
Bioactive Compounds for Quorum Sensing Signal-Response Systems in Marine Phycosphere


1
Key Lab of Coastal Biology and Biological Resource Conservation, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai 264003, China
2
College of Marine Sciences, University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2022, 10(5), 699; https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050699
Submission received: 6 May 2022
/
Accepted: 18 May 2022
/
Published: 20 May 2022
(This article belongs to the Special Issue Critical Reviews in Natural Products and Enzymes from Marine Organisms)
{kind=link}
{kind=link}
Abstract
:Quorum sensing in the phycosphere refers to a sensor system in which bacteria secrete bioactive compounds to coordinate group behavior relying on cell density. It is an important way for algae and bacteria to communicate with each other and achieve interactions. It has been determined that quorum sensing is widely presented in the marine phycosphere, which involves a variety of bioactive compounds. Focused on these compounds in marine phycosphere, this review summarizes the types and structures of the compounds, describes the methods in detection and functional evaluation, discusses the ecological functions regulated by the compounds, such as modulating microbial colonization, achieving algae–bacteria mutualism or competition and contributing to marine biogeochemical cycles. Meanwhile, the application prospects of the compounds are also proposed, including controlling harmful algal blooms and producing biofuel. Future research should focus on improving detection techniques, developing more model systems and investigating the effects of climate change on the quorum-sensing pathway to further understand the mechanism and application potential of quorum sensing compounds. This review aims to present an overview of current research carried out in order to provide the reader with perspective on bioactive compounds involved in quorum sensing.
1. Introduction
Algae are the pivotal primary producers in the marine food web due to their high photosynthetic efficiency and rapid biomass production. They convert approximately half of the primary production into dissolved organic matter for heterotrophic prokaryotes such as bacteria [1]. The term “phycosphere” refers to a unique microenvironment surrounding the algal cell. It is composed of algal extracellular substances and possesses distinctive bacterial community composition from an ocean environment [2]. Bacteria can gather in the phycosphere and show higher concentration than in seawater. This microscale region is regulated by the release of algal organic matter, which feeds a large consortium of inhabiting bacteria [3].
The algae–bacteria interactions that occur in the phycosphere contribute to many biogeochemical processes in the marine ecosystem, including carbon, nitrogen and sulfur cycles. These interspecies communications require enormous metabolic activities linked to a complex signaling network [4]. Quorum sensing (QS) is a common bacterial communication system correlating to produce, release, accumulate and detect the extracellular autoinduced signaling molecules. It allows bacteria to synchronize behaviors on a population-wide scale, and thus achieve biological functions such as establishing biofilms, expressing virulence and bioluminescence [5]. Since the discovery of QS signals during cyanobacterial blooms in 1998, many reports have proved the existence of QS in the phycosphere [6,7,8]. In parallel, parts of the mechanisms are also elucidated in several ecological models, such as the Roseobacter lineage and marine algae Emiliania huxleyi [4].
The diverse natural products for QS have been identified in the marine phycosphere. Their structures and working mechanisms have gradually been illustrated. Firstly, they regulate intraspecific signaling pathway in bacteria. Now, their functions in regulating behavior of bacterial hosts are proposed, such as modulating algal settlement [9]. Previously, their roles were highly underrated because of their low density in the ocean [10]. Now, it is clear that these substances effect bacteria inhabitation in the coral surface mucus layer; thus, bacteria are capable of protecting coral health [11]. Moreover, evidence shows that the products can influence the activity of hydrolytic enzymes on sinking particulate organic carbon, and thus impact the marine biological carbon pump [12]. In this context, exploring how the QS compounds impact interspecific interactions is of great significance.
The role of QS in marine microbial ecosystems has been extensively studied during the last decade [13]. Diverse QS substances have been isolated from phycosphere; nevertheless, their potential roles in modulating algae–bacteria interactions and impacting marine ecology remain restricted. Despite the increasing use of omics approaches in cooperation with conventional procedures, more sensitive analytical techniques are yet to be developed. Simultaneously, additional research is required to explore the substances involved in the QS pathway. In this review, we focus on the most recent progress in this field, with the goal of expanding current knowledge about natural products for QS in marine phycosphere.
2. Diverse Substances Involved in QS Pathway
QS exists in both Gram-positive and Gram-negative bacteria. Although the types of signal compounds are various, the QS processes are in the similar routine that refers to the production, release and group-wide detection of signaling molecules [14]. Two groups of these diffusible molecules were mainly emphasized in a previous study: N-acyl homoserine lactones (AHLs) in Gram-negative bacteria and oligopeptides in Gram-positive bacteria. Aside from these, autoinducer-2 (AI-2) was suggested to be a universal signal across species because it was found in both Gram-positive and Gram-negative bacteria [15]. Other classes of signals were also identified in recent work; a majority of them were not first known as semiochemicals, such as tropodithietic acid (TDA), which was known as an antibacterial agent before being described as a QS signal [16]. In this review, we will mainly discuss AHLs, quinolones and AI-2 for their key roles in the phycosphere. We will also discuss several other types of compounds, such as TDA and IAA.
2.1. AHLs Type Signaling Molecules
In the marine environment, AHLs are the most common signals. N-2-oxohexanoyl-L-homoserine lactone (3OC6-HSL) was the first QS signal detected, which was capable of controlling the luminescence of the marine bacterium Vibrio fischeri [17]. The genetic components in its system had been subsequently addressed: a synthase gene luxI that produces the signal molecule; a transcription-activator gene luxR; and a bidirectional lux promoter [18]. These molecules possess the same structures, which contain a core N-acylated homoserine-lactone ring and a 4–18 carbon acyl chain [19].
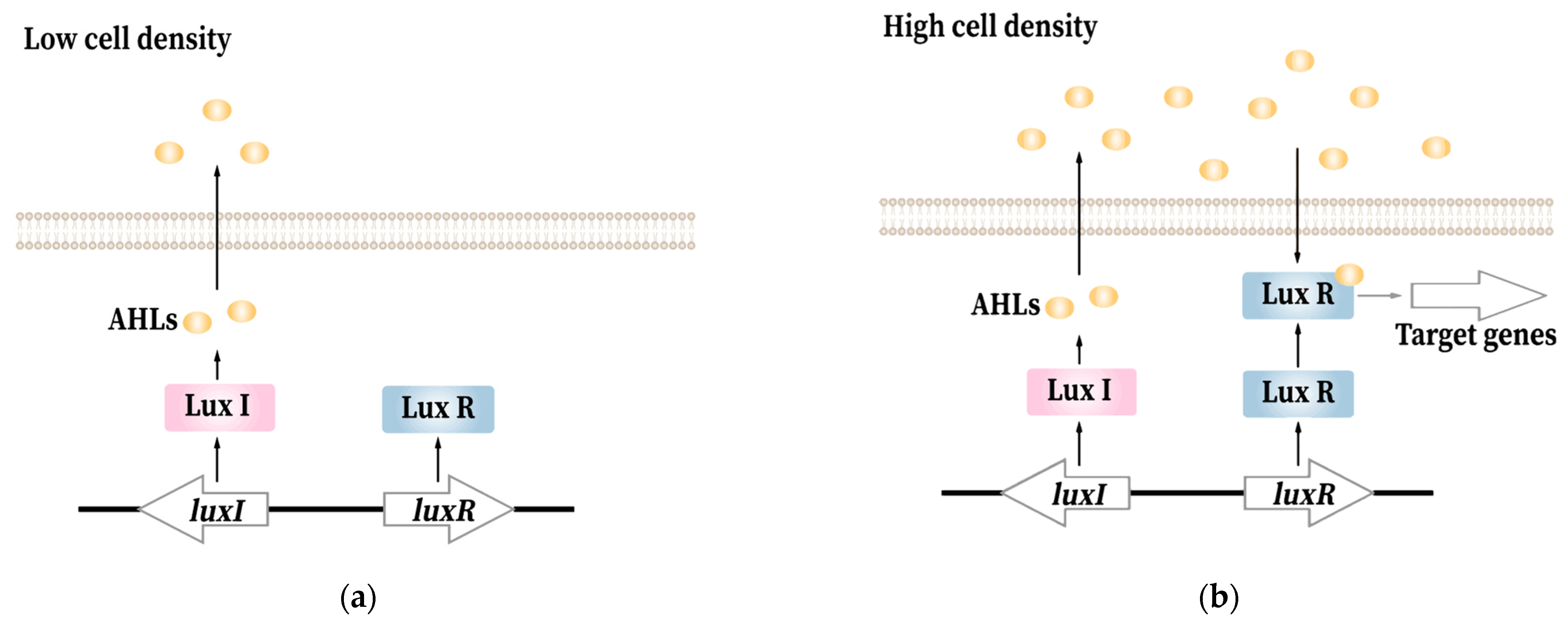
AHL-QS is widespread in Gram-negative bacteria. Its compounds include proteins homologous to the LuxI-AHL synthase and the LuxR transcriptional regulator proteins encoded by V.fischeri. Through the LuxI protein, AHLs are produced and freely diffuse out of the cell [13]. With the density of the cell population increasing, the concentration of AHLs accumulates in the near microenvironment. The receptor protein LuxR binds its cognate AHL. The luxR-AHL complex further triggers gene expression (Figure 1) [19].
The AHLs molecules detected in the phycosphere usually possess a long acyl chain. According to an analysis of more than 100 isolated marine bacterial strains, the acyl chains of AHLs ranged from C8 to C18. Some bacteria associated with diatom showed the capability to synthesize more than one type of AHL [7]. The production of AHLs includes a common pathway of deriving the lactone moiety from S-adenosylmethionine (SAM) and obtaining the acyl chain from intermediates of fatty-acid biosynthesis through the effect of LuxI enzymes [14]. There are exceptions. Microalgae Emiliania huxleyi secrete pCA when they age. Their associated roseobacters switch their metabolism to produce algaecides in response to pCA and use pCA as substrate for synthase p-coumaroyl-HSL (pCA-HSL) signals [20,21]. Another member of Roseobacter clade Nautella italica R11 contains a unique luxR-type gene, varR, which is verified as controlling bacterial colonization and virulence. It was the first example to demonstrate that luxR-type gene plays a role in bacterial-induced macroalgal disease [22]
2.2. Quinolones Type Signaling Molecules
Quinolones refer to a class of molecules which are structurally derived from the heterobicyclic. The differences between these compounds are various substitutions in carbocyclic and heteroaromatic rings [23]. Several members of quinolones have antibacterial activity, and thus become promising clinic drugs [24]. In the marine environment, several alkylquinolones (AQs) have been identified, and some of them are utilized by bacteria as QS signals [23].
The 2-heptyl-4-quinolones (HHQ) is an important member of those alkylquinolones, which is mainly synthesized by marine bacteria Pseudomonas and Alteromonas strains [23]. Multiple genes are involved in HHQ biosynthesis, such as pqsR, pqsH, pqsL and pqsABCDE. Among all of them, pqsR plays an essential role in encoding PsqR, which is the transcriptional regulator of pqsABCDE. Along with that, pqsABCDE controls the production of all AQs, which involves the synthesis of enzymes and inactive intermediates [25]. At the nanomolar concentrations, HHQ were capable of arresting algal cell growth without inducing cell mortality [26]. Recently, the finding that HHQ possesses algicidal effects on microalgae indicates it may modulate algal proliferation in the marine environment [27].
2.3. AI-2 Type Signaling Molecules
2.3.1. AI-2 in Marine Phycosphere
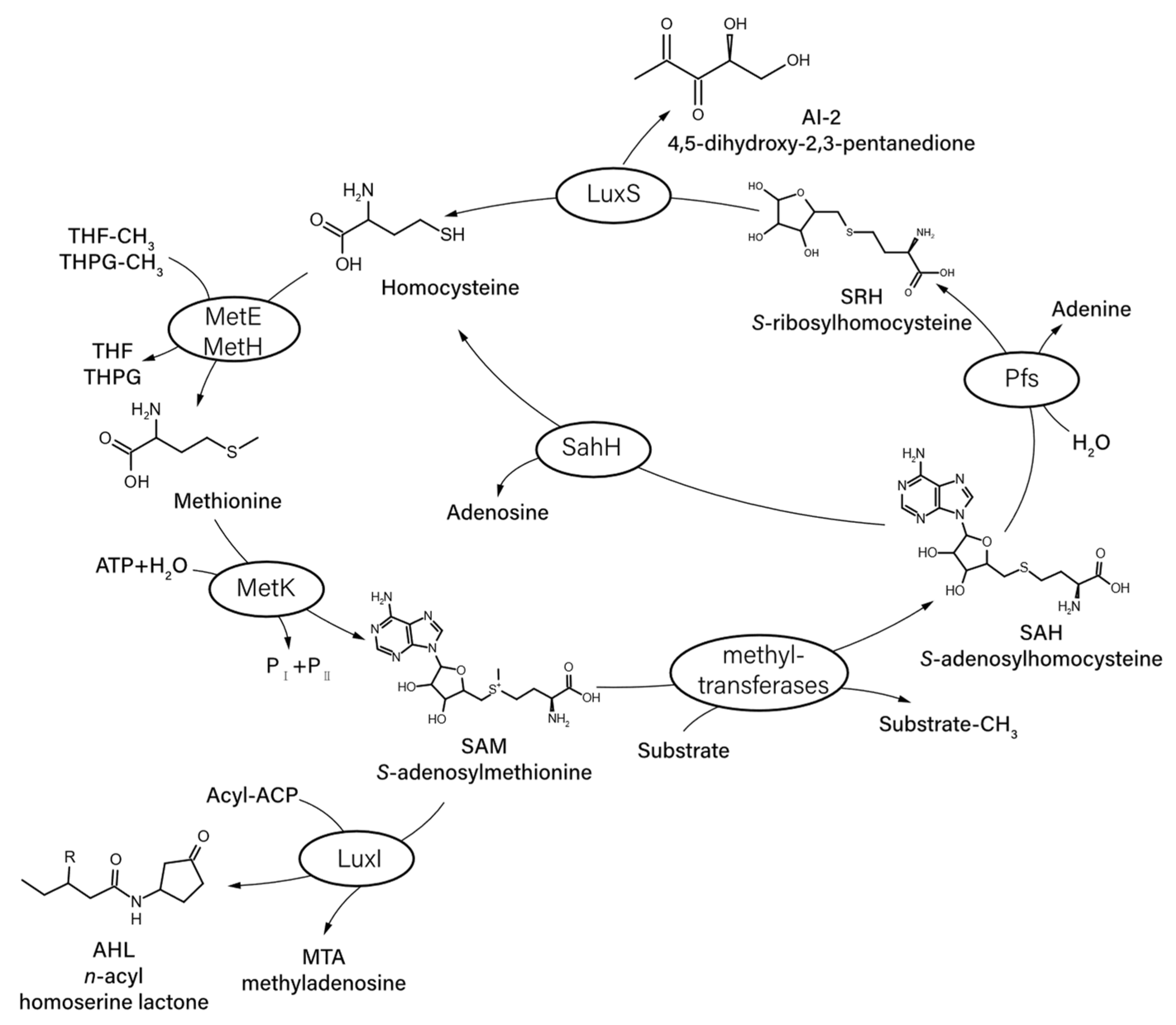
A new class of autoinducers called AI-2 was proposed for observation from the result that V. harveyi mutant strains defective in AHLs synthesis remained capable of QS-dependent gene activation [28]. AI-2 molecules contain a series of interconverting molecules derived from the precursor, 4,5-dihydroxy-2,3-pentanedione (DPD) [19]. The DPD is an unstable molecule that can spontaneously cyclize into various furanone moieties under different conditions [29]. With the rapid conversion of DPD, bacteria detect different forms of DPD as their AI-2 signals. The core gene responsible for AI-2 is luxS. Two other genes of a three-gene operon with luxS are involved in the activated methyl cycle, an essential metabolic pathway for the recycling of SAM [30].
The AI-2-QS serves for both Gram-positive and -negative bacteria. However, it is hard to detect AI-2 in a natural environment due to their variable structures. The first exploration of AI-2 structure was achieved by utilizing the high-affinity nature of the V. harveyi receptor, LuxP, to match the corresponding AI-2 ligand [31]. AI-2 molecules are frequently observed, produced by Vibrio isolated from corals and sponges in the ocean [13]. Using metagenomics, a number of AI-2 homologs were identified from a dinoflagellate bloom and were likely involved in the regulation of symbiotic relationships [8]. The study supported that AI-2 also plays a part in the communication between algae and bacteria.
2.3.2. The Special Role of SAM
Interestingly, SAM serves as the substrate for the synthesis of both AHLs and AI-2. Methyltransferase enzymes and nucleosidase help transfer the methyl group from SAM and form S-ribosylhomocysteine (SRH). Then, SRH is converted to DPD, the linear form of AI-2 (Figure 2) [32]. As for the synthesizing pathway of AHLs, SAM also serves as substrate in combination with acyl carrier proteins (Acyl-ACP). Integrating complex chemical interactions among the bacterial community may be advantageous. The addition of long-chain-length AHLs (C10-, C12- and C14-HSL) caused an increase in the activity of alkaline phosphatases (APases), whereas AI-2 had the opposite effect [33]. One explanation is that a subpopulation may have employed AHLs to upregulate APase activity when it benefited them and AI-2 to downregulate APase activity in situations when overall cell abundances were higher and other competing subpopulations might benefit.
2.4. Other Types of Signaling Molecules
TDA is a structurally unique antibiotic synthesized by marine Roseobacter. The effects of TDA on gene expression patterns in TDA-producing bacteria indicate that TDA can serve as a QS signal [34]. Silicibacter sp. TM1040, a member of Roseobacter, participates in a symbiosis with dinoflagellates, using TDA as their QS signal. Genetic evidence suggests that TDA works in a manner similar to AHLs, and its synthesis requires the expression of tdaABCDEF (tdaA-F) and six additional genes (cysI, malIJK, and tdaH) [35]. Indole-3-ecetic (IAA) is well known as a plant hormone that promotes plant growth via a series of cellular mechanisms. Its potential role in mediating QS pathway was recently reported. A diatom-associated Sulfitobacter species synthesizes IAA from diatom-secreted tryptophan, which enhances diatom cell division [36].
TDA and IAA were previously recognized for their additional functions before being identified as QS signals. This is also applicable for AQs, which were formerly known as antibiotics. Further investigation is required to evaluate the potential chemical diversity in the QS pathway within the phycosphere.
3. Functions in the Phycosphere
The functions regulated by QS substances have been already illustrated in bacterial community, including biofilm formation, virulence factor secretion, bioluminescence, antibiotic production, sporulation and competence for nutrients [37]. In the phycosphere, QS also participates in algae–bacteria interactions and mediates various ecological functions.
3.1. Modulating Microbial Colonization
Chemotaxis is the ability to sense and respond to chemical substances [38]. It allows bacteria to approach favorable chemicals or avoid harmful substances [39]. Algae release a huge amount of dissolved organic matter (DOM) in the phycosphere and attract bacteria [40]. These bacteria are encouraged to switch their free-living lifestyle into colonization of the phycosphere [41]. They benefit from algal DOM and protect algae against antibiotics or toxins [42]. In this process, QS helps inhibit bacterial motility and forms biofilm. For example, several species in the Roseobacter group could attach to diatom transparent exopolymeric particles. Compared to the other strains in the experiment, they hold complete QS systems and synthesize 3-oxo-C16:1-HSL to modulate their settlement in the phycosphere [43]. Nautella italica R11, a member of the Roseobacter clade, contains a unique luxR-type gene, varR, which controls its colonization in the phycosphere and virulence to their host Delisea pulchra. The luxR-type gene knock-out strain was difficult, forming biofilm and causing disease in its host [22].
Biofilm is a microenvironment created by a microorganism, consisting of exopolysaccharides (EPS) [44]. Within the biofilms, QS regulates the production of exopolysaccharides and extracellular hydrolytic enzymes [13]. Red algae Delisea pulchra are special when it comes to resisting colonization of biofilms because they can produce halogenated furanone to inhibit AHL-QS-regulated phenotypes, including biofilms formation [45]. Compared to free-living cells, bacteria in biofilm have greater access to algal nutrients [42]. The QS-regulated abilities in motility and chemotaxis provide bacteria with a competitive advantage when searching for nutrient sources. Then, the formation of biofilm makes the nutrient exchange between bacteria and algae more available. By using periphytic biofilm as the microbial-aggregate model, the pathway of a bacterial phosphorus entrapment has been stimulated, including the up-regulation of pyrimidine, purine and energy metabolism regulated by AHLs [46]. However, the bacterial density in marine biofilms is three times higher than in water [13]. Interactions between cells are enhanced and it is hard to avoid QS crosstalk in the tiny, spatially heterogeneous environment [14]. To successfully achieve cellular communications, bacteria must overcome difficulties in detecting certain signals. For example, AHLs is taxon specific for the structural variations on the acyl side chain. Their receptor protein LuxR contains an amino-terminal ligand-binding domain and a carboxy-terminal DNA-binding domain. LuxR possesses a specific acyl-binding pocket to bind its cognate signal [47]. Some bacteria adopt a strategy that uses unusual signals, such as TDA and indole-3-acetic acid (IAA) [36,48]. The various signal substances help bacteria compete for ecological niches and contribute to modulation of microbial colonization in the phycosphere.
3.2. Achieving Algae–Bacteria Mutualism and Competition in the Phycosphere
The exchanges of nutrients lay the foundation of mutualism between bacteria and their host algae. As mentioned above, bacteria consume a mass of algal dissolved organic matter. In turn, they supply abundant nutrients to support algal growth. A metabolome survey of two strains of Rosoebacter clade showed that bacteria provide vitamins, amino acids and plant growth promotors to algae [49]. Some of these substances are indispensable to algal growth, such as Vitamin B12 (cobalamin), which is required by approximately half of all microalgal species [50]. However, most of the microalgae exhibit vitamin auxotrophy, which can only obtain vitamins from prokaryotes [51]. Picoeukaryotic algae Ostreococcus tauri lack the genetic capacity to grow without Vitamin B12. Through co-culture with heterotrophic bacteria Dinoroseobacter shibae, a member of the Rhodobacteraceae family of alpha-proteobacteria able to synthesize the Vitamin B12, auxotrophy of O.tauri was alleviated [52].
For the purpose of promoting their own growth, algae may nurture particular microbiomes by releasing certain metabolites, such as polyunsaturated fatty acid, eicosapentaenoic acids, polyunsaturated aldehydes [53,54]. The ubiquitous diatom Asterionellopsis glacialis promotes the attachment of beneficial bacteria while suppressing the attachment of opportunists by secreting rosmarinic acid into the phycosphere and uses azelaic acid to enhance the development of beneficial bacteria while inhibiting the growth of opportunists [55]. Interfering QS pathway is another strategy for resisting opportunists and harmful bacteria. Some algae, such as D. pulchra, are able to secret compounds that mimic QS signals, such as halogenated furanones to occupy AHLs binding site of LuxR homologues and inactivate QS [45]. In addition, some algae use enzymes to inhibit QS. Benthic diatom Nitzschia cf pellucida degrades AHLs by cleaving the halogenated N-acyl chain of signal molecules using haloperoxidase [56]. That might explain why various algal phycosphere possesses distinct microbial community compositions. Meanwhile, bacteria can regulate their behavior through QS, or even secrete algicides, in order to maximize the use of algal organic matter. Under the regulation of AHLs, algicidal bacteria γ-proteobacterium produce red pigment (PG-L-1), which shows potent algicidal activity against red tide algae [57]. The same phenomena are common in the process of algal blooms. The algicidal Flavobacterium Kordia algicida isolated from Skeletonema costatum bloom is able to lyse a broad range of algal species, including the diatoms Skeletonema costatum, Thalassiosira weissflogii and Phaeodactylum tricornutum [58]. According to metagenomics examinations, QS autoinducers are ubiquitous in a complete dinoflagellate bloom. The algicidal bacteria do not display algicidal behavior until the population density has reached a certain threshold, suggesting QS controls algicidal activity during this process [8]. Based on the short-time interactions, QS modulates mutualistic or competitive relationships between algae and bacteria.
3.3. Contributing to Marine Biogeochemical Cycles
The significant role of algae and their associated bacteria in marine aquatic food web has already been addressed [59]. At present, the in-depth study of the bacterial QS mechanism makes us realize the importance of these microscale interactions. One important aspect is that QS regulates the activities of extracellular hydrolytic enzymes. This function was first identified from bacteria-colonized marine snow particles [12]. A similar observation was carried out on heterotrophic bacteria of Trichodesmiun. These strains use alkaline phosphatases (APases) to acquire phosphate from dissolved organic phosphorus. The addition of AHLs increased the activity of bacterial APase, while the addition of AI-2 showed the opposite effect [33].
For the single bacterial species, their bioactivity in acquiring nutrients changes in response to the host state. Dimethyl-sulfoniopropionate (DMSP) are the most conspicuous molecules produced by some algal species and are deemed as the principal source of organic sulfur in the marine environment. It also carries the functions inducing bacteria to algal cells and stimulating bacterial production of QS molecules [60]. Roseobacter Phaeobacter gallaeciensis symbiosed with algae Emiliania huxleyi are able to degrade DMSP into dimethyl sulfide (DMA). DMA further reacts within the atmosphere, where it is easily oxidized to sulfate aerosols, which increase the abundance of cloud condensation and mitigate the impacts of global warming [61]. During E. huxleyi bloom, DMSP activates the normally silent biosynthesis pathway in P. gallaeciensis to produce TDA that helps algae resist harmful bacteria. As the bloom fades, bacteria consume algal breakdown products and use them to biosynthesize QS signals pCA. In the regulation of pCA, P. gallaeciensis elicit a powerful algicide, roseobacticides, to induce algal apoptosis [21]. These dynamic metabolic processes speed up the bloom fading and accelerate the matter circulation.
DMSP degradation dominated by Roseobacter clade plays an essential ecological role in marine sulfur cycles. Roseobacter members are the most abundant bacteria in the phycosphere, accounting for about 15–50% of the entire planktonic bacterial community, according to prior study [62]. Moreover, Roseobacter are the main marine DMSP lytic bacteria, with 12 species capable of degrading and metabolizing sulfides [48]. The concentration of DMSP increases, accompanied by algal growth, and QS bacteria degrade DMSP to produce DMS. DMS further helps with the formation of clouds, which has an impact on algal photosynthesis [63]. QS-based regulation in tiny-scale interactions might be crucial in marine biogeochemical cycles. QS coordinates the interactions between bacterial communities and algae to jointly realize ecological processes. Furthermore, this communication mechanism is widespread in the ocean. However, there is still a lot to learn about QS functions in marine biogeochemical cycles.
4. Methods in Study: Detection and Functional Evaluation
The conventional workflow of detecting QS signals in the phycosphere is using biosensors. Take AHLs detection, for example; the biosensors refer to genetically modified bacteria with defective LuxI synthase, which is dependent on the LuxR protein in demonstrating selectivity binding towards the cognate AHLs capable of activating reporter-gene transcription [64]. These mutants encode the purple pigment violacein or the Green Fluorescent Protein (GFP) and luminesce when exposed to AHLs [3]. After isolating from phycosphere, the expression of bacterial AHLs can be detected by biosensors [65]. In the early stages, the functions of QS signals were commonly evaluated by adding exogenous metabolites. For example, after adding exogenous AHLs and AI-2 molecules into the cyanobacterium Trichodesmium culture, the activity of bacterial APase showed an obvious fluctuation [33].
However, biosensors can commonly only detect a narrow range of signals, so this method is not suitable for quantification [66]. Given this limitation, signals detection by bacterial reporter strains is always in combination with GC-MS and LC-MS methods. With the development of genomics and metabolomics approaches, the presence of AHLs biosynthesis genes in metagenomes can indicate a genetic capability for producing signals. Meanwhile, the development of transcriptomic and proteomic approaches and settlement of model strains has contributed to the exploration of QS-regulated traits. The majority of research has relied on model strains, in which key genes involved in QS are mutated, in particular the lux-like genes. Through establishing the gene knock-out strain, the role of a unique luxR-type gene in Roseobacter clade has been clarified: it controls bacterial colonization and virulence by hindering forming biofilm [22]. In future research, improving the accuracy of detection in signal molecules and analyzing the dynamic variation in community are important.
5. Potential Applications
5.1. Potential Applications in Controlling HABs
Harmful algal blooms (HABs) refer to algal blooms which accompany natural toxin production, dissolved oxygen depletion or other negative impacts [67]. HABs occur worldwide in marine, estuarine and freshwater ecosystems as a considerable threat to human health [68]. Currently, physical and chemical approaches are often utilized to control HABs. These treatments can be effective in a short period of time, but they can generate secondary contamination. Given that limitation, it is necessary to develop environmentally friendly biological treatment methods.
Through screening and detecting the QS bacteria population during the Scrippsiella trochoidea blooming period, it was found that the bacterial dynamic characteristics were correlated with the blooming stage [69]. Meanwhile, the bacteria in the phycosphere provide some of necessary nutrients for host algae. Developing QS inhibitors to interfere bacterial biological functions might be a promising option in biological control of HABs. It was actually proven to be feasible by laboratory work. Hong et al. inhabited the growth of Microcystis aeruginosa by adding QS inhibitor ethyl2-methylacetoacetate (EMA) [70]. Nakashima et al. confirmed that pigment produced by γ-proteobacterium was controlled by QS and showed potent algicidal activity against red tide algae [57]. The QS-based method is more attractive than traditional methods, since it avoids secondary contamination created by chemical reagents. However, there are two problems that need to be explored. First of all, there is no successful ecological application case so far, and the research it is still in the laboratory. As a result, the effectiveness of the application in a natural setting is remains unknown. Second, the mechanism of its activity needs to be further studied, including the consequences of exogenous QS signals on microbial response and whether microbial contamination will be caused.
5.2. Potential Applications in Production of Biofuel
In response to the rising energy needs, a variety of alternative energy sources such as solar, wind and geothermal have been researched and implemented [71]. Microalgae are regarded as an excellent alternative for fossil fuels among renewable energy sources. Unlike terrestrial crops grown for biofuel, marine algae do not require agricultural land for cultivation and hence avoid competing for the land and fresh water needed for food production [72]. Current research focuses mostly on the cultivation of microalgae, utilizing wastewater, genetic engineering modification of microalgae species and optimization of nutritional conditions [73]. Meanwhile, harnessing the effect of bacterial QS on microalgae has emerged as a viable technique for increasing microalgae biomass accumulation. Classes such as Chlorophyta are able to receive, sense and respond to bacterial AHLs, in the way that AHLs induce bacterial creation of aromatic proteins, which subsequently influences cell aggregation [74]. Furthermore, three bacteria strains with AHLs-secreting activity from Chlorella culture significantly increase the polysaccharides and proteins synthesis in microalgae cells, which ultimately stimulates the microalgae biomass accumulation [75]. C14-HSL can upregulate genes involved in intracellular signaling, and oxo-C14-HSL can shift lipid metabolism toward enhanced fatty acid degradation while downregulating cell-cycle genes. As a result, employing AHLs secretory bacteria or introducing exogenous AHLs may help promote algal growth and metabolism, thus increasing biofuel production. However, further research is needed to determine their effectiveness, stability, and degradability in water bodies. At the same time, the molecular mechanism of biofuel production via the algae bacteria system should be revealed [75].
6. Conclusions and Perspectives
Bacterial species that live in the marine phycosphere are commonly within specific classes. Both algae and bacteria are responsible for this phenomenon. Firstly, chemotaxis results in bacteria colonizing and populating in the phycosphere, using the nutrients released by algae and secreting vitamin B and other substances to support algae growth. Algae can identify colonizing bacteria as harmful or beneficial and stimulate the colonization and proliferation of beneficial bacteria. The ecological role of quorum sensing as a bacterial communication language in the phycosphere is not well understood. At present, it has been confirmed that its functions include regulating bacterial-population-level biological activities, promoting the formation of biofilm and ensuring that bacteria maximize access and utilization of nutrients. Meanwhile, QS signal molecules can be directly detected by host algae as a means of regulating dynamic population changes in the phycosphere. QS thus indirectly affects the nutrient exchange between algae and bacteria, and then has an impact on the marine biogeochemical cycle. To summarize, QS, as a small-scale biological activity mediated by diverse substances, has a complicated mechanism of action yet has significant ecological impact. These findings may spark fresh research into the interaction of algae and bacteria in the marine phycosphere. Future work can be carried out from the following aspects:
- 1.
- More research is needed to develop techniques for detection and isolation of signals. A large number of AHLs compounds have been discovered in the phycosphere, while other QS systems have been much less described. Omics techniques are promising for future study, especially the comparative proteomics and metabolomics, which may help to discover new signals. Moreover, it is difficult to detect and quantify the molecules in water directly due to their low concentration and chemical stability. A recent study mentions a progress in AHLs quantification from environmental samples. After flash freezing and solvent extraction, samples were analyzed with an ultra-high-performance liquid chromatography-high resolution mass spectrometry (UHPLC-HRMS) method; AHLs are quantified in high sensitivity. More research into identifying low-concentration QS signals in seawater could help us better understand the algae–bacteria interactions.
- 2.
- More model bacteria–algae systems are required for future study. Many laboratory model systems have confirmed functions regulated by QS signals so far. However, there are only a few strains that can be studied in the lab. Developing new co-culture systems could open up new avenues for research into the biological effects of QS.
- 3.
- Climate influences on QS pathway need to be further explored. Since preindustrial times, CO2 concentration in the atmosphere has increased from ~280 ppm to ~400 ppm today. According to the estimation, anthropogenic CO2 emissions will be projected to lower the pH of the ocean by 0.3 units by 2100. Since the structural integrity of AHLs is susceptible to pH, the fluctuations in pH may result in AHLs hydrolysis. It may have serious consequence among bacteria as well as marine algae. Zoospores of macroalga Ulva detect AHLs to identify bacterial biofilms for preferential settlement. AHLs triggers intracellular Ca2+ in zoospores cells, resulting in a decrease in swimming speed. QS signals are used by organisms as a part of their environmental sensing capacity; the changes in marine chemical factors may have profound consequences for both QS-bacterial and their associated algae.
In conclusion, natural products for QS in marine phycosphere are diverse. Research on them may allow us to better understand their contribution to shape algae–bacteria partnerships and antagonisms. These microscale interactions are further involved in large-scale biogeochemical processes. Future studies should be oriented towards research on the natural matrices mediating the QS process in order to deeply understand algae–bacteria interactions.
Author Contributions
Writing—original draft preparation, Z.Q.; writing—review and editing, J.L.; visualization, Z.Q.; supervision, S.Q.; funding acquisition, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (Grant No. 2018YFD0901102), the National Natural Science Foundation of China (Grant No. 42176131) and the Key Deployment Project of Centre for Ocean Mega-Research of Science, Chinese Academy of Science (Grant No. COMS2020Q09).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cai, Z.; Zhu, W.; Zeng, Y.; Zhou, J. Advances in extracellular polymeric substances in phycosphere environment. Weishengwu Xuebao 2020, 60, 1521–1533. [Google Scholar]
- Rolland, J.L.; Stien, D.; Sanchez-Ferandin, S.; Lami, R. Quorum Sensing and Quorum Quenching in the Phycosphere of Phytoplankton: A Case of Chemical Interactions in Ecology. J. Chem. Ecol. 2016, 42, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, R.; Kim, B.-H.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Algae-bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demuth, D.R.; Lamont, R.J. Quorum Sensing and Cell-to-Cell Communication in the Dental Biofilm; Cambridge University Press: Cambridge, UK, 2006; pp. 175–197. [Google Scholar]
- Bachofen, R.; Schenk, A. Quorum sensing autoinducers: Do they play a role in natural microbial habitats? Microbiol. Res. 1998, 153, 61–63. [Google Scholar] [CrossRef]
- Wagner-Dobler, I.; Thiel, V.; Eberl, L.; Allgaier, M.; Bodor, A.; Meyer, S.; Ebner, S.; Hennig, A.; Pukall, R.; Schulz, S. Discovery of complex mixtures of novel long-chain quorum sensing signals in free-living and host-associated marine alphaproteobacteria. Chembiochem 2005, 6, 2195–2206. [Google Scholar] [CrossRef]
- Huang, X.; Zhu, J.; Cai, Z.; Lao, Y.; Jin, H.; Yu, K.; Zhang, B.; Zhou, J. Profiles of quorum sensing (QS)-related sequences in phycospheric microorganisms during a marine dinoflagellate bloom, as determined by a metagenomic approach. Microbiol. Res. 2018, 217, 1–13. [Google Scholar] [CrossRef]
- Wheeler, G.L.; Tait, K.; Taylor, A.; Brownlee, C.; Joint, I. Acyl-homoserine lactones modulate the settlement rate of zoospores of the marine alga Ulva intestinalis via a novel chemokinetic mechanism. Plant Cell Environ. 2006, 29, 608–618. [Google Scholar] [CrossRef]
- Brown, E.R.; Cepeda, M.R.; Mascuch, S.J.; Poulson-Ellestad, K.L.; Kubanek, J. Chemical ecology of the marine plankton. Nat. Prod. Rep. 2019, 36, 1093–1116. [Google Scholar] [CrossRef]
- Garrido, A.G.; Machado, L.F.; Zilberberg, C.; Leite, D.C.d.A. Insights into ‘Symbiodiniaceae phycosphere’ in a coral holobiont. Symbiosis 2021, 83, 25–39. [Google Scholar] [CrossRef]
- Hmelo, L.R.; Mincer, T.J.; Van Mooy, B.A.S. Possible influence of bacterial quorum sensing on the hydrolysis of sinking particulate organic carbon in marine environments. Environ. Microbiol. Rep. 2011, 3, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Hmelo, L.R.; Annual, R. Quorum Sensing in Marine Microbial Environments. In Annual Review of Marine Science; Annual Review: Palo Alto, CA, USA, 2017; Volume 9, pp. 257–281. [Google Scholar]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Hense, B.A.; Kuttler, C.; Mueller, J.; Rothballer, M.; Hartmann, A.; Kreft, J.-U. Opinion-Does efficiency sensing unify diffusion and quorum sensing? Nat. Rev. Microbiol. 2007, 5, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.; Belas, R. Expression of Tropodithietic Acid Biosynthesis Is Controlled by a Novel Autoinducer. J. Bacteriol. 2010, 192, 4377–4387. [Google Scholar] [CrossRef] [Green Version]
- Engebrecht, J.; Nealson, K.; Silverman, M. Bacterial Bioluminescence-Isolation and Genetic-Analysis of Functions from Vibrio Fischeri. Cell 1983, 32, 773–781. [Google Scholar] [CrossRef]
- Eberhard, A.; Burlingame, A.L.; Eberhard, C.; Kenyon, G.L.; Nealson, K.H.; Oppenheimer, N.J. Structural Identification of Autoinducer of Photobacterium-Fischeri Luciferase. Biochemistry 1981, 20, 2444–2449. [Google Scholar] [CrossRef]
- Galloway, W.R.J.D.; Hodgkinson, J.T.; Bowden, S.D.; Welch, M.; Spring, D.R. Quorum Sensing in Gram-Negative Bacteria: Small-Molecule Modulation of AHL and Al-2 Quorum Sensing Pathways. Chem. Rev. 2011, 111, 28–67. [Google Scholar] [CrossRef]
- Schaefer, A.L.; Greenberg, E.P.; Oliver, C.M.; Oda, Y.; Huang, J.J.; Bittan-Banin, G.; Peres, C.M.; Schmidt, S.; Juhaszova, K.; Sufrin, J.R.; et al. A new class of homoserine lactone quorum-sensing signals. Nature 2008, 454, 595–599. [Google Scholar] [CrossRef]
- Seyedsayamdost, M.R.; Case, R.J.; Kolter, R.; Clardy, J. The Jekyll-and-Hyde chemistry of Phaeobacter gallaeciensis. Nat. Chem. 2011, 3, 331–335. [Google Scholar] [CrossRef]
- Gardiner, M.; Fernandes, N.D.; Nowakowskil, D.; Raftery, M.; Kjelleberg, S.; Zhong, L.; Thomas, T.; Egan, S. VarR controls colonization and virulence in the marine macroalgal pathogen Nautella Italica R11. Front. Microbiol. 2015, 6, 1130. [Google Scholar] [CrossRef] [Green Version]
- Heeb, S.; Fletcher, M.P.; Chhabra, S.R.; Diggle, S.P.; Williams, P.; Camara, M. Quinolones: From antibiotics to autoinducers. FEMS Microbiol. Rev. 2011, 35, 247–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.-l.; Cui, Y.-b.; Cao, S.-h. Progress in research on quinolone antibacterials. Zhongguo Kangshengsu Zazhi 2011, 36, 255–263. [Google Scholar]
- Deziel, E.; Lepine, F.; Milot, S.; He, J.X.; Mindrinos, M.N.; Tompkins, R.G.; Rahme, L.G. Analysis of Pseudomonas aeruginosa 4-hydroxy-2-alkylquinolines (HAQs) reveals a role for 4-hydroxy-2-heptylquinoline in cell-to-cell communication. Proc. Natl. Acad. Sci. USA 2004, 101, 1339–1344. [Google Scholar] [CrossRef] [Green Version]
- Whalen, K.E.; Becker, J.W.; Schrecengost, A.M.; Gao, Y.; Giannetti, N.; Harvey, E.L. Bacterial alkylquinolone signaling contributes to structuring microbial communities in the ocean. Microbiome 2019, 7, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dow, L.; Stock, F.; Peltekis, A.; Szamosvari, D.; Prothiwa, M.; Lapointe, A.; Boettcher, T.; Bailleul, B.; Vyverman, W.; Kroth, P.G.; et al. The Multifaceted Inhibitory Effects of an Alkylquinolone on the Diatom Phaeodactylum tricornutum. Chembiochem 2020, 21, 1206–1216. [Google Scholar] [CrossRef]
- Bassler, B.L.; Wright, M.; Showalter, R.E.; Silverman, M.R. Intercellular Signaling in Vibrio-Harveyi-Sequence and Function of Genes Regulating Expression of Luminescence. Mol. Microbiol. 1993, 9, 773–786. [Google Scholar] [CrossRef]
- Pereira, C.S.; Thompson, J.A.; Xavier, K.B. AI-2-mediated signalling in bacteria. Fems Microbiol. Rev. 2013, 37, 156–181. [Google Scholar] [CrossRef] [Green Version]
- Schauder, S.; Shokat, K.; Surette, M.G.; Bassler, B.L. The LuxS family of bacterial autoinducers: Biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 2001, 41, 463–476. [Google Scholar] [CrossRef]
- Chen, X.; Schauder, S.; Potier, N.; Van Dorsselaer, A.; Pelczer, I.; Bassler, B.L.; Hughson, F.M. Structural identification of a bacterial quorum-sensing signal containing boron. Nature 2002, 415, 545–549. [Google Scholar] [CrossRef]
- Winzer, K.; Hardie, K.R.; Burgess, N.; Doherty, N.; Kirke, D.; Holden, M.T.G.; Linforth, R.; Cornell, K.A.; Taylor, A.J.; Hill, P.J.; et al. LuxS: Its role in central metabolism and the in vitro synthesis of 4-hydroxy-5-methyl-3(2H)-furanone. Microbiol.-Sgm 2002, 148, 909–922. [Google Scholar] [CrossRef] [Green Version]
- Van Mooy, B.A.S.; Hmelo, L.R.; Sofen, L.E.; Campagna, S.R.; May, A.L.; Dyhrman, S.T.; Heithoff, A.; Webb, E.A.; Momper, L.; Mincer, T.J. Quorum sensing control of phosphorus acquisition in Trichodesmium consortia. ISME J. 2012, 6, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, N.; Lindqvist, L.L.; Wibowo, M.; Sonnenschein, E.C.; Bentzon-Tilia, M.; Gram, L. Role is in the eye of the beholder-the multiple functions of the antibacterial compound tropodithietic acid produced by marine Rhodobacteraceae. FEMS Microbiol. Rev. 2022, 46, fuac007. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.F.; Belas, R. TdaA Regulates Tropodithietic Acid Synthesis by Binding to the tdaC Promoter Region. J. Bacteriol. 2011, 193, 4002–4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, S.A.; Hmelo, L.R.; van Tol, H.M.; Durham, B.P.; Carlson, L.T.; Heal, K.R.; Morales, R.L.; Berthiaume, C.T.; Parker, M.S.; Djunaedi, B.; et al. Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature 2015, 522, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.-L.; Bassler, B.L. Bacterial Quorum-Sensing Network Architectures. Annu. Rev. Genet. 2009, 43, 197–222. [Google Scholar] [CrossRef] [Green Version]
- Raina, J.B.; Fernandez, V.; Lambert, B.; Stocker, R.; Seymour, J.R. The role of microbial motility and chemotaxis in symbiosis. Nat. Rev. Microbiol. 2019, 17, 284–294. [Google Scholar] [CrossRef]
- Lux, R.; Shi, W.Y. Chemotaxis-guided movements in bacteria. Crit. Rev. Oral Biol. Med. 2004, 15, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Smriga, S.; Fernandez, V.I.; Mitchell, J.G.; Stocker, R. Chemotaxis toward phytoplankton drives organic matter partitioning among marine bacteria. Proc. Natl. Acad. Sci. USA 2016, 113, 1576–1581. [Google Scholar] [CrossRef] [Green Version]
- Mayali, X.; Franks, P.J.S.; Burton, R.S. Temporal attachment dynamics by distinct bacterial taxa during a dinoflagellate bloom. Aquat. Microb. Ecol. 2011, 63, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Samo, T.J.; Kimbrel, J.A.; Nilson, D.J.; Pett-Ridge, J.; Weber, P.K.; Mayali, X. Attachment between heterotrophic bacteria and microalgae influences symbiotic microscale interactions. Environ. Microbiol. 2018, 20, 4385–4400. [Google Scholar] [CrossRef]
- Fei, C.; Ochsenkuhn, M.A.; Shibl, A.A.; Isaac, A.; Wang, C.; Amin, S.A. Quorum sensing regulates ‘swim-or-stick’ lifestyle in the phycosphere. Environ. Microbiol. 2020, 22, 4761–4778. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G. Microbial Colonization in Marine Environments: Overview of Current Knowledge and Emerging Research Topics. J. Mar. Sci. Eng. 2020, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Manefield, M.; Welch, M.; Givskov, M.; Salmond, G.P.C.; Kjelleberg, S. Halogenated furanones from the red alga, Delisea pulchra, inhibit carbapenem antibiotic synthesis and exoenzyme virulence factor production in the phytopathogen Erwinia carotovora. Fems Microbiol. Lett. 2001, 205, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Curtis, T.; Dolfing, J.; Wu, Y.; Rittmann, B.E. N-acyl-homoserine-lactones signaling as a critical control point for phosphorus entrapment by multi-species microbial aggregates. Water Res. 2021, 204, 117627. [Google Scholar] [CrossRef]
- Vannini, A.; Volpari, C.; Gargioli, C.; Muraglia, E.; Cortese, R.; De Francesco, R.; Neddermann, P.; Di Marco, S. The crystal structure of the quorum sensing protein TraR bound to its autoinducer and target DNA. Embo J. 2002, 21, 4393–4401. [Google Scholar] [CrossRef]
- Majzoub, M.E.; McElroy, K.; Maczka, M.; Schulz, S.; Thomas, T.; Egan, S. Genomic Evolution of the Marine Bacterium Phaeobacter inhibens during Biofilm Growth. Appl. Environ. Microbiol. 2021, 87, e00769-21. [Google Scholar] [CrossRef]
- Wienhausen, G.; Noriega-Ortega, B.E.; Niggemann, J.; Dittmar, T.; Simon, M. The Exometabolome of Two Model Strains of the Roseobacter Group: A Marketplace of Microbial Metabolites. Front. Microbiol. 2017, 8, 1985. [Google Scholar] [CrossRef]
- Helliwell, K.E.; Wheeler, G.L.; Leptos, K.C.; Goldstein, R.E.; Smith, A.G. Insights into the Evolution of Vitamin B-12 Auxotrophy from Sequenced Algal Genomes. Mol. Biol. Evol. 2011, 28, 2921–2933. [Google Scholar] [CrossRef] [Green Version]
- Scott, A.I.; Roessner, C.A. Biosynthesis of cobalamin (vitamin B-12). Biochem. Soc. Trans. 2002, 30, 613–620. [Google Scholar] [CrossRef]
- Cooper, M.B.; Kazamia, E.; Helliwell, K.E.; Kudahl, U.J.; Sayer, A.; Wheeler, G.L.; Smith, A.G. Cross-exchange of B-vitamins underpins a mutualistic interaction between Ostreococcus tauri and Dinoroseobacter shibae. ISME J. 2019, 13, 334–345. [Google Scholar] [CrossRef] [Green Version]
- Seymour, J.R.; Amin, S.A.; Raina, J.-B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton-bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lyu, Y.; Richlen, M.L.; Anderson, D.M.; Cai, Z. Quorum Sensing Is a Language of Chemical Signals and Plays an Ecological Role in Algal-Bacterial Interactions. Crit. Rev. Plant Sci. 2016, 35, 81–105. [Google Scholar] [CrossRef] [PubMed]
- Shibl, A.A.; Isaac, A.; Ochsenkuhn, M.A.; Cardenas, A.; Fei, C.; Behringer, G.; Arnoux, M.; Drou, N.; Santos, M.P.; Gunsalus, K.C.; et al. Diatom modulation of select bacteria through use of two unique secondary metabolites. Proc. Natl. Acad. Sci. USA 2020, 117, 27445–27455. [Google Scholar] [CrossRef] [PubMed]
- Syrpas, M.; Ruysbergh, E.; Blommaert, L.; Vanelslander, B.; Sabbe, K.; Vyverman, W.; De Kimpe, N.; Mangelinckx, S. Haloperoxidase Mediated Quorum Quenching by Nitzschia cf pellucida: Study of the Metabolization of N-Acyl Homoserine Lactones by a Benthic Diatom. Mar. Drugs 2014, 12, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, T.; Miyazaki, Y.; Matsuyama, Y.; Muraoka, W.; Yamaguchi, K.; Oda, T. Producing mechanism of an algicidal compound against red tide phytoplankton in a marine bacterium gamma-proteobacterium. Appl. Microbiol. Biotechnol. 2006, 73, 684–690. [Google Scholar] [CrossRef]
- Paul, C.; Pohnert, G. Interactions of the Algicidal Bacterium Kordia algicida with Diatoms: Regulated Protease Excretion for Specific Algal Lysis. PLoS ONE 2011, 6, e21032. [Google Scholar] [CrossRef]
- Amin, S.A.; Parker, M.S.; Armbrust, E.V. Interactions between Diatoms and Bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 667. [Google Scholar] [CrossRef] [Green Version]
- Cirri, E.; Pohnert, G. Algae-bacteria interactions that balance the planktonic microbiome. New Phytol. 2019, 223, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Geng, H.; Belas, R. Molecular mechanisms underlying roseobacter-phytoplankton symbioses. Curr. Opin. Biotechnol. 2010, 21, 332–338. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, L.; Wang, S.; Zhao, Y.; Xu, X.; Han, B.; Hu, T. Quorum Sensing Bacteria in the Phycosphere of HAB Microalgae and Their Ecological Functions Related to Cross-Kingdom Interactions. Int. J. Environ. Res. Public Health 2022, 19, 163. [Google Scholar] [CrossRef]
- Saint-Macary, A.D.; Barr, N.; Armstrong, E.; Safi, K.; Marriner, A.; Gall, M.; McComb, K.; Dillingham, P.W.; Law, C.S. The Influence of Ocean Acidification and Warming on DMSP & DMS in New Zealand Coastal Water. Atmosphere 2021, 12, 181. [Google Scholar]
- Tan, W.-S.; Yunos, N.Y.M.; Tan, P.-W.; Mohamad, N.I.; Adrian, T.-G.-S.; Yin, W.-F.; Chan, K.-G. Characterisation of a Marine Bacterium Vibrio Brasiliensis T33 Producing N-acyl Homoserine Lactone Quorum Sensing Molecules. Sensors 2014, 14, 12104–12113. [Google Scholar] [CrossRef] [PubMed]
- Daly, G.; Perrin, E.; Viti, C.; Fondi, M.; Adessi, A. Scaling down the microbial loop: Data-driven modelling of growth interactions in a diatom-bacterium co-culture. Environ. Microbiol. Rep. 2021, 13, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chai, Y.R.; Zhong, Z.T.; Li, S.P.; Winans, S.C. Agrobacterium bioassay strain for ultrasensitive detection of N-acylhomoserine lactone-type quorum-sensing molecules: Detection of autoinducers in Mesorhizobium huakuii. Appl. Environ. Microbiol. 2003, 69, 6949–6953. [Google Scholar] [CrossRef] [Green Version]
- Zohdi, E.; Abbaspour, M. Harmful algal blooms (red tide): A review of causes, impacts and approaches to monitoring and prediction. Int. J. Environ. Sci. Technol. 2019, 16, 1789–1806. [Google Scholar] [CrossRef]
- Patel, S.S.; Lovko, V.J.; Lockey, R.F. Red Tide: Overview and Clinical Manifestations. J. Allergy Clin. Immunol. -Pract. 2020, 8, 1219–1223. [Google Scholar] [CrossRef]
- Lv, H.; Zhou, J.; Cai, Z. Dynamic variation of quorum sensing bacteria in a Scrippsiella trochoidea bloom. Ecol. Sci. 2016, 35, 23–30. [Google Scholar]
- Merlo, S.; Durany, X.G.; Tonon, A.P.; Rossi, S. Marine Microalgae Contribution to Sustainable Development. Water 2021, 13, 1373. [Google Scholar] [CrossRef]
- Milledge, J.J.; Smith, B.; Dyer, P.W.; Harvey, P. Macroalgae-Derived Biofuel: A Review of Methods of Energy Extraction from Seaweed Biomass. Energies 2014, 7, 7194–7222. [Google Scholar] [CrossRef]
- Maeda, Y.; Yoshino, T.; Matsunaga, T.; Matsumoto, M.; Tanaka, T. Marine microalgae for production of biofuels and chemicals. Curr. Opin. Biotechnol. 2018, 50, 111–120. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Guo, G.; Pu, Y.; Yan, B. The isolation and antioxidant activity of polysaccharides from the marine microalgae Isochrysis galbana. Carbohydr. Polym. 2014, 113, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Sun, W.; Su, Y.; Ren, Q.; Ji, Z.; Zhang, A. Utilization of N-Acyl Homoserine Lactone-Secreting Bacteria in Algal Environment to Increase Biomass Accumulation of Chlorella. Bioenergy Res. 2022, 5. [Google Scholar] [CrossRef]
- Stock, F.; Bilcke, G.; De Decker, S.; Osuna-Cruz, C.M.; Van den Berge, K.; Vancaester, E.; De Veylder, L.; Vandepoele, K.; Mangelinckx, S.; Vyverman, W. Distinctive Growth and Transcriptional Changes of the DiatomSeminavis robustain Response to Quorum Sensing Related Compounds. Front. Microbiol. 2020, 11, 1240. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The mechanism of quorum sensing in Gram-negative bacteria: (a) At low bacterial cell density, LuxI catalyzes the synthesis of AHLs and then AHLs diffuse into the extracellular environment; (b) when bacteria are at high cell-density, AHLs diffuse into bacterial cell and bind to the receptor LuxR, and thus activate the transcription of genes under QS control.
Figure 1.
The mechanism of quorum sensing in Gram-negative bacteria: (a) At low bacterial cell density, LuxI catalyzes the synthesis of AHLs and then AHLs diffuse into the extracellular environment; (b) when bacteria are at high cell-density, AHLs diffuse into bacterial cell and bind to the receptor LuxR, and thus activate the transcription of genes under QS control.

Figure 2.
Diagram illustrating the AI-2 and AHLs biosynthetic pathway (modified from [29]). SAM is transformed into SRH via a series of reactions, and SRH is subsequently converted to DPD, the linear form of AI-2. SAM also serves as substrate with Acyl-ACP for the synthesis of AHLs.
Figure 2.
Diagram illustrating the AI-2 and AHLs biosynthetic pathway (modified from [29]). SAM is transformed into SRH via a series of reactions, and SRH is subsequently converted to DPD, the linear form of AI-2. SAM also serves as substrate with Acyl-ACP for the synthesis of AHLs.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qiao, Z.; Li, J.; Qin, S. Bioactive Compounds for Quorum Sensing Signal-Response Systems in Marine Phycosphere. J. Mar. Sci. Eng. 2022, 10, 699. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050699
AMA Style
Qiao Z, Li J, Qin S. Bioactive Compounds for Quorum Sensing Signal-Response Systems in Marine Phycosphere. Journal of Marine Science and Engineering. 2022; 10(5):699. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050699
Chicago/Turabian StyleQiao, Zhen, Jialin Li, and Song Qin. 2022. "Bioactive Compounds for Quorum Sensing Signal-Response Systems in Marine Phycosphere" Journal of Marine Science and Engineering 10, no. 5: 699. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse10050699
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.