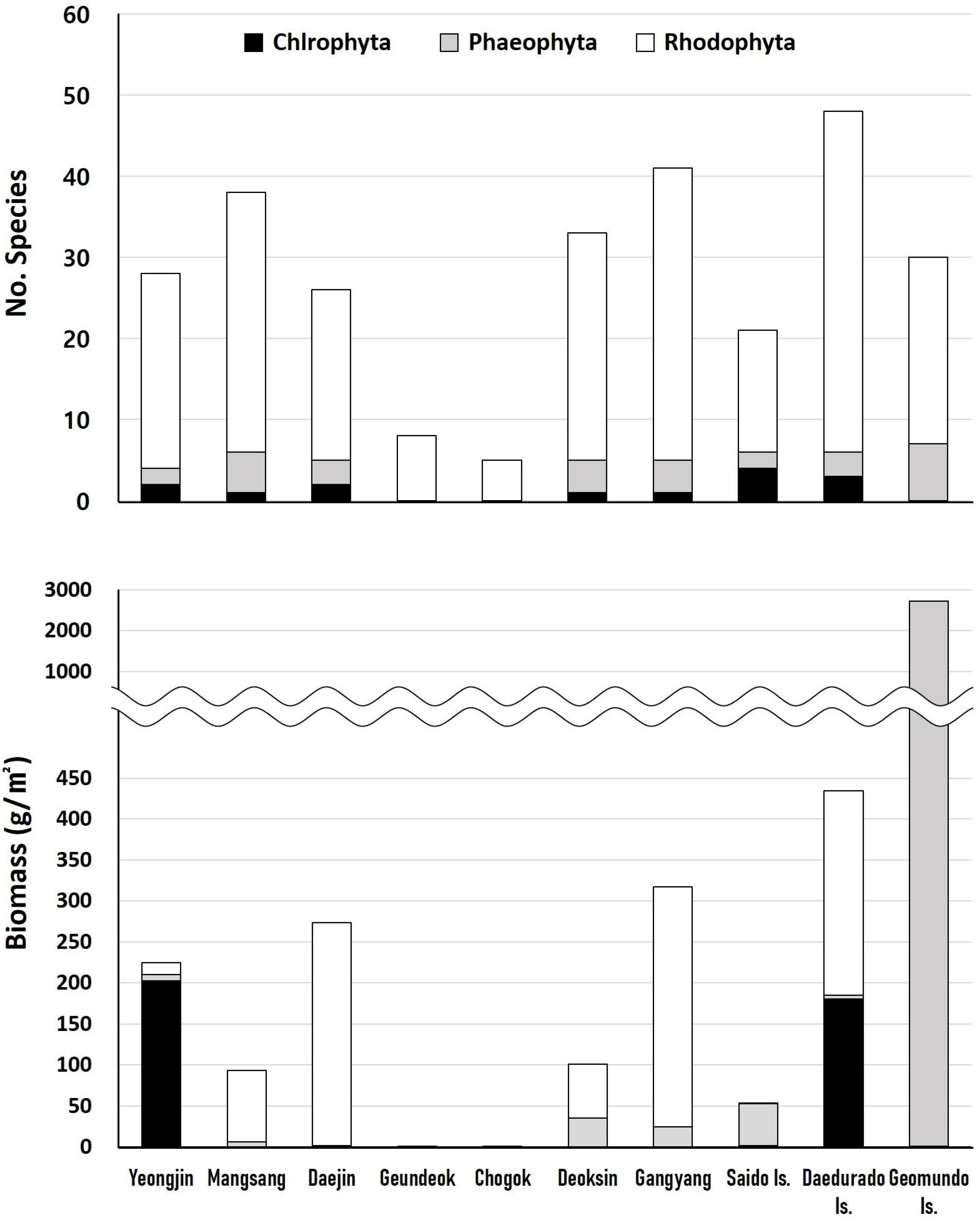
In total, 111 species of macroalgae appeared during the study period: 8 green algae, 16 brown algae, and 87 red algae, including 85 species in the eastern coasts and 72 species in the southern coasts. The proportion of red algae was very high, at 81–100% in the eastern coasts and 77–88% in the southern coasts. By sea area, 48 species appeared around Daedurado Is., which was the most, and 5 species appeared around Samcheok Chogok, which was the fewest (Figure 2). Melobesidean algae appeared in all sea areas. The frequency of Symphyocladia marchantioides and Plocamium telfairiae was high throughout the sea areas, and about half of all species appeared as fine-sized filamentous algae, such as Bryopsidaceae, Derbesiaceae, Cladophoraceae, Erythrotrichiaceae, Bonnemaisoniaceae, Ceramiaceae, Dasyaceae, Stylonemataceae, Colaconemataceae, and Callithamniaceae.
3.3. Biomass, Dominant Species, and Ecological Index
The average biomass in the seven sea areas on the eastern coasts was 114.42 g/m2, including 29.18 g/m2 for green algae, 10.64 g/m2 for brown algae, and 104.60 g/m2 for red algae. That in the three sea areas on the southern coasts were 1056.84 g/m2, including 60.63 g/m2 for green algae, 907.60 g/m2 for brown algae, and 88.60 g/m2 for red algae. The results showed that the biomass of red algae was high in the eastern coasts and that of brown algae was high in the southern coasts. By sea area, the biomass around Geomundo Is. was remarkably high, at 2683.02 g/m2, and that at Geundeok (0.33 g/m2) and Chogok (0.29 g/m2) were very low, due to the severity of the barren ground phenomenon (Figure 2).
The dominant species in each sea area were 197.73 g/m2 (87.9%) of Codium fragile in Yeongjin, 61.41 g/m2 (65.7%) of P. telfairiae in Mangsang, 252.62 g/m2 (92.3%) of P. telfairiae in Daejin, 0.24 g/m2 (74.7%) of Acrosorium ciliolatum in Geundeok, 0.28 g/m2 (98.6%) of A. ciliolatum in Chogok, 21.85 g/m2 (21.6%) of Sargassum horneri in Deoksin, 85.00 g/m2 (26.8) of Gelidium elegans in Gangyang, 51.29 g/m2 (96.7%) of S. horneri in Saido Is., 180.43 g/m2 (41.5%) of Ulva australis in Daedurado Is., and 1513.83 g/m2 (56.4%) of S. macrocarpum in Geomundo Is. Examining the top dominant species by sea area, r-selection species (annual opportunistic species) dominated the remaining seas except that around Geomundo Is.
The ecological index was excluded because the biomass in Geundeok and Chogok were significantly low. The species richness, evenness, diversity, and dominance indices were in the ranges of 3.67–8.16, 0.05–0.69, 0.16–2.40, and 0.12–0.94, respectively. The species richness index was highest in Mangsang, the evenness and diversity indices were highest in Deoksin, and the dominance index was highest in Saido Is., respectively (Table 3).
The K-dominance curves for the eight stations—excluding Geundeok and Chogok, whose biomass was significantly low—varied slightly, depending on sea areas (Figure 3). The three sea areas, Saido Is., Daejin, and Yeongjin, had 5–6 dominant species, and the dominance index of the top dominant species was significantly high as “severe”. The dominance index of the top dominant species was ≥60% in Mangsang. However, the dominance index of subdominant species was not as large as “major”. The dominance index of the top five dominant species in Geomundo Is. was almost 100%, showing the “major” state with a relatively steep slope in the curve. The species in the three sea areas—-Daedurado Is., Gangyang, and Deoksin—were relatively evenly distributed in a “moderate” state.
3.4. Current Status of Barren Ground and Community States
Macroalgae coverage by each sea area was 84.50% in Yeongjin, 90.35% in Mangsang, 96.31% in Daejin, 100.14% in Geundeok, 100.65% in Chogok, 58.31% in Deoksin, 101.22% in Gangyang, 20.70% in Saido Is., 103.26% in Daedurado Is., and 98.69% in Geomundo Is., the coverage of which was mostly close to 100%. The coverage of Deoksin and Saido Is. was relatively low compared to other sea areas; in particular, the coverage of Saido Is. was significantly low at 20%.
According to the hierarchical structure of the species appearing in each sea area, the species were classified into four groups: canopy-forming, sub-canopy-forming, turf-forming, and non-geniculate corallines. The results showed the coverage of melobesidean algae in Geundeok and Chogok reached 100%. The coverage of melobesidean algae in the five sea areas of Youngjin, Mangsang, Daejin, Deoksin, and Gangyang and Daedurado Is. was high, at 43.27–73.30%, and turf-forming and canopy-forming species were mixed. The coverage of melobesidean algae around Geomundo Is., unlike other sea areas, was low, and the coverage of canopy-forming and sub-canopy-forming species was significantly higher.
Analysis of the ecological status class (ESC), which represents the ecological state of the community, was conducted using the ecological evaluation index continuous formula (EEI-c). The results showed that as a cover of macroalgae species, when melobesidean algae were included, the ESC was high in Mangsang, Daejin, Geundeok, Chogok, Geomundo Is., good–high in Yeongjin, Deoksin, Gangyang, and Saido Is., and good–moderate in Daedurado Is.. When melobesidean algae were excluded, the ESC was high in Geomundo Is., good–high in Daejin, good–moderate in Mangsang, Deoksin, and Saido Is., moderate–low in Yeongjin and Gangyang, and bad in Daedurado Is. As a result, the ESC was evaluated as 1–2 grades lower in all sea areas, except Geomundo Is. (Table 4).
3.5. Density of Grazers
The average density of grazers was 0.07 ind/m2 in Geomundo Is., which was the lowest, 0.73 ind/m2 in Daedurado Is., 1.00–1.93 ind/m2 in Youngjin, Mangsang, Deoksin, Gangyang, and Saido Is., and 3.47 ind/m2 and 5.73 ind/m2 in Daejin and Chogok, respectively, and 18.40 ind/m2 in Geundeok, which was the highest. The density of sea urchins was high in most sea areas and that of snails was high in Gangyang, Daedurado Is., and Geomundo Is. Nearly no abalone or aplysia were observed (Figure 1).
4. Discussion
According to the results of species composition and average biomass by sea area analyses, the eastern coasts had 5–48 species and an average biomass of 114.42 g/m2 (0.29–273.60 g/m2), while the southern coasts had 21–48 species and an average biomass of 1056.84 g/m2 (53.03–2683.02 g/m2), showing a significant difference by sea area. Regarding the ratio of occurrence by taxon, red algae showed the highest ratio, which was common across the surveyed sea areas. The biomass ratio of red algae was high on the eastern coasts, while that of brown algae was high in the southern coasts. To identify the change in seaweed flora, a comparison was made with previous studies on seaweed communities. In the case of the eastern coasts, previous studies [13,14,15,16,17,18,19,20,37] reported that the number of species was 12–152 and the average biomass was 1355.67 g/m2 (155.89–7349.56 g/m2), which was significantly lower in this study. In the case of the southern coasts, previous studies [25,38,39,40,41] reported the number of species was 21–118, and the average biomass was 1140.62 g/m2 (548.96–2230.00 g/m2). By comparison, this study showed that the number of species has decreased, while biomass remains similar. These results are similar to those from the recent FIRA [8,10] analysis on the occurrence of barren ground areas, which confirmed that the decrease in seaweed communities due to barren ground is more severe on the eastern than southern coasts.
Regarding the community structure analyzed by species biomass, biomass was low, and opportunistic species were dominant in all sea areas, except Geomundo Is., whose cluster state suggested disturbance was severe in each sea area [33,42,43,44]. These results suggest that the community state can vary rapidly, depending on surrounding environment or seasonal changes. Furthermore, because the biomass in the community constituent species, excluding the dominant species, is poor in most sea areas, a rapid change in the dominant species or the severe extinction of macroalgae could occur. The biomass of these major dominant species is also shown in the results of the ecological index and K-dominance curve analysis. Since Saido Is., Daejin, and Yeongjin seas are in a “severe” state, changes in communities would be severe because of disturbances. Although Mangsang, Daedurado Is., Gangyang, and Deoksin are in “major” or “moderate” states, considering that the community is composed mainly of opportunistic species, changes to communities would be significant, depending on seasonal transitions. K-dominance was evaluated to appropriately reflect the current state. This is because k-dominance was determined as a “major” state, despite the dominance of perennial large brown algae, such as Sargassum spp. and Ecklonia cava surrounding Geomundo Is. This is in consideration of the change in the biomass of large brown algae, depending on the season effects on the entire community [45,46].
Comparisons of the coverage of the species that appeared showed all macroalgae were low in Deoksin and Saido Is. due to the predominantly sandy substrate characteristics. Turf-forming macroalgae were mixed in the central and north-eastern coasts, with melobesidean algae covering the entire bedrock. The coverage of melobesidean algae was relatively low in Gangyang, in the southern part of the eastern coasts, and around Daedurado Is. in the southern coasts, and the coverage of turf-forming algae tended to increase. The coverage of melobesidean algae was high at stations located in the eastern coasts, and that of other macroalgae was low, indicating the severe state of barren ground [3,47]. Moreover, the state of barren ground was determined to be severe in Geundeok and Chogok, where few macroalgae other than melobesidean algae were growing. The coverage of the K-selection canopy-forming and sub-canopy-forming algae was high in Geomundo Is., while that of opportunistic turf-forming algae was low. The biomass also showed similar characteristics to the coverage and ecological indices; for example, the species richness and diversity indices were also low. Geomundo Is. may provide favorable conditions for large brown algae to grow because it is less affected by external pressures, such as the influx of pollutants from land or artificial disturbances induced by human activities, due to its geographical characteristics of being far from the coasts [33]. Large brown algae, such as S. macrocarpum, S. fulvellum, and S. horneri, form a canopy to block light, thereby inhibiting the growth of turf-forming algae that constitute the understory [48,49].
Analysis results of the ecological state of the community, using the EEI-c model from Orfanidis et al. [36], showed that because melobesidean algae are classified as ESG IC due to their transitional characteristics, a difference of 1–2 grades occurred for each sea area, depending on the presence of melobesidean algae. When conducting an analysis excluding melobesidean algae, Mangsang, Daejin, Deoksin, and Saido Is., which had poor seaweed community states, showed “good–moderate” or “good–high” states. Gangyang, where the seaweed flora were the richest on the eastern coasts, showed a “moderate–low” state, indicating different results from the average biomass and total coverage of the community. The EEI-c indicated the community state only by the relative ratio of ESG I and ESG II, without considering the total coverage rate of macroalgae, and showed the community state of “good–moderate”, with a default value of 5.74. Thus, obtaining clear results on the community state when applying the EEI-c is difficult for a sea area with a low coverage of total macroalgae.
Barren ground refers to a phenomenon in which sheet-form macroalgae become extinct due to various causes, such as an increase in water temperature, artificial disturbances, and environmental pollution. Melobesidean algae cover rock mass, and excessive grazing pressure by grazers is also known to have certain effects [50,51]. In addition, melobesidean algae, a major causative organism, have an antifouling characteristic that limits the attachment of other macroalgae spores onto the surface through sloughing [52,53,54]; this mechanism acts species specifically [55]. Thus, although the correlations between marine environment by sea area, the average biomass of macroalgae, the coverage of melobesidean algae, and the density of grazers were analyzed, the correlations among marine environment factors are not clear, except for the fact that Geomundo Is., which had the highest biomass, had a significantly higher DIN value of 140.1 µg/L compared to other sea areas and that its DIP value was low at 12.0 µg/L. Although the coverage of melobesidean algae showed positive correlations (r2 = 0.641, p < 0.05) with sea surface temperature and the density of grazers, these correlations were not significant (Table 5). Future long-term monitoring studies are necessary to closely understand the relationship between the condition of seaweed beds and the actual status of barren ground based on physical and ecological factors.
Based on a comprehensive review of the above results, since sea areas, such as Deoksin and Saido Is., are unsuitable for macroalgae growth due to their predominantly sandy characteristics, it is necessary to establish a growth base for the recruitment and growth of macroalgae. Gangyang and Daedurado Is. have a low coverage of melobesidean algae, which are mostly composed of turf-forming macroalgae. Although these turf-forming algae contribute greatly to the production and circulation of [56], these algae, as an opportunistic species, are subjected to disturbances by seasonal or environmental changes. Therefore, they lower the stability of the entire seaweed community. Thus, it is necessary to perform seasonal monitoring to more closely analyze the characteristics of sea areas. Furthermore, the transplantation of large perennial brown algae, such as Sargassum spp. and Ecklonia spp., can be considered to increase the stability of communities. Despite Geomundo Is.’s stable community, formed by dense seaweed forests made of large brown algae, Geomundo Is. has low species variety, as its canopy-forming macroalgae inhibit the growth of other microalgae. If this phenomenon continues, melobesidean algae populations could expand, as these species adapt well to where the light is inhibited] [57,58]. Thus, continuous monitoring and action plans to manage seaweed forests are required. The five sea areas—Yeongjin, Mangsang, Daejin, Chogok, and Geundeok—exhibited the characteristic of distributing only some turf-forming algae in the state where melobesidean algae covered the entire rock mass. In this sea area, it is necessary to consider an improvement in substrates, such as rocky cleaning, out of consideration for the antifouling mechanism of widely distributed melobesidean algae [59]. In particular, in the case of Geundeok and Chogok, it is necessary to reduce the density of sea urchins to increase the recruitment and early growth of macroalgae [60,61]. In relation to the species-specific antifouling mechanism characteristic of melobesidean algae on the eastern coasts, practical ecological restoration is necessary to overcome barren ground by conducting ecological community studies on species composition and the species-inhibiting actions of melobesidean algae.
5. Conclusions
This study conducted a community investigation to find out the characteristics of seaweed communities and the current status of barren ground in natural seaweed beds. The characteristics of seaweed communities differed by sea area, and red algae, an annual opportunistic species, dominated the eastern coasts, and perennial large brown algae dominated the southern coasts. As a result of the evaluation on community status using the EEI-c model, the status of the community differed, depending on the inclusion of melobesidean algae, and the model application was not suitable in habitats with low seaweed coverage. As a result of the study, the seaweed community showed a tendency to gradually decrease according to the barren ground phenomenon, and there was a significant difference in the characteristics of seaweed community in the seaweed beds and barren ground area. The seaweed bed areas were less affected by external pressures or disturbances, so it was a place where large brown algae could grow in a good marine environment. Barren ground refers to a phenomenon in which macroalgae become extinct due to various causes, such as an increase in water temperature, artificial disturbance, and environmental pollution, and excessive grazing pressure by grazers is also known to have certain effects. Therefore, it seems necessary to introduce a marine ecological restoration and management system, incorporating the concept of ecological engineering in the sea area where barren ground is serious.
Author Contributions
Data curation, S.W.J.; Resources, H.S.R.; Writing—original draft, C.G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Korea Institute of Ocean Science & Technology (PEA0013 and PEA0016).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Choi, C.G.; Kim, H.G.; Sohn, C.H. Transplantation of young fronds of Sargassum horneri for construction of seaweed beds. J. Korean Fish. Soc. 2003, 36, 469–473. [Google Scholar] [CrossRef]
- Kang, S.K. An exploratory study on the ecosystem service and benefit indicators of natural seaweed beds. J. Fish. Bus Adm. 2016, 47, 53–69. [Google Scholar] [CrossRef] [Green Version]
- Park, G.J.; Ju, H.; Choi, O.I.; Choi, C.G. Community structure and distribution of natural seaweed beds on the Eastern Coast of Korea. J. Korean Soc. Mar. Environ. Saf. 2017, 23, 338–346. [Google Scholar] [CrossRef]
- Smale, D.A.; Burrows, M.T.; Moore, P.; O’Connor, N.; Hawkins, S.J. Threats and knowledge gaps for ecosystem services provided by kelp forests: A northeast Atlantic perspective. Ecol. Evol. 2013, 3, 4016–4038. [Google Scholar] [CrossRef] [Green Version]
- Teagle, H.; Hawkins, S.J.; Moore, P.J.; Smale, D.A. The role of kelp species as biogenic habitat formers in coastal marine ecosystems. J. Exp. Mar. Biol. Ecol. 2017, 492, 81–98. [Google Scholar] [CrossRef]
- Kang, J.W. Illustrated Encyclopedia of Fauna & Flora of Korea, Vol. 8. Marine Algae; Ministry of Education: Seoul, Korean, 1968; 465p. [Google Scholar]
- KORDI. A Preliminary Marine Ecological Study for Gojeong-Ri Power Plant Site; BSPI 00014-14-3; Korea Ocean Research & Development Institute: Ansan, Korea, 1978; 138p. [Google Scholar]
- FIRA. Barren Grounds Spreads over 60% in Bedrock on the East Sea. Press Release (15 July 2015). Available online: https://www.fira.or.kr/fira/fira_010201_2.jsp?mode=view&article_no=9800&board_wrapper=%2Ffira_010201_2.jsp&pager.offset=870&board_no=7 (accessed on 5 May 2022).
- Choi, Y.W.; Hwang, S.D.; Kim, D.K. Monitoring of algae composition and barren ground in Yeosu, South Korea. J. Fish. Res. Manage 2012, 2, 13–28. [Google Scholar]
- FIRA. The Beautiful South Sea! However…the Barren Grounds Progresses. Press Release (2 September 2016). Available online: https://www.fira.or.kr/fira/fira_010201_2.jsp?mode=view&article_no=13372&board_wrapper=%2Ffira_010201_2.jsp&pager.offset=780&board_no=7 (accessed on 5 May 2022).
- Lee, K.W. Observation of algal community near Dongbaeksum, Haeundae. Bull. Fish. Jeju. Univ. 1974, 5, 319–331. [Google Scholar]
- Lee, K.W.; Kang, J.W. A preliminary survey of the algal flora and communities of Dongbaiksum, Pusan. Publ. Mar. Lab. Pusan. Fish. Coll. 1971, 4, 29–37. [Google Scholar]
- Choi, C.G.; Chowdhury, M.T.H.; Choi, I.Y.; Hong, Y.K. Marine algal flora and community structure in Kijang on the Southern East coast of Korea. Sea 2010, 15, 133–139. [Google Scholar]
- Choi, C.G.; Rho, H.S. Marine algal community of Ulsan, on the Eastern coast of Korea. Korean J. Fish. Aquat. Sci. 2010, 43, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Han, S.H.; Hwang, Y.H.; Son, M.H.; Choi, H.G.; Jang, J.G. Seasonal variation in seaweed community structure in the subtidal zone of the Southern part of the East coast of Korea. Korean J. Fish. Aquat. Sci. 2018, 51, 571–578. [Google Scholar] [CrossRef]
- Jeong, H.D.; Hong, S.E.; Kim, S.W.; Han, M.S.; Jang, S.H. Community structure and biological indicator species of marine benthic algal at intertidal zone in the three areas of the East coast of Korea. J. Korean Soc. Mar. Environ. Saf. 2014, 20, 609–618. [Google Scholar] [CrossRef]
- Kim, Y.D.; Park, M.S.; Yoo, H.I.; Kim, S.W.; Jeong, H.D.; Min, B.H.; Jin, H.J. Characteristics of seasonal variations of subtidal seaweed community structure at three areas in the East coast of Korea. J. Environ. Sci. Int. 2012, 21, 1407–1418. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.D.; Park, M.S.; Yoo, H.I.; Min, B.H.; Jin, H.J. Seasonal variations of seaweed community structure at the subtidal zone of Bihwa on the East coast of Korea. Korean J. Fish. Aquat. Sci. 2012, 45, 262–270. [Google Scholar] [CrossRef]
- Kwon, C.J.; Choi, C.G. Marine algal flora and community structure in subtidal zone of Wangdol-cho on the East Coast of Korea. Sea 2014, 19, 191–201. [Google Scholar] [CrossRef]
- Shin, B.G.; Kwon, C.J.; Lee, S.M.; Choi, C.G. Marine algal flora and community structure of Igidea area in Busan, Korea. J. Korean Soc. Mar. Environ. Saf. 2014, 20, 121–129. [Google Scholar] [CrossRef]
- Song, S.H.; Choi, J.S.; Sohn, C.H. Summer algal flora at Odongdo, Yeosu. Thesis Collect. Yeosu Fish. Tech. Coll. 1970, 4, 18–28. [Google Scholar]
- Heo, J.S.; Yoo, H.I.; Park, E.J.; Ha, D.S. Macroalgal community structure on the subtidal of Southern six Islands, Korea. Korean J. Environ. Biol. 2017, 35, 595–603. [Google Scholar] [CrossRef]
- Oh, J.C.; Aha, J.K.; Kim, C.D.; Jeong, J.B.; Choi, H.G. Seasonal variations in the macroalgal flora and community structure in Hallyeohaesang national park on the South coast of Korea. Korean J. Fish. Aquat. Sci. 2015, 48, 768–775. [Google Scholar] [CrossRef]
- Oh, J.C.; Choi, H.G.; Kim, C.D.; Ahn, J.K. Ecological evaluation of marine national parks based on seaweed community index. Korean J. Fish. Aquat. Sci. 2016, 49, 385–392. [Google Scholar] [CrossRef]
- Park, M.S.; Yoo, H.I.; Heo, J.S.; Kim, Y.D.; Choi, H.G. Seasonal variation in the marine algal flora and community structure along the Tongyeong coast, Korea. Korean J. Fish. Aquat. Sci. 2011, 44, 732–739. [Google Scholar] [CrossRef]
- Song, J.N.; Park, S.K.; Heo, J.S.; Kim, B.Y.; Yoo, H.I.; Choi, H.G. Summer seaweed flora and community structure of uninhabited islands in Goheung, Korea. Korean J. Fish. Aquat. Sci. 2011, 44, 524–532. [Google Scholar] [CrossRef]
- Kim, H.S.; Boo, S.M.; Lee, I.K.; Sohn, C.H. National List of Species of Korea [Marine Algae]; National Institute of Biological Resources: Incheon, Korea, 2013; 336p. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; Univ-Illinois Press: Urbana, IL, USA, 1963; 125p. [Google Scholar]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Pielou, E.C. Mathematical Ecology, J; Wiley & Sons: New York, NY, USA, 1977. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N. Clarke KR Permanova+ for Primer: Guide to Software and Statistical Methods; PRIMER-E, Plymouth Marine Laboratory: Plymouth, UK, 2008; 214p. [Google Scholar]
- Arévalo, R.; Pinedo, S.; Ballesteros, E. Changes in the composition and structure of Mediterraneanrocky-shore communities following a gradient of nutrient enrichment: Descriptive study and test of proposed methods toassess water quality regarding macroalgae. Mar. Pollut. Bull. 2007, 55, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Irving, A.D.; Connell, S.D. Predicting understorey structure from the presence and composition of canopies: An assembly rule for marine algae. Oecologia 2006, 148, 491–502. [Google Scholar] [CrossRef]
- Marks, L.M.; Reed, D.C.; Holbrook, S.J. Life history traits of the invasive seaweed Sargassum horneri at Santa Catalina Island, California. Aquat. Invasions 2018, 13, 339–350. [Google Scholar] [CrossRef]
- Orfanidis, S.; Panayotidis, P.; Ugland, K. Ecological evaluation index continuous formula (EEI-c) application: A step forward for functional groups, the formula and reference condition values. Medit. Mar. Sci. 2011, 12, 199–231. [Google Scholar] [CrossRef] [Green Version]
- Park, G.J.; Choi, C.G. A study on the community structure of intertidal benthic marine algae in Youngil Bay, Eastern coast of Korea. Korean J. Fish. Aquat. Sci. 2009, 42, 664–673. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.G.; Kim, J.H.; Chung, I.K. Temporal Variation of Seaweed Biomass in Korean coasts: Yokjido, Gyeongnam Province. Algae 2008, 23, 311–316. [Google Scholar] [CrossRef]
- Kang, R.S.; Je, J.G.; Hong, J.S. Summer algal communities in the rocky shore of the South Sea of Korea I. Intertidal communities. Bull. Korean Fish. Soc. 1993, 26, 49–62. [Google Scholar]
- Kang, R.S.; Je, J.G.; Sohn, C.H. Summer algal communities in the rocky shore of the South Sea of Korea II. Subtidal communities. Bull. Korean Fish. Soc. 1993, 26, 182–197. [Google Scholar]
- Kang, P.J.; Nam, K.W. An ecological evaluation of marine algal vegetation in the coastal waters of Goseong, Southern Korea. Korean J. Fish. Aquat. Sci. 2015, 48, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; de Torres, M. A new methodology based on littoral community cartography dominated by macroalgae for the implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Rinne, H.; Korpinen, S.; Mattila, J.; Salovius-Lauréna, S. Functionality of potential macroalgal indicators in the northern Baltic Sea. Aquat. Bot. 2018, 149, 52–60. [Google Scholar] [CrossRef]
- Wells, E.; Wilkinson, M.; Wood, P.; Scanlan, C. The use of macroalgal species richness and composition on intertidal rocky seashores in the assessment of ecological quality under the European water framework directive. Mar. Pollut. Bull. 2007, 55, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Crawford, C.; Macleod, C.; Mitchell, I. Evaluation of Techniques for Environmental Monitoring of Salmon Farms in Tasmania; TAFI Technical Report (8); Tasmanian Aquaculture & Fisheries Institute: Hobart, Australia, 2002; 134p. [Google Scholar]
- Matthews, T.J.; Whittaker, R.J. On the species abundance distribution in applied ecology and biodiversity management. J. Appl. Ecol. 2015, 52, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.G.; Kwak, S.N.; Sohn, C.H. Community structure of subtitdal marine algae at Uljin on the East Coast of Korea. Algae 2006, 21, 463–470. [Google Scholar] [CrossRef]
- Goodsell, P.J.; Connell, S.D. Disturbance initiates diversity in recruitment of canopy-forming algae: Interactive effects of canopy-thinning and substratum availability. Phycologia 2005, 44, 632–639. [Google Scholar] [CrossRef]
- Wernberg, T.; Thomsen, M.S. The effect of wave exposure on the morphology of Ecklonia radiata. Aquat. Bot. 2005, 83, 61–70. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Env. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef] [Green Version]
- Vergés, A.; Doropoulos, C.; Malcolm, H.A.; Skye, M.; Garcia-Pizá, M.; Marzinelli, E.M.; Campbell, A.H.; Ballesteros, E.; Hoey, A.S.; Vila-Concejo, A.; et al. Long-term empirical evidence of ocean warming leading to tropicalization of fish communities, increased herbivory, and loss of kelp. Proc. Natl. Acad. Sci. USA 2016, 113, 13791–13796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keats, D.W.; Knight, M.A.; Pueschel, C.M. Antifouling effects of epithallial shedding in three crustose coralline algae (Rhodophyta, Coralinales) on a coral reef. J. Exp. Mar. Biol. Ecol. 1997, 213, 281–293. [Google Scholar] [CrossRef]
- Masaki, T.; Fujita, D.; Hagen, N.T. The surface ultrastructure and epithallium shedding of crustose coralline algae in an ‘Isoyake’ area of southwestern Hokkaido, Japan. Hydrobiologia 1984, 116–117, 218–223. [Google Scholar] [CrossRef]
- Pueschel, C.M.; Keats, D.W. Fine structure of deep-layer sloughing and epithallial regeneration in Lithophyllum neoatalayense (Corallinales, Rhodophyta). Phycol. Res. 1997, 45, 1–8. [Google Scholar] [CrossRef]
- Villas Bôas, A.B.; Figueiredo, M.A.d.O. Are anti-fouling effects in coralline algae species specific? Braz. J. Oceanogr. 2004, 52, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Copertino, M.; Connell, S.D.; Cheshire, A. The prevalence and production of turf-forming algae on a temperate subtidal coast. Phycologia 2005, 44, 241–248. [Google Scholar] [CrossRef]
- Payri, C.E.; Maritorena, S.; Bizeau, C.; Rodière, M. Photoacclimation in the tropical coralline alga Hydrolithon onkodes (Rhodophyta, Corallinaceae) from a French Polynesian reef. J. Phycol. 2001, 37, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Steneck, R.S. The ecology of coralline algal crust: Convergent patterns and adaptative strategies. Annu. Rev. Ecol. Syst. 1986, 17, 273–303. [Google Scholar] [CrossRef]
- Kwak, C.W.; Chung, E.Y.; Gim, T.Y.; Lee, J.H.; Kim, Y.S. Marine algal assemblages on artifical reefs in Jeju-do before and after rocky cleaning and the growth pattern of Ecklonia cava with water depth. J. Korean Soc. Fish. Mar. Edu. 2014, 26, 34–48. [Google Scholar] [CrossRef] [Green Version]
- Fagerli, C.W.; Norderhaug, K.M.; Christie, H.C. Lack of sea urchin settlement may explain kelp forest recovery in overgrazed areas in Norway. Mar. Ecol. Prog. Ser. 2013, 488, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Jeon, B.H.; Yang, K.M.; Kim, J.H. Changes in macroalgal assemblage with sea urchin density on the east coast of South Korea. Algae 2015, 30, 139–146. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Coverage of seaweed group and grazer density at the study sites.

Figure 2.
K-dominance curve of seaweed beds at the study sites.

Figure 3.
Species composition and biomass of seaweed beds at the study sites.


{kind=link}
{kind=link}
{kind=link}
Table 1.
GPS coordinates at study sites.
| Region | GPS Coordinates | |
|---|---|---|
| Yeongjin | 37°52′19.72″ N | 128°50′54.44″ E |
| Mangsang | 37°35′38.21″ N | 129°06′09.41″ E |
| Daejin | 37°34′35.86″ N | 129°07′05.22″ E |
| Geundeok | 37°20′28.64″ N | 129°16′03.49″ E |
| Chogok | 37°18′50.16″ N | 129°17′45.70″E |
| Deoksin | 36°52′37.75″ N | 129°25′20.20″ E |
| Gangyang | 35°23′25.96″ N | 129°21′16.68″ E |
| Saido Is. | 34°40′03.76″ N | 128°16′03.67″ E |
| Daedurado Is. | 34°33′48.81″ N | 127°43′18.89″ E |
| Geomundo Is. | 34°03′26.71″ N | 127°16′56.20″ E |
Table 2.
Marine environmental data at the study sites.
| Region | Dominated Substrate | Transparency (m) | SST (℃) | Salinity (psu) | pH | DO (mg/L) | DIN (μg/L) | DIP (μg/L) | Chl-a (μg/L) | WQI |
|---|---|---|---|---|---|---|---|---|---|---|
| Yeongjin | rock | 11.9 | 15.13 | 33.75 | 8.27 | 8.04 | 72.1 | 19.1 | 0.77 | 1 |
| Mangsang | rock | 12.1 | 16.01 | 33.66 | 8.20 | 7.94 | 57.2 | 18.7 | 0.71 | 1 |
| Daejin | rock | 12.7 | 16.39 | 33.65 | 8.21 | 7.90 | 71.7 | 19.5 | 0.77 | 1 |
| Geundeok | rock | 16.5 | 16.23 | 33.69 | 8.20 | 8.22 | 59.8 | 16.4 | 0.63 | 3 |
| Chogok | rock | 12.3 | 16.86 | 33.63 | 8.21 | 7.78 | 55.5 | 17.8 | 1.10 | 3 |
| Deoksin | sand | 8.0 | 17.54 | 33.73 | 8.23 | 7.75 | 47.6 | 17.3 | 0.92 | 2 |
| Gangyang | gravel | 4.7 | 16.83 | 33.64 | 8.14 | 7.57 | 83.0 | 21.6 | 1.03 | 2 |
| Saido Is. | sand | 4.5 | 18.67 | 33.11 | 8.13 | 7.88 | 39.6 | 19.2 | 0.76 | 2 |
| Daedurado Is. | sand & mud | 2.8 | 16.57 | 33.06 | 8.08 | 6.61 | 75.4 | 21.5 | 0.68 | 3 |
| Geomundo Is. | bedrock | 3.5 | 17.54 | 32.81 | 8.35 | 6.68 | 140.1 | 12.0 | 0.64 | 2 |
Table 3.
Ecological indices of seaweed beds at the study sites.
| Region | Richness | Eveness | Diversity | Dominance |
|---|---|---|---|---|
| Yeongjin | 4.99 | 0.18 | 0.62 | 0.78 |
| Mangsang | 8.16 | 0.37 | 1.35 | 0.46 |
| Daejin | 4.46 | 0.11 | 0.37 | 0.86 |
| Deoksin | 6.93 | 0.69 | 2.40 | 0.12 |
| Gangyang | 6.95 | 0.57 | 2.12 | 0.17 |
| Saido Is. | 5.04 | 0.05 | 0.16 | 0.94 |
| Daedurado Is. | 7.74 | 0.46 | 1.78 | 0.25 |
| Geomundo Is. | 3.67 | 0.37 | 1.25 | 0.38 |
Table 4.
Evaluation of the status of seaweed beds using EEI-c model at the study sites.
| Contents | Yeongjin | Mangsang | Daejin | Geundeok | Chogok | Deoksin | Gangyang | Saido Is. | Daedurado Is. | Geomundo Is. |
|---|---|---|---|---|---|---|---|---|---|---|
| ESG IA | 0.45 | 0.00 | 0.00 | 0.00 | 0.00 | 0.64 | 0.18 | 0.00 | 0.45 | 1.45 |
| ESG IB | 1.36 | 13.42 | 25.89 | 0.07 | 0.00 | 9.08 | 9.17 | 10.91 | 3.69 | 79.77 |
| ESG IC | 71.08 | 73.58 | 66.94 | 100.01 | 99.64 | 44.76 | 60.64 | 9.49 | 53.64 | 18.66 |
| ESG IIA | 1.43 | 3.35 | 3.30 | 0.04 | 1.00 | 4.95 | 31.12 | 0.05 | 28.18 | 0.09 |
| ESG IIB | 10.22 | 0.21 | 0.18 | 0.02 | 0.00 | 0.09 | 0.12 | 0.28 | 17.99 | 0.25 |
| Including non-geniculate coralline | ||||||||||
| ESG I | 44.19 | 54.88 | 60.88 | 60.06 | 59.78 | 34.76 | 43.90 | 14.42 | 35.59 | 76.47 |
| ESG II | 11.36 | 2.89 | 2.82 | 0.05 | 0.81 | 4.05 | 25.01 | 0.32 | 40.53 | 0.33 |
| EEI-c | 8.41 | 9.91 | 10.00 | 10.00 | 10.00 | 8.38 | 7.27 | 7.05 | 5.62 | 10.00 |
| ESC | G-H | High | High | High | High | G-H | G-H | G-H | G-M | High |
| Excluding non-geniculate coralline | ||||||||||
| ESG I | 1.65 | 11.03 | 20.73 | Not applicable | 8.79 | 7.68 | 8.75 | 3.85 | 66.30 | |
| ESG II | 11.36 | 2.89 | 2.82 | 4.05 | 25.01 | 0.32 | 40.53 | 0.33 | ||
| EEI-c | 4.93 | 6.51 | 7.37 | 6.21 | 4.44 | 6.54 | 3.10 | 10.00 | ||
| ESC | M-L | G-M * | G-H * | G-M * | M-L * | G-M * | Bad | High | ||
* G-H, Good-High; G-M, Good-Moderate; M-L, Moderate-Low.
Table 5.
Results of Pearson’s correlation analysis for marine environment and seaweed beds.
| Transparency | SST | Salinity | pH | DO | DIN | DIP | Chl-a | Grazer Density | |
|---|---|---|---|---|---|---|---|---|---|
| Mean Biomass | −0.281 | −0.374 | 0.022 | 0.125 | −0.025 | −0.026 | −0.138 | −0.182 | −0.309 |
| Coverage of Non-geniculate corallines | 0.365 | 0.641 * | −0.223 | −0.015 | 0.118 | −0.252 | −0.334 | −0.142 | 0.641 * |
*, p < 0.05.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jung, S.W.; Rho, H.S.; Choi, C.G. Seaweed Beds and Community Structure in the East and South Coast of Korea. J. Mar. Sci. Eng. 2022, 10, 689. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050689
AMA Style
Jung SW, Rho HS, Choi CG. Seaweed Beds and Community Structure in the East and South Coast of Korea. Journal of Marine Science and Engineering. 2022; 10(5):689. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050689
Chicago/Turabian StyleJung, Seung Wook, Hyun Soo Rho, and Chang Geun Choi. 2022. "Seaweed Beds and Community Structure in the East and South Coast of Korea" Journal of Marine Science and Engineering 10, no. 5: 689. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050689
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.