
Effects of the Marine Biotoxins Okadaic Acid and Dinophysistoxins on Fish


1
IPMA—Portuguese Institute for the Sea and Atmosphere, Av. Brasília, 1449-006 Lisbon, Portugal
2
CIRSA—Centro Interdipartimentale di Ricerca per le Scienze Ambientali, Università di Bologna, Via Sant’Alberto, 163-48100 Ravenna, Italy
3
CCMAR—Centre of Marine Sciences, University of Algarve, Campus of Gambelas, 8005-139 Faro, Portugal
*
Author to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2021, 9(3), 293; https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9030293
Submission received: 25 January 2021
/
Revised: 28 February 2021
/
Accepted: 1 March 2021
/
Published: 7 March 2021
(This article belongs to the Section Marine Biology)
Abstract
:Natural high proliferations of toxin-producing microorganisms in marine and freshwater environments result in dreadful consequences at the socioeconomically and environmental level due to water and seafood contamination. Monitoring programs and scientific evidence point to harmful algal blooms (HABs) increasing in frequency and intensity as a result of global climate alterations. Among marine toxins, the okadaic acid (OA) and the related dinophysistoxins (DTX) are the most frequently reported in EU waters, mainly in shellfish species. These toxins are responsible for human syndrome diarrhetic shellfish poisoning (DSP). Fish, like other marine species, are also exposed to HABs and their toxins. However, reduced attention has been given to exposure, accumulation, and effects on fish of DSP toxins, such as OA. The present review intends to summarize the current knowledge of the impact of DSP toxins and to identify the main issues needing further research. From data reviewed in this work, it is clear that exposure of fish to DSP toxins causes a range of negative effects, from behavioral and morphological alterations to death. However, there is still much to be investigated about the ecological and food safety risks related to contamination of fish with DSP toxins.
1. Introduction
Harmful algal blooms (HABs) are a natural phenomenon that occurs at all latitudes and in all aquatic environments (freshwater, brackish, and marine). During the last decades they appear to be emerging in terms of frequency, duration, and severity throughout the world. Such increase is in part related to awareness and improved monitoring programs of HABs [1], but scientific evidence also indicates that the increase in HABs may be correlated with meso and large-scale physico-chemical processes [2]. Nevertheless, the main drivers are attributed to natural environmental factors (intensification or weakening of local nutrient upwelling, storms events), climate change (increasing temperature, enhanced surface stratification, alteration of ocean currents, stimulation of photosynthesis by elevated CO2), and anthropogenic activity represented by sources of nutrients such as sewage and animal wastes, agricultural and fertilizer runoff, as well as the growing aquaculture industry in coastal areas [3,4,5]. Moreover, HAB events can occur after complex interactions between those factors. Recent evidence suggests that the interactive effects of eutrophication and climate change could increase the likelihood of HABs occurrence and severity [6].
Small marine pelagic fish, such as sardines and anchovies, that are planktivorous filter-feeders have been previously identified as vectors of marine biotoxins (domoic acid) in the trophic chain during blooms of toxic algae [7,8]. Indeed, several poisoning outbreaks with massive mortalities of marine top predators, such as marine mammals and seabirds, have been reported worldwide after ingestion of small pelagic fish containing high levels of marine biotoxins, namely saxitoxin and derivatives [9]. In addition to planktivorous filter-feeders, fish may also be exposed to marine toxins due to intense feeding on bivalves harboring large amounts of HAB-toxins.
During HABs, toxic phytoplankton cells are easily ingested in large amounts by filter-feeding organisms such as bivalve mollusks, rendering them unsafe for consumption. For this reason, most coastal countries have a monitoring program in place to minimize the risk of acute intoxication by banning contaminated bivalves from reaching the markets. But in addition to their well-defined role as vectors of HAB toxins to humans, bivalve mollusks may also act as vectors within the marine food web and potentiate the transfer of HAB toxins to their marine predators. With the number and size of mussel production platforms increasing throughout several coastal areas, and considering that mussels as shellfish species are particularly prone to accumulate marine biotoxins such as okadaic acid [10,11,12], it is relevant to study the impact of implementing these structures on the environment in terms of availability of toxins to the surrounding food web. Longline mussel farms are complex, three-dimensional floating structures that commonly occupy a considerable area of sheltered environments. A shift on the pelagic fish diet around mussel farms has been observed, with mussels representing the dominant food item of these fish [13].
Okadaic acid (OA) and related toxins are commonly abundant and frequent groups of marine toxins in southern Europe, Asia, and South America [14]. These toxins accumulate to a great extent in bivalve mollusks, recurrently reaching high concentrations in tissues of certain species like mussels that can remain contaminated for long periods of time [11,15]. Although regularly available in the marine environment, little is known about either the accumulation/elimination dynamics of diarrhetic shellfish poisoning (DSP) toxins in fish or their effects, which contrasts with the number of research studies carried out regarding paralytic shellfish poisoning (PSP)and amnesic shellfish poisoning (ASP) toxins [7,9].
The aim of this study is to raise attention concerning the exposure of fish to the abundant and commonly available DSP toxins by gathering and summarizing the data dispersed in the literature about bioaccumulation and effects of OA-group toxins in fish and discussing their ecosystem-wide impacts.
2. Okadaic Acid-Group Toxins
Okadaic acid (OA) and its related compounds dinophysistoxins (DTXs) constitute a complex structural family of lipophilic polyether compounds. These toxins are responsible for the human foodborne diarrhetic shellfish poisoning (DSP), which is characterized by gastrointestinal disorders with acute symptoms including diarrhea, abdominal cramps, nausea, and vomiting [16].
Although first discovered and isolated from the black sponges Halichondria okadai and Halichondria melanodocia in 1981 [17], OA was later found to be globally distributed and produced as a secondary metabolite by marine dinoflagellates of the genera Dinophysis, Prorocentrum, and Phalacroma [18,19,20,21]. In addition to OA, other related compounds named dinophysistoxins (DTXs) that include isomers (DTX-2), analogues (DTX-1), and diol-ester derivatives (e.g., DTX-4 and 5) are produced by Dinophysis and Prorocentrum species. Also, a number of fatty acid ester derivatives of OA and DTX-1&2 resulting from shellfish metabolism are known and designated as DTX-3 [22,23,24] (Figure 1).
OA and DTX-1&2 are cell-permeable compounds that induce intestine damage, which is primarily caused by inhibition of the serine/threonine protein phosphatases (PP), PP1 and PP2A [25], leading to alterations in several intracellular processes and causing severe mucosal damages in the intestinal tract [26]. These phosphatases are also involved in other key cellular metabolic processes, including growth, division, death, and maintenance of cytoskeletal structure. In addition to its role as a strong inducer of diarrhetic activity, resulting from inhibition of proteins phosphatase (PP1 and PP2A), OA is known to exert cytotoxic, neurotoxic, immunotoxic, carcinogenic, and genotoxic effects [27]. Therefore, OA has been considered a key molecule with a high potential impact on the biology of aquatic animals, including effects on fitness and dependent functions of fishes [28,29].
3. OA-Group Toxins in Fish
3.1. Field Observations
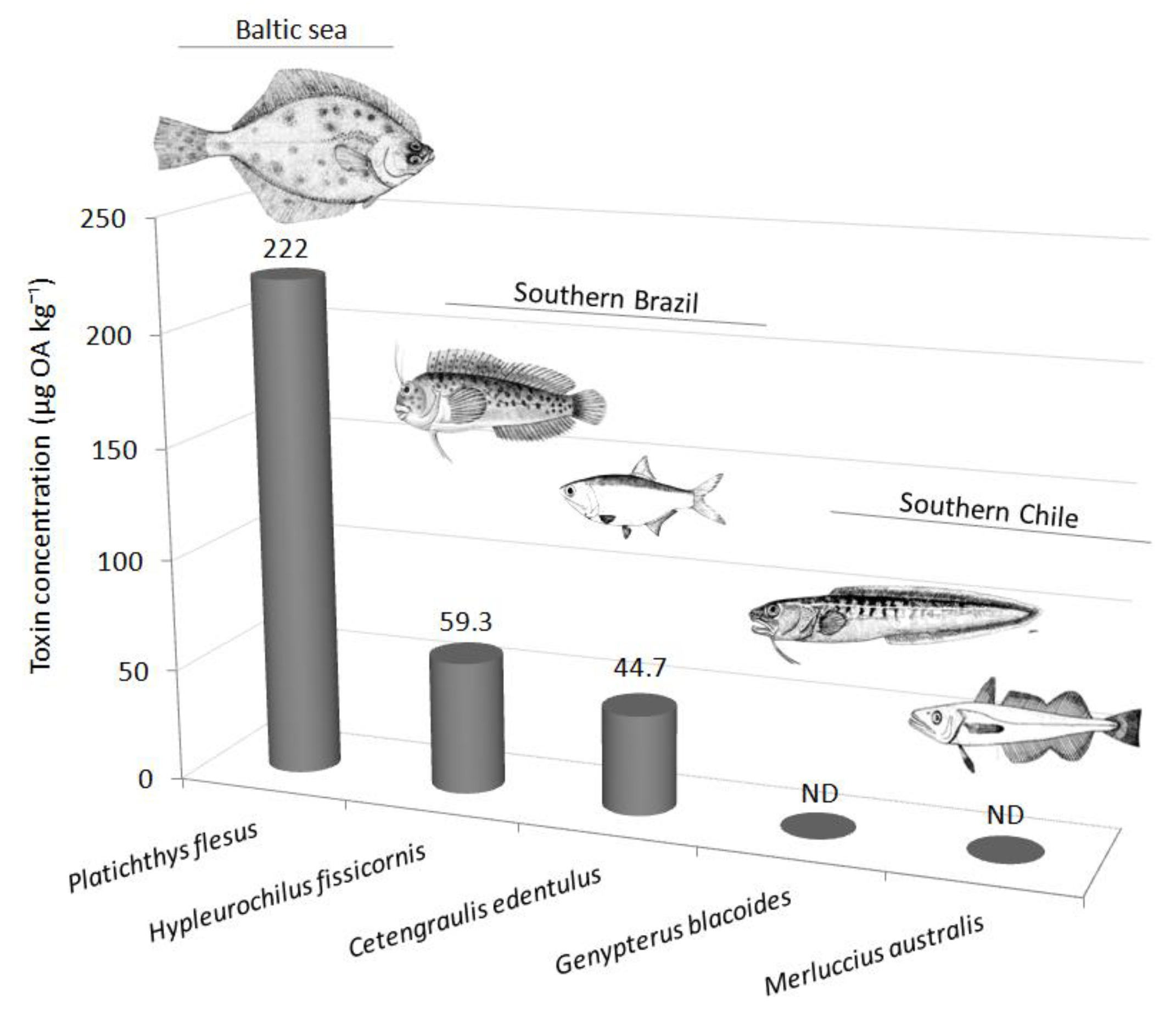
The first observation of OA in marine fishes dates back to 1996 in the northern Baltic Sea [30]. OA was found in the liver of flounders (Platichthys flesus) collected in the Gulf of Finland after a Dinophysis acuminata bloom (520–1960 cells L−1). Caught at depths between 6 and 20 m, flounders revealed toxin concentrations in the liver up to 222 ± 10 µg OA kg−1 [30]. Subsequent studies proved that OA could be detected as well in filter-feeder fish [30] and grazers [2].
A pooled sample of liver and digestive tract tissues from the filter-feeder Cetengraulis edentulus (the Atlantic anchoveta) sampled in Paranaguá Bay during a D. acuminata bloom contained 44.7 µg OA kg−1. Samples of Mugil sp. collected in the same area did not present detectable levels of OA, and DTX-1 was below the limit of detection in both species. The reported toxin levels were moderate considering the concentration of D. acuminata in water, which reached 19,500 cells L−1 at the surface. While microalgae abundance was greater at the surface, their cellular toxin content was significantly higher in D. acuminata sampled close to the bottom [31]. This can explain the occurrence of important levels of OA toxin in benthic fish, even at comparatively lower dinoflagellate water concentrations [30].
The distribution of free and esterified OA (i.e., DTX-3) in different trophic levels and along bloom development was studied in the southern coast of Brazil [2]. In the combtooth blenny fish Hypleurochilus fissicornis, mostly free OA was detected. Fish were contaminated for shorter periods than bivalves, crustaceans, and polychaetes and attained their maximum during the mid-bloom period, accumulating 59.3 µg OA kg−1 with 54 ± 7% in its free form [2]. Paradoxically, amphipods, some of their main prey, accumulated relatively high OA levels only a few days later, at the peak of the bloom [2]. Another survey in Aysén Region, Southern Chile, examined several endemic marine organisms during Dinophysis blooms without, however, detecting OA toxins in fish [32]
These studies demonstrate that OA may enter the food webs through vectors other than filter-feeding organisms. The few studies dealing with the accumulation of toxins in the environment suggest that fish accumulate mainly free OA in viscera for short periods of time. However, data of OA and related compounds in fish are still limited to a very reduced number of species (Figure 2).
3.2. Investigation of OA Effects in Marine Fish under Laboratory Controlled Conditions
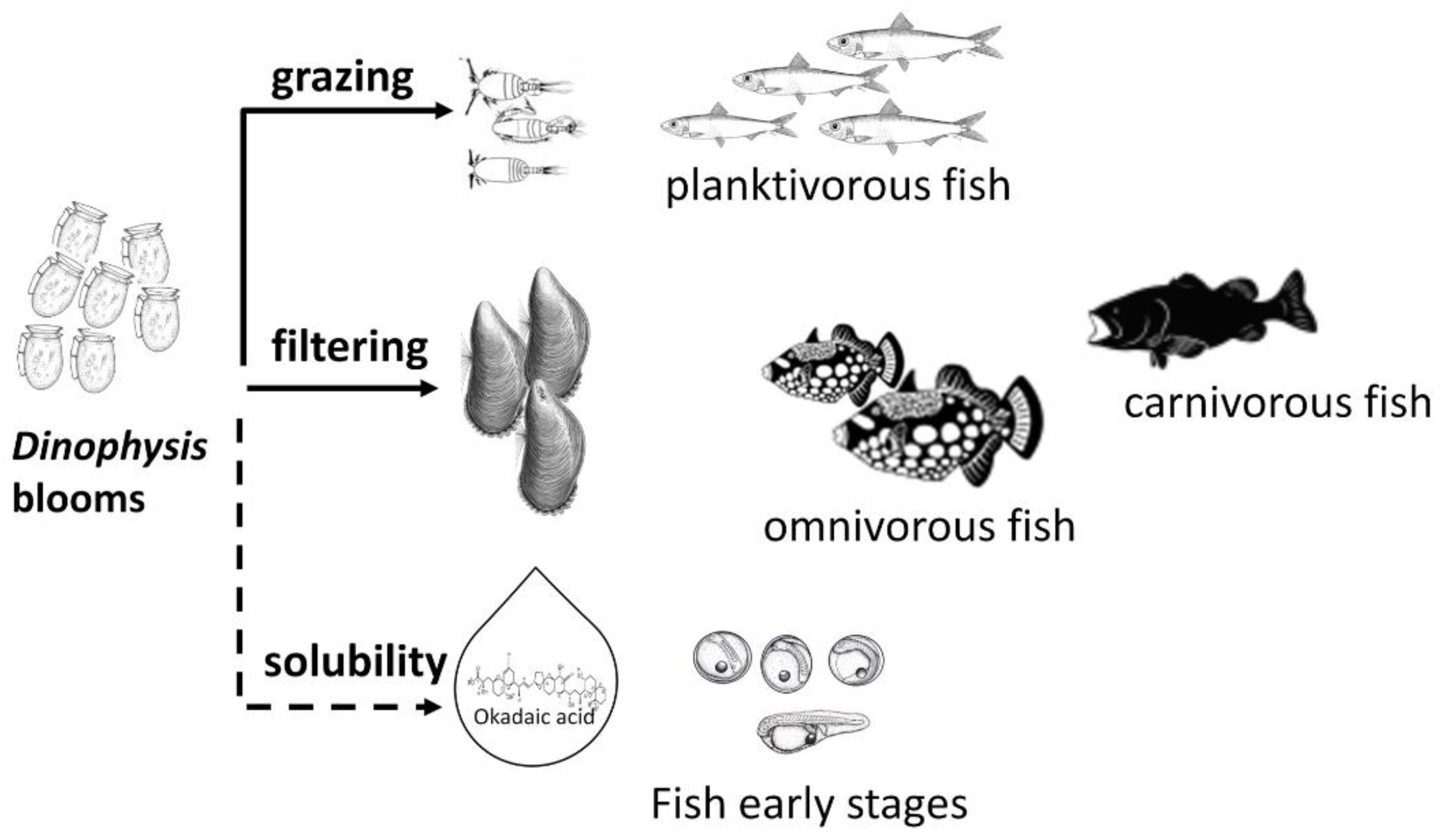
Although limited data are available regarding field observations, there is evidence of okadaic acid impacting marine fish at different life stages from embryo to adults, and either exposed via dietary route or via toxins dissolved in seawater. Figure 3 aims to illustrate the main routes of fish exposure to these toxins.
Fish may be exposed to marine toxins, such as okadaic acid, in their early life stages. Fish embryos may be particularly sensitive to marine toxins as they have not developed yet any effective enzymatic system to eliminate the toxins and may be adversely affected due to higher metabolic growth rates [33]. Therefore, a drastic impact on hatching success was observed in fertilized eggs from natural spawning of longfin yellowtail (Seriola rivoliana) exposed to two concentrations of dissolved OA and DTX-1 [29]. Reduction of 62% in hatching success was observed in fish eggs exposed to the lowest concentration (120 µg OA eq. L−1) and no survival was registered with the highest concentration tested (175 µg OA eq. L−1). Le Du and colleagues [29] were also able to show that OA and DTX-1 significantly inhibit protein and alkaline phosphatases, affecting the regulation pathways associated with embryogenesis and altering gene expression.
The impact of OA in the early life stages of other fish species was investigated in newly hatched eleutheroembryos and one week-old larvae of two estuarine species (Menidia beryllina and Cyprinodon variegatus). Exposure of these fish to toxin-producing dinoflagellates Alexandrium catenella (producer of saxitoxins) and Dinophysis acuminata (producer of OA) promoted lethal and sublethal (i.e., growth, grazing rate, and swimming activity) effects [34]. An additive effect in terms of fish survival was observed in specimens exposed to a mixture of A. catenella and D. acuminata though the effect was largely derived from A. catenella.
Juveniles and adult fish are also known to be adversely affected by OA and DTXs. Oxidative stress, histological alterations in several organs, behavior changes, and, in extreme cases, death of fish have been reported [28,35,36]. Fish mortality was registered in juvenile seabass (Dicentrarchus labrax) after three weeks feeding on brine shrimp (Artemia that has fed on P. lima cells) and fish that were fed a commercial diet containing live P. lima cells [28]. Moreover, abnormal behavior characterized by jumps, uncoordinated swims, fast left-right turns, surface swims, poor feeding coordination, and inactivity were observed when exposed to the culture medium of P. lima dinoflagellates [28]. However, OA and DTX-1 concentrations were not determined and the provided nominal doses were not assessed. Similar studies using brine shrimp as the trophic link species for phycotoxins transfer were recently carried out by Neves and colleagues [36], assessing sublethal effects of short-term exposure of the ringneck blenny (Parablennius pilicornis). Increased opercular activity, which can be considered an indicator of aerobic metabolism in fish, was observed and suggested as a mechanism of detoxification [36]. To investigate whether OA affects respiratory metabolism in fish and their ability to meet oxygen demands, Corriere and colleagues [37] fed zebra seabream (Diplodus cervinus) during three days with an artificial diet containing 1300 μg OA eq. kg−1 and challenged the fish with increasing flow velocities in a swim tunnel. While no statistical differences in demands of oxygen consumption were found, a reduction in swimming performance was observed for fish exposed to OA under increased stimulus. Corriere and colleagues [37] suggest that OA may have a cryptic effect that is not easily detected under favorable environmental conditions, but it is disclosed once fish have to deal with multiple environmental stressors, leading to lower metabolic fitness and reduction of swimming performances.
OA effects at the histological level were highlighted in the early studies carried out by Ajuzie [28], indicating the gills and the liver as the organs most affected by OA and DTXs. Recently, further investigations were performed to better understand the impact of these toxins, their histopathological damages, and induction of oxidative stress. Exposure of gilthead seabream (Sparus aurata) to dissolved high OA doses (7.5 µg mL−1) showed a time-dependent induction of oxidative stress [35]. Analysis of individual antioxidant enzymes such as catalase activity (CAT), glutathione level (GSH), and the assessment of the accumulation of lipid peroxidation product malondialdehyde (MDA) in liver tissue revealed the highest activity of CAT and GSH after 24 h exposure. Severe lipid peroxidation was also observed after 24 h with increased levels of MDA. Measurement of MDA as an end product of lipid peroxidation is widely used as a simple and reliable approach to evaluate the ROS-induced lipid peroxidation [35].
From histological observations the most pronounced changes were observed in the liver after 24 h of OA exposure, corresponding to capillary hyperemia, vascular dilation, alteration in the size of the hepatocyte, and visible hepatocellular membrane disintegration. The gills’ histopathology showed many histological changes reaching the maximum of lesions frequency after 24 h of exposure to the toxin, where hypertrophy was observed in the secondary lamellae with the enlargement of the tissues and a fusion of the primary lamella, dealing with a necrotic alteration [35].
It is important to recognize that a limited number of studies have been performed. Table 1 summarizes the studies carried out to investigate the effects of OA and DTXs in fish. Some of them did not provide data on the dose of exposure and others were not able to justify the exposure with realistic, ecologically relevant levels. Further research is critically needed to comprehensively understand the mechanisms of OA toxicity in fish, their biotransformation and elimination. Nevertheless, more information can be obtained from (1) studies that used freshwater fish species, such as zebra and medaka fish, as organism models to assess the exposure of OA, and (2) exposure to the cyanobacterial toxin microcystin-LR, also an inhibitor of the protein phosphatases PP1 and PP2A.
3.3. Investigation of OA Effects Using Zebra and Medaka as Fish Models
Fish models, such as medaka (Oryzias latipes) and zebra fish (Danio rerio) are increasingly used in toxicological and biomedical studies for their well-known physiology and easy manipulation [38,39]. Despite the fact that these species would rarely be exposed to OA in the environment, they provide useful data to determine potential damages in other organisms [40].
Medaka fish embryos exposed to OA and Prorocentrum arenarium crude extracts until hatching displayed reduced survival rate in a dose-dependent manner, reaching 100% mortality at concentrations 0.75 μg mL−1 for pure OA (EC50 0.52 μg mL−1) and 0.22 μg OA eq mL−1 for the toxic extract (EC50 0.17 μg OA eq mL−1). In surviving embryos a delay in development was also observed. Histological studies revealed abnormal development of organs that differed from fish treated with pure OA and P. arenarium extracts. Pure toxins affected mainly liver and digestive tracts while an increase in global body and vitellus areas was detected in extract-treated embryos [41].
The early life stages of zebra fish are also sensitive to OA and DTX-1. Larvae exposed to sublethal concentrations of OA or DTX-1 (EC50 8 μg OA mL−1 and 6 μg DTX-1 mL−1, 24 h) displayed a developmental delay and evident macroscopic alterations (pericardial edema, cyclopia, and reduced anteroposterior axis). Oxidative damage and altered levels of the biochemical biomarkers superoxide dismutase (SOD) and catalase (CAT), glutathione peroxidase (GPx), and glutathione reductase (GR) were manifested in a dose-dependent way at the different larval stages [42]. DTX-1 produced effects at lower concentrations than OA, or in shorter periods of time. For instance, morphological abnormalities were not specific to a certain toxin but DTX-1 produced more abnormalities. The same trend was observed when comparing OA or DTX-1 inhibitory effects on the SOD and CAT complex and increases in GPx and GR activity [42].
In zebra fish adult stages, OA has been shown to produce changes in liver gene expression after intraperitoneal injection. These regulated genes were involved in several essential functions such as response to xenobiotics, protein phosphatase activity, heat-shock protein binding, and transmembrane transportation, among others. An increase of OA concentration by 10 caused a proportional number of up and low regulated genes. Consequently, the expression of 91 genes was altered in low-dosed fish (0.176 μg OA g−1 bw) while 985 appeared up and low-regulated in high-dosed individuals (1.76 μg OA g−1 bw). This group presented evident signs of toxicity as well, namely change of color, impaired movements, and death only 30 min after injection [43].
Recent studies devoted to Alzheimer’s disease have used the capability of OA to induce Alzheimer-like pathology in adult zebra fish. Fish exposed to OA exhibited learning and memory deficits. Deposition of a β-fragment, hyperphosphorylation of tau protein, and formation of senile plaques were also observed, likewise in Alzheimer’s disease [44,45,46]. Although these studies were not primarily aimed at the neurotoxicity evaluation of OA in fish, they emphasize the ecological impact that OA can have on fish in their natural environment. Learning and spatial cognitive behavior may be crucial for fish survival.
3.4. Research Clues from the Freshwater Environment
Okadaic acid (OA) and microcystins (MCs), which are the most common cyanobacterial toxins found in the water systems, from fresh to brackish waters, are structurally unrelated compounds produced by phylogenetically unrelated species which are known to inhibit a common target, the protein ser/thr phosphatases. MCs are produced by numerous genera of cyanobacteria with emphasis on Microcystis, the most common bloom-forming and main producer genus of MCs in freshwater ecosystems [47].
The MCs inhibition of protein phosphatases is remarkably similar to OA [48]. It was demonstrated that the carboxylic group of OA folds on itself to form a pseudo-ring structure with a hydrophobic tail similar to that of the most common Microcystin Luecine Arginine (MC-LR).
MCs received considerable attention due to their widespread occurrence in fresh waters and deleterious effects on humans and aquatic biota. Bivalve mollusks, as well as in the case of OA and other marine toxins, are among the most threatened group of aquatic organisms because of their filtration feeding behavior [49]. Fish can come in contact with MCs in two main ways: oral uptake of toxin-containing cells or via epithelium surfaces by immersion in water containing dissolved toxins. Effects of MCs in fish were extensively explored and summarized by [49,50] in their reviews. Both studies reported histopathological effects in the fish liver, intestine, kidney, and gills, although the degree to which these effects were measured varied with the exposure route.. And other effects, namely cardiac, hematological, biochemical, and immunological alterations, ionic imbalance, and reduced growth were also detected in several fish species, indicating that MCs promote oxidative stress, inducing the production of ROS and lipid peroxidation. From the range of effects associated with MCs exposure, it is important to focus on those related to protein phosphatase inhibition. In fish hepatocytes, MCs inhibit PP1 and PP2A and this inhibition can disturb the cellular phosphorylation balance, causing a marked increase of ROS leading towards oxidant shock. If antioxidant enzymes cannot succeed in eliminating ROS, the oxidant stress leads to apoptosis and even necrosis of cells [49,50]. As well as for OA, early life stages seem more susceptible to MCs’ toxicity than adults or juveniles, especially during the critical phases of the development of embryos and larvae [33].
Although common in freshwater environments, and in brackish waters too, the discharge of intense Microcystis blooms into the coastal ocean has been leading to microcystins’ contamination of the marine ecosystems [51]. The most significant and striking case was derived from MCs’ accumulation in top marine predators at extremely high concentrations, which resulted in the deaths of 21 sea otters (Enhydra lutris nereis) [52]. Until now two studies have described MCs’ accumulation and effects in marine/estuarine fish [53,54]. During high density Microcystis aeruginosa blooms (32 × 10⁹ cells L−1) in San Francisco Estuary, USA, histopathology observations revealed high damage in liver tissue of fish (striped bass, Morone saxatilis, and Mississippi silversides, Menidia audens) [53].
4. Final Considerations
To understand the impact of OA and DSP toxins in fish it is also necessary to analyze the ecosystem responses to these toxins. The toxigenic dinoflagellate genus Dinophysis and Prorocentrum spp. contribute to blooms maintenance. Alterations in nutritional and environmental factors, such as nitrogen or phosphorus limitation and low salinity, may increase levels of DSP toxins, especially in Prorocentrum species. Dinophysis populations depend on favorable abiotic conditions but are also affected by mixotrophy, considered a key process for their development [2,18]. The toxins produced by blooms of these dinoflagellates can be transferred along the food web leading to cascading effects on community structures in marine systems. However, data on bioaccumulation of DSP toxins in organisms other than bivalve mollusks, such as fish, are particularly limited, even if OA and DTXs are among the most common and abundant HAB-toxins worldwide. As reviewed in the present study, exposure of fish to OA-group toxins is foreseen to increase, either due to anthropogenic actions such as the increasing number of mussels farming nets and growing ropes in many of the coastal areas [13], or as a result of climate change conditions that may favor Dinophysis blooms formation [1] and toxin production [55], or even a combination of the two factors [56]. Despite the reduced data available it is clear that exposure of fish to OA-group toxins produces a range of unreported negative effects at different life stages, either by waterborne or dietary routes, that goes from impacts in fish fitness, swimming performance, feeding, foraging, and escaping from predators to ultimately reduced fish abundance. The overall ecological risk associated with contamination of fish with OA-group toxins may increase when blooms of different marine harmful species co-occur, as described by Rountos et al. [34], or overlap with persistent cyanobacterial blooms discharged from estuaries into nearshore coastal areas.
Author Contributions
Conceptualization, P.R.C. and L.S.; investigation, M.C.; resources, P.R.C.; writing—original draft preparation, M.C.; writing—review and editing, P.R.C. and L.S.; project administration, P.R.C.; funding acquisition, P.R.C. All authors have read and agreed to the published version of the manuscript.
Funding
The research leading to these results has received funding from the project Cigua (PTDC/CTA-AMB/30557/2017) supported by the Portuguese Foundation for Science and Technology (FCT) and FEDER. This study also received Portuguese national funds from FCT through project UID/Multi/04326/2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, T.P.; Mafra, L.L. Diel variations in cell abundance and trophic transfer of diarrheic toxins during massive Dinophysis bloom in southern Brazil. Toxins 2018, 10, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grattan, L.M.; Holobaugh, S.; Morris, J.G. Harmful algal blooms and public health. Harmful Algae 2016, 57, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallegraeff, G.M. Ocean climate change, phytoplankton community responses, and harmful algal blooms: A formidable predictive challenge. J. Phycol. 2010, 46, 220–235. [Google Scholar] [CrossRef]
- Reizopoulou, S.; Strogyloudi, E.; Giannakourou, A.; Pagou, K.; Hatzianestis, I.; Pyrgaki, C.; Granéli, E. Okadaic acid accumulation in macrofilter feeders subjected to natural blooms of Dinophysis acuminata. Harmful Algae 2008, 7, 228–234. [Google Scholar] [CrossRef]
- Pavagadhi, S.; Balasubramania, R. Toxicological evaluation of microcystins in aquatic fish species: Current knowledge and future directions. Aquat. Toxicol. 2013, 142–143, 1–16. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Frame, E.R.; Kendrick, P.S. Domoic acid and fish behavior: A review. Harmful Algae 2012, 13, 126–130. [Google Scholar] [CrossRef]
- Costa, P.R.; Garrido, S. Domoic acid accumulation in the sardine Sardina pilchardus and its relationship to Pseudonitzschia diatom ingestion. Mar. Ecol. Prog. Ser. 2004, 284, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.R. Impact and effects of paralytic shellfish poisoning toxins derived from harmful algal blooms to marine fish. Fish Fish. 2016, 17, 226–248. [Google Scholar] [CrossRef]
- Vale, P.; Sampayo, M.A.D.M. Esterification of DSP toxins by Portuguese bivalves from the Northwest coast determined by LC-MS—A widespread phenomenon. Toxicon 2002, 40, 33–42. [Google Scholar] [CrossRef]
- Vale, P.; Botelho, M.J.; Rodrigues, S.M.; Gomes, S.S.; Sampayo, M.A.D.M. Two decades of marine biotoxin monitoring in bivalves from Portugal (1986–2006): A review of exposure assessment. Harmful Algae 2008, 7, 11–25. [Google Scholar] [CrossRef]
- McCarthy, M.; O’Halloran, J.; O’Brien, N.M.; Van Pelt, F.F.N.A.M. Does the marine biotoxin okadaic acid cause DNA fragmentation in the blue mussel and the pacific oyster? Mar. Environ. Res. 2014, 101, 153–160. [Google Scholar] [CrossRef]
- Šegvić-Bubić, T.; Grubišić, L.; Karaman, N.; Tičina, V.; Jelavić, K.M.; Katavic, I. Damages on mussel farms potentially caused by fish predation—Self service on the ropes? Aquaculture 2011, 319, 497–504. [Google Scholar] [CrossRef]
- EFSA. Opinion of the Scientific Panel on Contaminants in the Food Chain on a Request from the European Commission on Marine Biotoxins in Shellfish—Okadaic Acid and Analogues. EFSA J. 2008, 589, 1–62. [Google Scholar]
- Rossignoli, A.E.; Fernandez, D.; Regueiro, J.; Marino, C.; Blanco, J. Esterification of okadaic acid in the mussel Mytilus gallo-provincialis. Toxicon 2011, 57, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Hossen, V.; Jourdan-da Silva, N.; Guillois-Bécel, Y.; Marchal, J.; Krys, S. Food poisoning outbreaks linked to mussels contaminated with okadaic acid and ester dinophysistoxin-3 in France, June 2009. Eurosurveillance 2011, 16. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, K.; Scheuer, P.J.; Tsukitani, Y.; Kikuchi, H.; Van Engen, D.; Clardy, J.; Gopichand, Y.; Schmitz, F.J. Okadaic acid, a cytotoxic polyether from two marine sponges of the genus Halichondria. J. Am. Chem. Soc. 1981, 103, 2469–2471. [Google Scholar] [CrossRef]
- Lee, T.C.-H.; Fong, L.L.-Y.; Ho, K.-C.; Lee, F.W.-F. The mechanism of diarrhetic shellfish poisoning toxin production in Proro-centrum spp.: Physiological Molecular Perspectives. Toxins 2016, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Identification of Dinophysis fortii as the causative organism of diarrhetic shellfish poisoning. Bull. Jpn. Soc. Sci. Fish. 1980, 46, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycol. 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Reguera, B.; Riobó, P.; Rodríguez, F.; Díaz, P.A.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis Toxins: Causative Organisms, Distribution and Fate in Shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef]
- Dominguez, H.J.; Paz, B.; Daranas, A.H.; Norte, M.; Franco, J.M.; Fernández, J.J. Dinoflagellate polyether within the yessotoxin, pectenotoxin and okadaic acid toxin groups: Characterization, analysis and human health implications. Toxicon 2010, 56, 191–217. [Google Scholar] [CrossRef]
- Rossignoli, A.E.; Blanco, J. Subcellular distribution of okadaic acid in the digestive gland of Mytilus galloprovincialis: First evidences of lipoprotein binding to okadaic acid. Toxicon 2010, 55, 221–226. [Google Scholar] [CrossRef]
- Hu, T.; Curtis, J.M.; Walter, J.A.; McLachlan, J.L.; Wright, L.C. Two new water soluble DSP toxin derivatives from dinoflagellate Prorocentrum maculosum: Possible storage and excretion products. Tetrahedron Lett. 1995, 36, 9273–9276. [Google Scholar] [CrossRef]
- Ikema, S.; Takumi, S.; Maeda, Y.; Kurimoto, T.; Bohda, S.; Chigwechokha, P.K.; Sugiyama, Y.; Shiozaki, K.; Furukawa, T.; Komatsu, M. 2015 Okadaic acid is taken-up into the cells hepatocytes transporter OATP1B3. Food Chem. Toxicol. 2015, 83, 229–236. [Google Scholar] [CrossRef]
- Matias, W.G.; Traore, A.; Creppy, E.E. Variations in the distribution of okadaic acid in organs and biological fluids of mice related to diarrhoeic syndrome. Hum. Exp. Toxicol. 1999, 18, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Pásaro, E.; Méndez, J.; Laffon, B. Okadaic Acid: More than a Diarrheic Toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajuzie, C.C. Toxic Prorocentrum lima induces abnormal behavior in juvenile sea bass. J. Appl. Phycol. 2007, 20, 19–27. [Google Scholar] [CrossRef]
- Le Du, J.; Tovar-Ramírez, D.; Núñez-Vázquez, E. Embryotoxic effects of dissolved okadaic acid on the development of Longfin yellowtail Seriola rivoliana. Aquat. Toxicol. 2017, 190, 210–216. [Google Scholar] [CrossRef]
- Sipiä, V.; Kankaanpää, H.; Meriluoto, J.; Høisæter, T. The first observation of okadaic acid in flounder in the Baltic Sea. Sarsia 2000, 85, 471–475. [Google Scholar] [CrossRef]
- Mafra, L.L.; Dos Santos Tavares, C.P.; Schramm, M.A. Diarrheic toxins in field-sampled and cultivated Dinophysis spp. cells from southern Brazil. J. Appl. Phycol. 2014, 26, 1727–1739. [Google Scholar] [CrossRef]
- Contreras, H.R.; García, C. Inter-species variability of okadaic acid group toxicity in relation to the content of fatty acids detected in different marine vectors. Food Addit. Contam. Part A 2019, 36, 464–482. [Google Scholar] [CrossRef]
- Vasconcelos, V.; Azevedo, J.; Silva, M.; Ramos, V. Effects of Marine Toxins on the Reproduction and Early Stages Development of Aquatic Organisms. Mar. Drugs 2010, 8, 59–79. [Google Scholar] [CrossRef] [Green Version]
- Rountos, K.J.; Kim, J.J.; Hattenrath-Lehmann, T.K.; Gobler, C.J. Effects of the harmful algae, Alexandrium catenella and Dinophysis acuminata, on the survival, growth, and swimming activity of early life stages of forage fish. Mar. Environ. Res. 2019, 148, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Souid, G.; Souayed, N.; Haouas, Z.; Maaroufi, K. Does the phycotoxin okadaic acid cause oxidative stress damages and his-tological alterations to seabream (Sparus aurata)? Toxicon 2018, 144, 55–60. [Google Scholar] [CrossRef]
- Neves, R.A.F.; Nascimento, S.M.; Santos, L.N. Sublethal fish responses to short-term food chain transfer of DSP toxins: The role of somatic condition. J. Exp. Mar. Biol. Ecol. 2020, 524, 151317. [Google Scholar] [CrossRef]
- Corriere, M.; Baptista, M.; Paula, J.R.; Repolho, T.; Rosa, R.; Costa, P.R.; Soliño, L. Impaired fish swimming performance following dietary exposure to the marine phycotoxin okadaic acid. Toxicon 2020, 179, 53–59. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Chiang, C.-Y.; Tsai, H.-J. Zebrafish and Medaka: New model organisms for modern biomedical research. J. Biomed. Sci. 2016, 23, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Schubert, S.; Keddig, N.; Hanel, R.; Kammann, U. Microinjection into zebrafish embryos (Danio rerio)—A useful tool in aquatic toxicity testing? Environ. Sci. Eur. 2014, 26, 1. [Google Scholar] [CrossRef] [Green Version]
- Berry, J.P.; Gantar, M.; Gibbs, P.D.; Schmale, M.C. The zebrafish (Danio rerio) embryo as a model system for identification and characterization of developmental toxins from marine and freshwater microalgae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escoffier, N.; Gaudin, J.; Mezhoud, K.; Huet, H.; Château-Joubert, S.; Turquet, J.; Crespeau, F.; Edery, M. Toxicity to medaka fish embryo development of okadaic acid and crude extracts of Prorocentrum dinoflagellates. Toxicon 2007, 49, 1182–1192. [Google Scholar] [CrossRef]
- Figueroa, D.; Signore, A.; Araneda, O.; Contreras, H.R.; Concha, M.; García, C. Toxicity and differential oxidative stress effects on zebrafish larvae following exposure to toxins from the okadaic acid group. J. Toxicol. Environ. Health Part A 2020, 83, 573–588. [Google Scholar] [CrossRef]
- Zhang, N.-S.; Li, H.-Y.; Liu, J.-S.; Yang, W.-D. Gene expression profiles in zebrafish (Danio rerio) liver after acute exposure to okadaic acid. Environ. Toxicol. Pharmacol. 2014, 37, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Nada, S.; Williams, F.; Shah, Z. Development of a novel and robust pharmacological model of okadaic acid-induced Alzheimer’s disease in zebrafish. CNS Neurol. Disord. Drug Targets 2010, 15, 86–94. [Google Scholar] [CrossRef]
- Koehler, D.; Shah, Z.A.; Hensley, K.; Williams, F.E. Lanthionine ketimine-5-ethyl ester provides neuroprotection in a zebrafish model of okadaic acid-induced Alzheimer’s disease. Neurochem. Int. 2018, 115, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Koehler, D.; Shah, Z.A.; Williams, F.E. The GSK3β inhibitor, TDZD-8, rescues cognition in a zebrafish model of okadaic ac-id-induced Alzheimer’s disease. Neurochem. Int. 2019, 122, 31–37. [Google Scholar] [CrossRef]
- Preece, E.P.; Moore, B.C.; Hardy, F.J. Transfer of microcystin from fresh water lakes to Puget Sound, WA and toxin accumulation in marine mussels (Mytilus trossulus). Ecotoxicol. Environ. Safe 2015, 122, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Mackintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Malbruck, C.; Kestemont, P. Effects of microcystins on fish. Environ. Toxicol. Chem. 2006, 25, 72–86. [Google Scholar] [CrossRef]
- Martins, J.C.; Vasconcelos, V.M. Microcystin Dynamics in Aquatic Organisms. J. Toxicol. Environ. Health Part B 2009, 12, 65–82. [Google Scholar] [CrossRef]
- Kruk, C.; Martínez, A.; Martínez de la Escalera, G.; Trinchin, R.; Manta, G.; Segura, A.M.; Piccini, C.; Brena, B.; Yannicelli, B.; Fabiano, G.; et al. Rapid freshwater discharge on the coastal ocean as a mean of long distance spreading of an unprecedented toxic cyanobacteria bloom. Sci. Total Environ. 2021, 754, 142362. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, W.A.; Miller, M.; Toy-Choutka, W.A.; Dominik, S.; et al. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lehman, P.W.; Teh, S.J.; Boyer, G.L.; Nobriga, M.L.; Bass, E.; Hogle, C. Initial impacts of Microcystis aeruginosa blooms on the aquatic food web in the San Francisco Estuary. Hydrobiology 2010, 637, 229–248. [Google Scholar] [CrossRef] [Green Version]
- De Pace, R.; Vita, V.; Bucci, M.S.; Gallo, P.; Bruno, M. Microcystin contamination in sea mussel farms from the Italian southern Adriatic coast following cyanobacterial blooms in the artificial reservoir. J. Ecosyst. 2014, 11. [Google Scholar] [CrossRef] [Green Version]
- Kamiyama, T.; Nagai, S.; Suzuki, T.; Miyamura, K. Effect of temperature on production of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 by Dinophysis acuminata in culture experiments. Aquat. Microb. Ecol. 2010, 60, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Hattenrath-Lehmann, T.K.; Marcoval, M.A.; Mittlesdorf, H.; Goleski, J.A.; Wang, Z.; Haynes, B.; Morton, S.L.; Gobler, C.J. Nitrogenous Nutrients Promote the Growth and Toxicity of Dinophysis acuminata during Estuarine Bloom Events. PLoS ONE 2015, 10, e0124148. [Google Scholar] [CrossRef]
Figure 1.
Chemical structure of the main diarrhetic shellfish poisoning (DSP) toxins: Okadaic acid and its derivatives, adapted from [18].
Figure 1.
Chemical structure of the main diarrhetic shellfish poisoning (DSP) toxins: Okadaic acid and its derivatives, adapted from [18].

Figure 2.
Documented levels of okadaic acid (µg kg−1) determined in fish worldwide [2,30,31,32]. ND: not detected.

Figure 3.
Foodborne (solid line) and waterborne (dashed line) exposure of marine fish to okadaic acid and dinophysistoxins.
Figure 3.
Foodborne (solid line) and waterborne (dashed line) exposure of marine fish to okadaic acid and dinophysistoxins.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of okadaic acid and dinophysistoxins in marine/estuarine fish.
| Species | Exposure Route/Challenge | Toxin Effects in Fish | Reference |
|---|---|---|---|
| Early life stages: | |||
| Longfin yellowtail (Seriola rivoliana) | Dissolved okadaic acid (120–175 µg L−1) |
| [29] |
| Sheepshead minnow (Cyprinodon variegates) Silversides (Menidia beryllina) | Mixture of toxic dinoflagellates: Dinophysis acuminata and Alexandrium catenella |
| [34] |
| Juveniles: | |||
| Seabass (Dicentrarchus labrax) | Toxic dinoflagellates (P. lima) and/or cell culture medium |
| [28] |
| Seabass (Dicentrarchus labrax) | Artemia that fed on P. lima and/or artificial feed containing P. lima |
| [28] |
| Gilthead seabream (Sparus aurata) | Dissolved okadaic acid (7500 µg L−1) |
| [35] |
| Ringneck blenny (Parablennius pilicornis) | Artemia that fed on P. lima |
| [36] |
| Zebra seabream (Diplodus cervinus) | Artificial feed containing 1300 μg OA eq. kg−1 followed by increased stimulus of swim current velocity |
| [37] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Corriere, M.; Soliño, L.; Costa, P.R. Effects of the Marine Biotoxins Okadaic Acid and Dinophysistoxins on Fish. J. Mar. Sci. Eng. 2021, 9, 293. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9030293
AMA Style
Corriere M, Soliño L, Costa PR. Effects of the Marine Biotoxins Okadaic Acid and Dinophysistoxins on Fish. Journal of Marine Science and Engineering. 2021; 9(3):293. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9030293
Chicago/Turabian StyleCorriere, Mauro, Lucía Soliño, and Pedro Reis Costa. 2021. "Effects of the Marine Biotoxins Okadaic Acid and Dinophysistoxins on Fish" Journal of Marine Science and Engineering 9, no. 3: 293. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9030293
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.