
Latitudinal Differentiation among Modern Planktonic Foraminiferal Populations of Central Mediterranean: Species–Specific Distribution Patterns and Size Variability

 , ,
, ,
Abstract
:1. Introduction
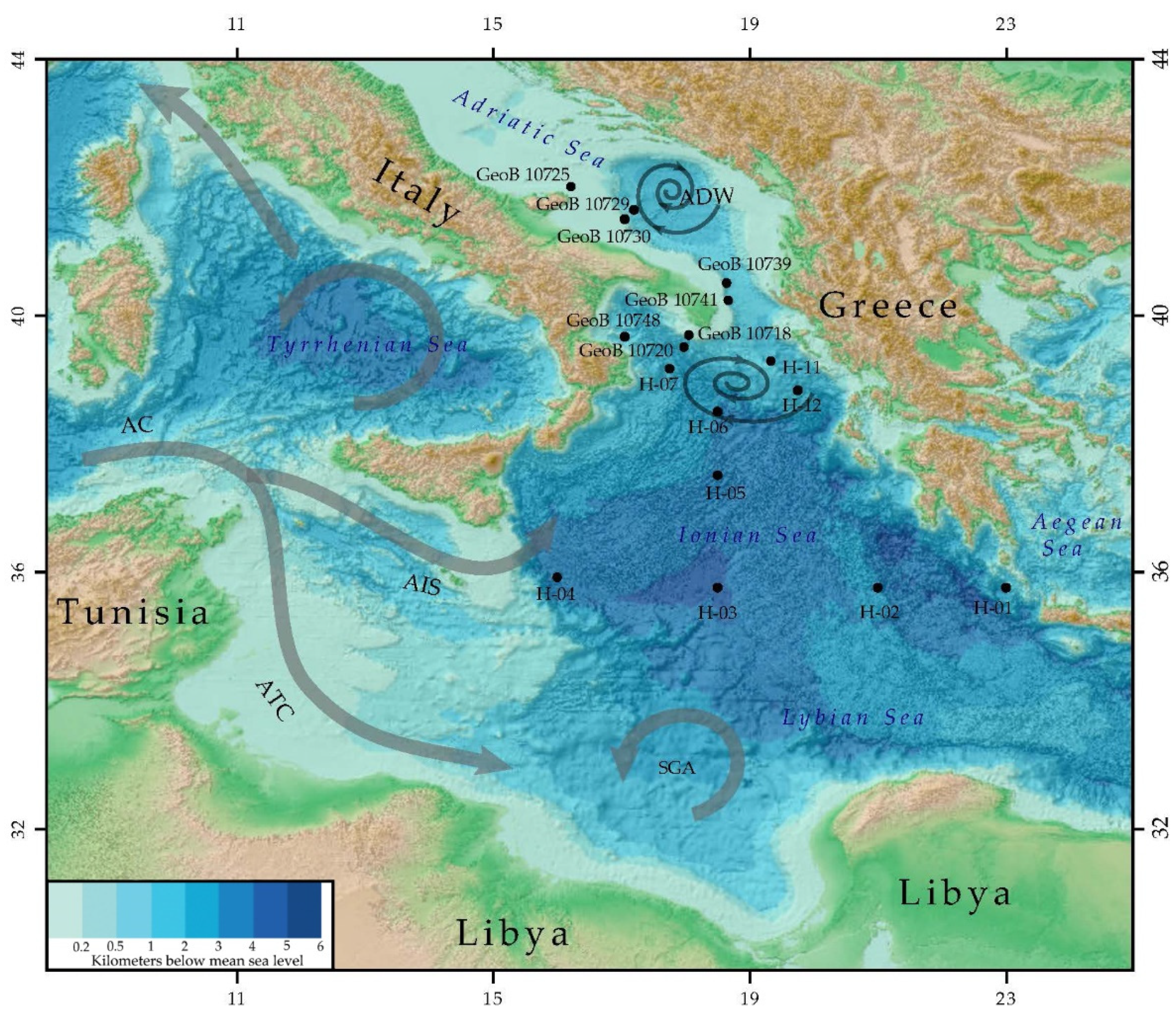
2. Study Area
2.1. Regional Oceanographic Setting
2.2. Water Masses and Circulation
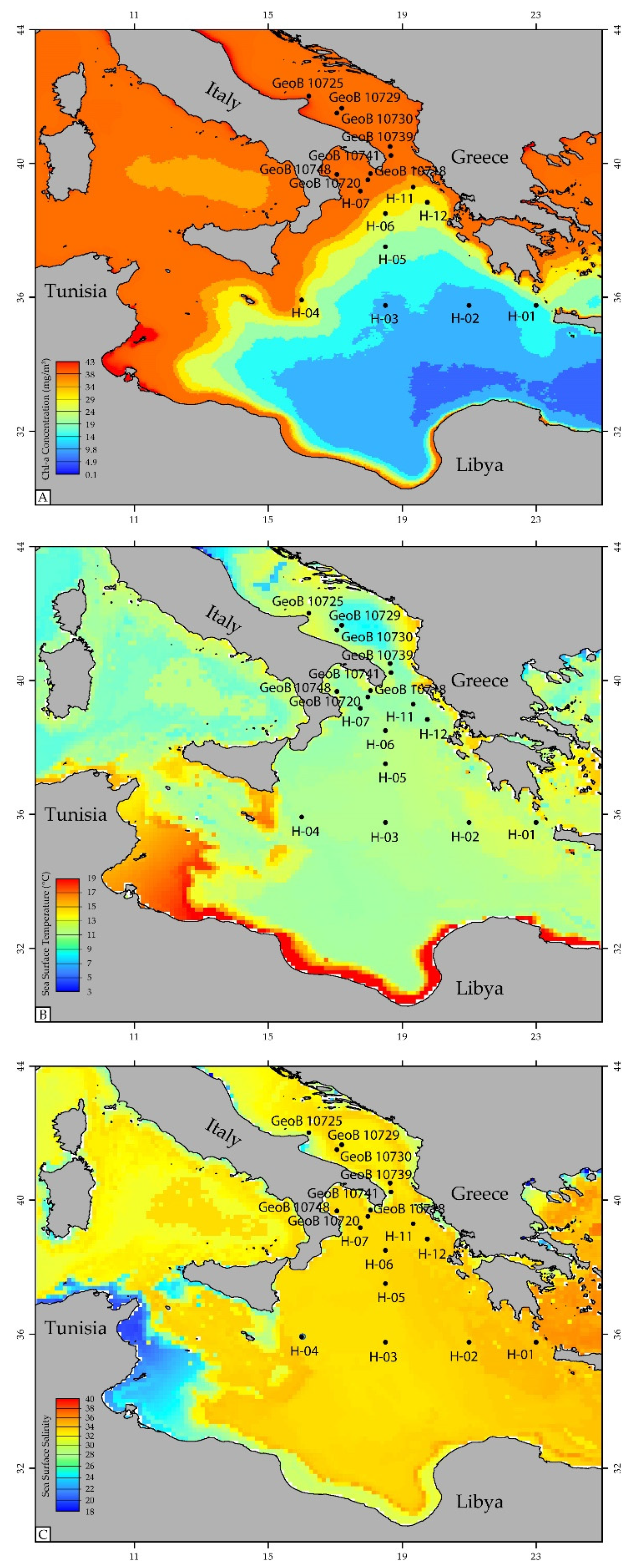
2.3. Productivity Regimes
3. Materials and Methods
3.1. Sediment Sampling Strategy and Site Selection
3.2. Micropaleontological Quantitative Analysis
3.3. Morphometric Analysis
4. Results
4.1. Relative Abundance Data
4.2. Preservation Regime and Size Variability
5. Discussion
5.1. Ecological and Oceanographic Context of the Planktonic Foraminiferal Biogeographic Distribution in Adriatic and Ionian Basins
5.2. On the Environmental Component on the Latitudinal Size Variability
5.3. The Possible Role of a Hidden Biological Diversity in Mediterranean Assemblages
5.4. Potential Depth Preferred Regulating Mechanism
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hemleben, C.; Anderson, O.R.; Spindler, M. Modern Planktonic Foraminifera; Springer: New York, NY, USA, 1989. [Google Scholar]
- Tsiolakis, E.; Tsaila-Monopoli, S.; Kontakiotis, G.; Antonarakou, A.; Sprovieri, M.; Geraga, M.; Ferentinos, G.; Zissimos, A. Integrated paleohydrology reconstruction and Pliocene climate variability in Cyprus Island (eastern Mediterranean). Iop Conf. Ser. Earth Environ. Sci. 2019, 362, 012103. [Google Scholar] [CrossRef]
- Antonarakou, A.; Kontakiotis, G.; Karageorgis, A.P.; Besiou, E.; Zarkogiannis, S.; Drinia, H.; Mortyn, G.P.; Tripsanas, E. Eco-biostratigraphic advances on late Quaternary geochronology and palaeoclimate: The marginal Gulf of Mexico analogue. Geol. Q. 2019, 63, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Budillon, F.; Lirer, F.; Iorio, M.; Macrì, P.; Sagnotti, L.; Vallefuoco, M.; Ferraro, L.; Garziglia, S.; Innangi, S.; Sahabi, M.; et al. Integrated stratigraphic reconstruction for the last 80 kyr in a deep sector of the Sardinia Channel (Western Mediterranean). Deep Sea Res. II 2009, 56, 725–737. [Google Scholar] [CrossRef] [Green Version]
- Drinia, H.; Antonarakou, A.; Tsourou, T.; Kontakiotis, G.; Psychogiou, M.; Anastasakis, G. Foraminifera eco-biostratigraphy of the southern Evoikos outer shelf, central Aegean Sea, during MIS 5 to present. Cont. Shelf Res. 2016, 126, 36–49. [Google Scholar] [CrossRef]
- Lirer, F.; Luca, M.; Iaccarino, S.; Gianfranco, S.; Turco, E.; Claudia, C.; Sierro, F.; Caruso, A. Mediterranean Neogene planktonic foraminifer biozonation and biochronology. Earth-Sci. Rev. 2019, 196. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Antonarakou, A.; Kouli, K.; Dimiza, M.; Kontakiotis, G.; Papanikolaou, M.D.; Ziveri, P.; Mortyn, P.G.; Lianou, V.; Lykousis, V.; et al. Late Glacial–Holocene ecostratigraphy of the south-eastern Aegean Sea, based on plankton and pollen assemblages. Geo-Mar. Lett. 2009, 29, 249–267. [Google Scholar] [CrossRef]
- Giamali, C.; Kontakiotis, G.; Koskeridou, E.; Ioakim, C.; Antonarakou, A. Key Environmental Factors Controlling Planktonic Foraminiferal and Pteropod Community’s Response to Late Quaternary Hydroclimate Changes in the South Aegean Sea (Eastern Mediterranean). J. Mar. Sci. Eng. 2020, 8, 709. [Google Scholar] [CrossRef]
- Giamali, C.; Koskeridou, E.; Antonarakou, A.; Kontakiotis, G.; Ioakim, C.; Karageorgis, A.; Roussakis, G.; Karakitsios, V. Multiproxy marine ecosystem response of abrupt Holocene climatic changes in the northeastern Mediterranean sedimentary archive. Quat. Res. 2019, 92, 665–685. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Besiou, E.; Antonarakou, A.; Zarkogiannis, S.D.; Kostis, A.; Mortyn, P.G.; Moissette, P.; Cornée, J.J.; Schulbert, C.; Drinia, H.; et al. Decoding sea surface and paleoclimate conditions in the eastern Mediterranean over the Tortonian-Messinian Transition. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 534, 109312. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Karakitsios, V.; Mortyn, P.G.; Antonarakou, A.; Drinia, H.; Anastasakis, G.; Agiadi, K.; Kafousia, N.; De Rafelis, M. New insights into the early Pliocene hydrographic dynamics and their relationship to the climatic evolution of the Mediterranean Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 459, 348–364. [Google Scholar] [CrossRef]
- Le Houedec, S.; Mojtahid, M.; Bicchi, E.; de Lange, G.J.; Hennekam, R. Suborbital Hydrological Variability Inferred From Coupled Benthic and Planktic Foraminiferal-Based Proxies in the Southeastern Mediterranean During the Last 19 ka. Paleoceanogr. Paleoclimatol. 2020, 35, e2019PA003827. [Google Scholar] [CrossRef]
- Louvari, M.A.; Drinia, H.; Kontakiotis, G.; Di Bella, L.; Antonarakou, A.; Anastasakis, G. Impact of latest-glacial to Holocene sea-level oscillations on central Aegean shelf ecosystems: A benthic foraminiferal palaeoenvironmental assessment of South Evoikos Gulf, Greece. J. Mar. Syst. 2019. [Google Scholar] [CrossRef]
- Margaritelli, G.; Cacho, I.; Català, A.; Barra, M.; Bellucci, L.G.; Lubritto, C.; Rettori, R.; Lirer, F. Persistent warm Mediterranean surface waters during the Roman period. Sci. Rep. 2020, 10, 10431. [Google Scholar] [CrossRef]
- Margaritelli, G.; Vallefuoco, M.; Di Rita, F.; Capotondi, L.; Bellucci, L.G.; Insinga, D.D.; Petrosino, P.; Bonomo, S.; Cacho, I.; Cascella, A.; et al. Marine response to climate changes during the last five millennia in the central Mediterranean Sea. Glob. Planet. Chang. 2016, 142, 53–72. [Google Scholar] [CrossRef]
- Quillévéré, F.; Nouailhat, N.; Joannin, S.; Cornée, J.-J.; Moissette, P.; Lécuyer, C.; Fourel, F.; Agiadi, K.; Koskeridou, E.; Escarguel, G. An onshore bathyal record of tectonics and climate cycles at the onset of the Early-Middle Pleistocene Transition in the eastern Mediterranean. Quat. Sci. Rev. 2019, 209, 23–39. [Google Scholar] [CrossRef]
- Siani, G.; Paterne, M.; Colin, C. Late glacial to Holocene planktic foraminifera bioevents and climatic record in the South Adriatic Sea. J. Quat. Sci. 2010, 25, 808–821. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Mortyn, P.G.; Antonarakou, A.; Martínez-Botí, M.A.; Triantaphyllou, M.V. Field-based validation of a diagenetic effect on G. ruber Mg/Ca paleothermometry: Core top results from the Aegean Sea (eastern Mediterranean). Geochem. Geophys. Geosyst. 2011, 12, Q09004. [Google Scholar] [CrossRef]
- Vasiliev, I.; Karakitsios, V.; Bouloubassi, I.; Agiadi, K.; Kontakiotis, G.; Antonarakou, A.; Triantaphyllou, M.; Gogou, A.; Kafousia, N.; de Rafélis, M.; et al. Large Sea Surface Temperature, Salinity, and Productivity-Preservation Changes Preceding the Onset of the Messinian Salinity Crisis in the Eastern Mediterranean Sea. Paleoceanogr. Paleoclimatol. 2019, 34, 182–202. [Google Scholar] [CrossRef]
- Avnaim-Katav, S.; Herut, B.; Rahav, E.; Katz, T.; Weinstein, Y.; Alkalay, R.; Berman-Frank, I.; Zlatkin, O.; Almogi-Labin, A. Sediment trap and deep sea coretop sediments as tracers of recent changes in planktonic foraminifera assemblages in the southeastern ultra-oligotrophic Levantine Basin. Deep Sea Res. Part II Top. Stud. Oceanogr. 2020, 171, 104669. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Antonarakou, A.; Mortyn, P.G.; Drinia, H.; Anastasakis, G.; Zarkogiannis, S.; Möbius, J. Morphological recognition of Globigerinoides ruber morphotypes and their susceptibility to diagenetic alteration in the eastern Mediterranean Sea. J. Mar. Syst. 2017, 174, 12–24. [Google Scholar] [CrossRef]
- Pujol, C.; Grazzini, C.V. Distribution patterns of live planktic foraminifers as related to regional hydrography and productive systems of the Mediterranean Sea. Mar. Micropaleontol. 1995, 25, 187–217. [Google Scholar] [CrossRef]
- Wilson, B. Biogeography and ecostratigraphy of Late Quaternary planktonic foraminiferal taphocoenoses in the Leeward Islands, Lesser Antilles, NE Caribbean Sea. Mar. Micropaleontol. 2012, 86–87, 1–10. [Google Scholar] [CrossRef]
- Zarkogiannis, S.; Kontakiotis, G.; Antonarakou, A. Recent planktonic foraminifera population and size response to Eastern Mediterranean hudrography. Rev. Micropaleontol. 2020, 69, 100450. [Google Scholar] [CrossRef]
- Bé, A.W.H.; Tolderlund, D.S. Distribution and ecology of living planktonic foraminifera in surface waters of the Atlantic and Indian Oceans. In The Micropaleontology of Oceans; Funnel, B.M., Riedel, W.R., Eds.; Cambridge University Press: Cambridge, UK, 1971; pp. 105–149. [Google Scholar]
- Kucera, M. Chapter Six Planktonic Foraminifera as Tracers of Past Oceanic Environments. In Developments in Marine Geology; Hillaire–Marcel, C., De Vernal, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, pp. 213–262. [Google Scholar]
- Imbrie, J.; Kipp, N. A new micropaleontological method for quantitative paleoclimatology: Application to a late pleistocene Caribbean core. In The Late Cenozoic Glacial Ages; Turekian, K.K., Ed.; Yale University Press: New Haven, CT, USA, 1971; pp. 71–181. [Google Scholar]
- Kontakiotis, G. Late Quaternary paleoenvironmental reconstruction and paleoclimatic implications of the Aegean Sea (eastern Mediterranean) based on paleoceanographic indexes and stable isotopes. Quat. Int. 2016, 401, 28–42. [Google Scholar] [CrossRef] [Green Version]
- Kucera, M.; Weinelt, M.; Kiefer, T.; Pflaumann, U.; Hayes, A.; Weinelt, M.; Chen, M.-T.; Mix, A.C.; Barrows, T.T.; Cortijo, E.; et al. Reconstruction of sea-surface temperatures from assemblages of planktonic foraminifera: Multi-technique approach based on geographically constrained calibration data sets and its application to glacial Atlantic and Pacific Oceans. Quat. Sci. Rev. 2005, 24, 951–998. [Google Scholar] [CrossRef]
- Berger, W.H. Kummerform foraminifera as clues to Oceanic environments: Abstract. Am. Assoc. Pet. Geol. Bull. 1969, 53. [Google Scholar] [CrossRef]
- Moller, T.; Schulz, H.; Kucera, M. The effect of sea surface properties on shell morphology and size of the planktonic foraminifer Neogloboquadrina pachyderma in the North Atlantic. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 391, 34–48. [Google Scholar] [CrossRef]
- Morey, A.; Mix, A.; Pisias, N. Planktonic foraminiferal assemblages preserved in surface sediments correspond to multiple environment variables. Quat. Sci. Rev. 2005, 24, 925–950. [Google Scholar] [CrossRef]
- Ortiz, J.D.; Mix, A.C.; Collier, R.W. Environmental control of living symbiotic and asymbiotic foraminifera of the California Current. Paleoceanography 1995, 10, 987–1009. [Google Scholar] [CrossRef] [Green Version]
- Rebotim, A.; Voelker, A.H.L.; Jonkers, L.; Waniek, J.J.; Meggers, H.; Schiebel, R.; Fraile, I.; Schulz, M.; Kucera, M. Factors controlling the depth habitat of planktonic foraminifera in the subtropical eastern North Atlantic. Biogeosci. Discuss. 2016, 2016, 1–48. [Google Scholar] [CrossRef] [Green Version]
- Renaud, S.; Schmidt, D. Habitat tracking as a response of the planktic foraminifer Globorotalia truncatulinoides to environmental fluctuations during the last 140 kyr. Mar. Micropaleontol. 2003, 49, 97–122. [Google Scholar] [CrossRef]
- Schiebel, R.; Waniek, J.; Bork, M.; Hemleben, C. Planktic foraminiferal production stimulated by chlorophyll redistribution and entrainment of nutrients. Deep Sea Res. Part I Oceanogr. Res. Pap. 2001, 48, 721–740. [Google Scholar] [CrossRef]
- Schmidt, D.N.; Renaud, S.; Bollmann, J.; Schiebel, R.; Thierstein, H.R. Size distribution of Holocene planktic foraminifer assemblages: Biogeography, ecology and adaptation. Mar. Micropaleontol. 2004, 50, 319–338. [Google Scholar] [CrossRef]
- Weinkauf, M.F.G.; Kunze, J.G.; Waniek, J.J.; Kučera, M. Seasonal variation in shell calcification of planktonic foraminifera in the NE Atlantic reveals species-specific response to temperature, productivity, and optimum growth conditions. PLoS ONE 2016, 11, e0148363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siokou-Frangou, I.; Bianchi, M.; Christaki, U.; Christou, E.D.; Giannakourou, A.; Gotsis, O.; Ignatiades, L.; Pagou, K.; Pitta, P.; Psarra, S.; et al. Carbon flow in the planktonic food web along a gradient of oligotrophy in the Aegean Sea (Mediterranean Sea). J. Mar. Syst. 2002, 33–34, 335–353. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the open Mediterranean Sea: A review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef] [Green Version]
- Wassmann, P.; Ypma, J.E.; Tselepides, A. Vertical flux of faecal pellets and microplankton on the shelf of the oligotrophic Cretan Sea (NE Mediterranean Sea). Prog. Oceanogr. 2000, 46, 241–258. [Google Scholar] [CrossRef]
- Schmuker, B.; Schiebel, R. Planktic foraminifers and hydrography of the eastern and northern Caribbean Sea. Mar. Micropaleontol. 2002, 46, 387–403. [Google Scholar] [CrossRef]
- Lykousis, V.; Chronis, G.; Tselepides, A.; Price, N.B.; Theocharis, A.; Siokou-Frangou, I.; Van Wambeke, F.; Danovaro, R.; Stavrakakis, S.; Duineveld, G.; et al. Major outputs of the recent multidisciplinary biogeochemical researches undertaken in the Aegean Sea. J. Mar. Syst. 2002, 33–34, 313–334. [Google Scholar] [CrossRef]
- Schiebel, R. Planktic foraminiferal sedimentation and the marine calcite budget. Glob. Biogeochem. Cycles 2002, 16, 1065. [Google Scholar] [CrossRef]
- Schiebel, R.; Hemleben, C. Modern planktic foraminifera. Palaontol. Z. 2005, 79, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Siccha, M.; Schiebel, R.; Schmidt, S.; Howa, H. Short-term and small-scale variability in planktic foraminifera test flux in the Bay of Biscay. Deep Sea Res. Part I Oceanogr. Res. Pap. 2012, 64, 146–156. [Google Scholar] [CrossRef] [Green Version]
- Bijma, J.; Faber, W.W.; Hemleben, C. Temperature and salinity limits for growth and survival of some planktonic foraminifers in laboratory cultures. J. Foraminifer. Res. 1990, 20, 95–116. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.J.; Parker, W.C. Biogeography of Planktonic Foraminifera; Springer: Dortrecht, The Netherlands; Boston, MA, USA; London, UK, 1999. [Google Scholar]
- Zarkogiannis, S.; Kontakiotis, G.; Antonarakou, A.; Mortyn, P.; Drinia, H. Latitudinal Variation of Planktonic Foraminifera Shell Masses During Termination I. Iop Conf. Ser. Earth Environ. Sci. 2019, 221, 012052. [Google Scholar] [CrossRef]
- Zarkogiannis, S.D.; Antonarakou, A.; Tripati, A.; Kontakiotis, G.; Mortyn, P.G.; Drinia, H.; Greaves, M. Influence of surface ocean density on planktonic foraminifera calcification. Sci. Rep. 2019, 9, 533. [Google Scholar] [CrossRef]
- Caromel, A.G.M.; Schmidt, D.N.; Phillips, J.C.; Rayfield, E.J. Hydrodynamic constraints on the evolution and ecology of planktic foraminifera. Mar. Micropaleontol. 2014, 106, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Caromel, A.G.M.; Schmidt, D.N.; Rayfield, E.J. Ontogenetic constraints on foraminiferal test construction. Evol. Dev. 2017, 19, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Zarkogiannis, S.; Kontakiotis, G.; Antonarakou, A. Logarithmic expression of Globigerina bulloides shell evolution through the biometric analysis: Paleoceanographic implications for the late Quaternary. Iop Conf. Ser. Earth Environ. Sci. 2019, 362, 012100. [Google Scholar] [CrossRef] [Green Version]
- Robinson, A.R.; Golnaraghi, M. The Physical and Dynamical Oceanography of the Mediterranean Sea. In Ocean Processes in Climate Dynamics: Global and Mediterranean Examples; Malanotte-Rizzoli, P., Robinson, A.R., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 255–306. [Google Scholar]
- Cushman-Roisin, B.; Gacic, M.; Poulain, P.-M.; Artegiani, A. Physical Oceanography of the Adriatic Sea: Past, Present and Future; Springer: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Ricci, P.; Libralato, S.; Capezzuto, F.; D’Onghia, G.; Maiorano, P.; Sion, L.; Tursi, A.; Solidoro, C.; Carlucci, R. Ecosystem functioning of two marine food webs in the North-Western Ionian Sea (Central Mediterranean Sea). Ecol. Evol. 2019, 9, 10198–10212. [Google Scholar] [CrossRef]
- Civitarese, G.; Gacic, M.; Lipizer, M.; Eusebi Borzelli, G.L. On the impact of the Bimodal Oscillating System (BiOS) on the biogeochemistry and biology of the Adriatic and Ionian Seas (Eastern Mediterranean). Biogeosciences 2010, 7, 3987–3997. [Google Scholar] [CrossRef] [Green Version]
- Specchiulli, A.; Bignami, F.; Marini, M.; Fabbrocini, A.; Scirocco, T.; Campanelli, A.; Penna, P.; Santucci, A.; D’Adamo, R. The role of forcing agents on biogeochemical variability along the southwestern Adriatic coast: The Gulf of Manfredonia case study. Estuar. Coast. Shelf Sci. 2016, 183, 136–149. [Google Scholar] [CrossRef]
- Béranger, K.; Mortier, L.; Crépon, M. Seasonal variability of water transport through the Straits of Gibraltar, Sicily and Corsica, derived from a high-resolution model of the Mediterranean circulation. Prog. Oceanogr. 2005, 66, 341–364. [Google Scholar] [CrossRef]
- Pinardi, N.; Masetti, E. Variability of the large scale general circulation of the Mediterranean Sea from observations and modelling: A review. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 158, 153–173. [Google Scholar] [CrossRef]
- Robinson, A.R.; Sellschopp, J.; Warn-Varnas, A.; Leslie, W.G.; Lozano, C.J.; Haley, P.J.; Anderson, L.A.; Lermusiaux, P.F.J. The Atlantic Ionian Stream. J. Mar. Syst. 1999, 20, 129–156. [Google Scholar] [CrossRef]
- Bonanno, A.; Placenti, F.; Basilone, G.; Mifsud, R.; Genovese, S.; Patti, B.; Di Bitetto, M.; Aronica, S.; Barra, M.; Giacalone, G.; et al. Variability of water mass properties in the Strait of Sicily in summer period of 1998-2013. Ocean Sci. Discuss. 2014, 11. [Google Scholar] [CrossRef]
- Sammari, C.; Millot, C.; Taupier-Letage, I.; Stefani, A.; Brahim, M. Hydrological characteristics in the Tunisia–Sardinia–Sicily area during spring 1995. Deep Sea Res. Part I Oceanogr. Res. Pap. 1999, 46, 1671–1703. [Google Scholar] [CrossRef]
- Gačić, M.; Schroeder, K.; Civitarese, G.; Cosoli, S.; Vetrano, A.; Eusebi Borzelli, G.L. Salinity in the Sicily Channel corroborates the role of the Adriatic–Ionian Bimodal Oscillating System (BiOS) in shaping the decadal variability of the Mediterranean overturning circulation. Ocean Sci. 2013, 9, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Pinardi, N.; Zavatarelli, M.; Adani, M.; Coppini, G.; Fratianni, C.; Oddo, P.; Simoncelli, S.; Tonani, M.; Lyubartsev, V.; Dobricic, S.; et al. Mediterranean Sea large-scale low-frequency ocean variability and water mass formation rates from 1987 to 2007: A retrospective analysis. Prog. Oceanogr. 2015, 132, 318–332. [Google Scholar] [CrossRef]
- Roether, W.; Manca, B.B.; Klein, B.; Bregant, D.; Georgopoulos, D.; Beitzel, V.; Kovačević, V.; Luchetta, A. Recent Changes in Eastern Mediterranean Deep Waters. Science 1996, 271, 333. [Google Scholar] [CrossRef]
- Gasparini, G.P.; Ortona, A.; Budillon, G.; Astraldi, M.; Sansone, E. The effect of the Eastern Mediterranean Transient on the hydrographic characteristics in the Strait of Sicily and in the Tyrrhenian Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2005, 52, 915–935. [Google Scholar] [CrossRef]
- Marini, M.; Grilli, F.; Guarnieri, A.; Jones, B.; Klajic, Z.; Nadia, P.; Sanxhaku, M. Is the southeastern Adriatic Sea coastal strip an eutrophic area? Estuar. Coast. Shelf Sci. 2010, 395–406. [Google Scholar] [CrossRef]
- Artegiani, A.; Paschini, E.; Russo, A.; Bregant, D.; Raicich, F.; Pinardi, N. The Adriatic Sea General Circulation. Part II: Baroclinic Circulation Structure. J. Phys. Oceanogr. 1997, 27, 1515–1532. [Google Scholar] [CrossRef]
- Marini, M.; Jones, B.H.; Campanelli, A.; Grilli, F.; Lee, C.M. Seasonal variability and Po River plume influence on biochemical properties along western Adriatic coast. J. Geophys. Res. Ocean. 2008, 113. [Google Scholar] [CrossRef]
- Orlic, M.; Dadic, V.; Grbec, B.; Leder, N.; Marki, A.; Matić, F.; Mihanovic, H.; Beg Paklar, G.; Pasaric, M.; Pasarić, Z.; et al. Wintertime buoyancy forcing, changing seawater properties, and two different circulation systems produced in the Adriatic. J. Geophys. Res. 2006, 112. [Google Scholar] [CrossRef] [Green Version]
- Giani, M.; Djakovac, T.; Degobbis, D.; Cozzi, S.; Solidoro, C.; Umani, S.F. Recent changes in the marine ecosystems of the northern Adriatic Sea. Estuar. Coast. Shelf Sci. 2012, 115, 1–13. [Google Scholar] [CrossRef]
- Manca, B.; Budillon, G.; Scarazzato, P.; Ursella, L. Evolution of dynamics in the eastern Mediterranean affecting water mass structures and properties in the Ionian and Adriatic Seas. J. Geophys. Res. 2003, 108. [Google Scholar] [CrossRef] [Green Version]
- Degobbis, D.; Precali, R.; Ivančić, I.; Smodlaka, N.; Fuks, D.; Kveder, S. Long-term Changes in the northern Adriatic ecosystem related to anthropogenic eutrophication. Int. J. Environ. Pollut. 2000, 13, 495–533. [Google Scholar] [CrossRef]
- Viličič, D.; Vučak, Z.; Škrivanić, A.; Grzetić, Z. Phytoplankton blooms in the oligotrophic open South Adriatic waters. Mar. Chem. 1989, 28, 89–107. [Google Scholar] [CrossRef]
- Totti, C.; Civitarese, G.; Acri, F.; Barletta, D.; Candelari, G.; Paschini, E.; Solazzi, A. Seasonal variability of phytoplankton populations in the middle Adriatic sub-basin. J. Plankton Res. 2000, 22, 1735–1756. [Google Scholar] [CrossRef] [Green Version]
- Zonneveld, K.; Chen, L.; Moebius, J.; Mahmoud, M. Environmental significance of dinoflagellate cysts from the proximal part of the Po-river discharge plume (off southern Italy, Eastern Mediterranean). J. Sea Res. 2009, 62, 189–213. [Google Scholar] [CrossRef]
- Marasović, I.; Grbec, B.; Morović, M. Long-term production changes in the adriatic. Neth. J. Sea Res. 1995, 34, 267–273. [Google Scholar] [CrossRef]
- Vilibić, I.; Šantić, D. Deep water ventilation traced by Synechococcus cyanobacteria. Ocean Dyn. 2008, 58, 119–125. [Google Scholar] [CrossRef]
- Vilibić, I.; Matijevic, S.; Šepić, J.; Kušpilić, G. Changes in the Adriatic oceanographic properties induced by the Eastern Mediterranean Transient. Biogeosciences 2012, 9, 2085–2097. [Google Scholar] [CrossRef] [Green Version]
- Palinkas, C.M.; Nittrouer, C.A. Clinoform sedimentation along the Apennine shelf, Adriatic Sea. Mar. Geol. 2006, 234, 245–260. [Google Scholar] [CrossRef]
- Boudena, M. Sea Surface Temperature of the Central Mediterranean during the Last Millenium. Ph.D. Thesis, Università degli Studi di Torino, Turin, Italy, 2004; p. 64. [Google Scholar]
- Frignani, M.; Langone, L.; Ravaioli, M.; Sorgente, D.; Alvisi, F.; Albertazzi, S. Fine-sediment mass balance in the western Adriatic continental shelf over a century time scale. Mar. Geol. 2005, 222–223, 113–133. [Google Scholar] [CrossRef]
- Incarbona, A.; Ziveri, P.; Di Stefano, E.; Lirer, F.; Mortyn, G.; Patti, B.; Pelosi, N.; Sprovieri, M.; Tranchida, G.; Vallefuoco, M.; et al. The Impact of the Little Ice Age on Coccolithophores in the Central Mediterranea Sea. Clim. Past 2010, 6, 795–805. [Google Scholar] [CrossRef] [Green Version]
- Grauel, A.L.; Bernasconi, S.M. Core-top calibration of δ18O and δ13C of G. ruber (white) and U. mediterranea along the southern Adriatic coast of Italy. Mar. Micropaleontol. 2010, 77, 175–186. [Google Scholar] [CrossRef]
- Thunell, R.C. Distribution of recent planktonic foraminifera in surface sediments of the Mediterranean Sea. Mar. Micropaleontol. 1978, 3, 147–173. [Google Scholar] [CrossRef]
- Zarkogiannis, S.D.; Kontakiotis, G.; Gkaniatsa, G.; Kuppili, V.S.C.; Marathe, S.; Wanelik, K.; Lianou, V.; Besiou, E.; Makri, P.; Antonarakou, A. An Improved Cleaning Protocol for Foraminiferal Calcite from Unconsolidated Core Sediments: HyPerCal—A New Practice for Micropaleontological and Paleoclimatic Proxies. J. Mar. Sci. Eng. 2020, 8, 998. [Google Scholar] [CrossRef]
- Incarbona, A.; Stefano, E.; Sprovieri, R.; Ferraro, S. The Uniqueness of Planktonic Ecosystems in the Mediterranean Sea: The Response to Orbital- and Suborbital-Climatic Forcing over the Last 130,000 Years. Open Geosci. 2016, 8. [Google Scholar] [CrossRef]
- Mallo, M.; Ziveri, P.; Mortyn, P.G.; Schiebel, R.; Grelaud, M. Low planktic foraminiferal diversity and abundance observed in a spring 2013 west–east Mediterranean Sea plankton tow transect. Biogeosciences 2017, 14, 2245–2266. [Google Scholar] [CrossRef] [Green Version]
- Al-Sabouni, N.; Kucera, M.; Schmidt, D.N. Vertical niche separation control of diversity and size disparity in planktonic foraminifera. Mar. Micropaleontol. 2007, 63, 75–90. [Google Scholar] [CrossRef]
- Antonarakou, A.; Kontakiotis, G.; Zarkogiannis, S.; Mortyn, P.G.; Drinia, H.; Koskeridou, E.; Anastasakis, G. Planktonic foraminiferal abnormalities in coastal and open marine eastern Mediterranean environments: A natural stress monitoring approach in recent and early Holocene marine systems. J. Mar. Syst. 2018, 181, 63–78. [Google Scholar] [CrossRef]
- Capotondi, L.; Erica, S.; Speranza, P.; Corselli, C. Late Quaternary planktonic foraminiferal distributions: Problems related to size fraction. In Proceedings of the First Italian Meeting on Environmental Micropaleontology Edition; Coccioni, R., Galeotti, S., Lirer, F., Eds.; Grzybowski Foundation Special Publication: Krakow, Poland, 2004; Volume 9, pp. 1–6. [Google Scholar]
- Cisneros, M.; Cacho, I.; Frigola, J.; Canals, M.; Masqué, P.; Martrat, B.; Casado, M.; Grimalt, J.O.; Pena, L.D.; Margaritelli, G.; et al. Sea surface temperature variability in the central-western Mediterranean Sea during the last 2700 years: A multi-proxy and multi-record approach. Clim. Past 2016, 12, 849–869. [Google Scholar] [CrossRef] [Green Version]
- Margaritelli, G.; Cisneros, M.; Cacho, I.; Capotondi, L.; Vallefuoco, M.; Rettori, R.; Lirer, F. Climatic variability over the last 3000 years in the central- western Mediterranean Sea (Menorca Basin) detected by planktonic foraminifera and stable isotope records. Glob. Planet. Chang. 2018, 169, 179–187. [Google Scholar] [CrossRef]
- Schiebel, R.; Hemleben, C. Planktic Foraminifers in the Modern Ocean; Springer: Berlin/Heidelberg, Germany, 2017; p. 358. [Google Scholar]
- Aurahs, R.; Grimm, G.W.; Hemleben, V.; Hemleben, C.; Kucera, M. Geographical distribution of cryptic genetic types in the planktonic foraminifer Globigerinoides ruber. Mol. Ecol. 2009, 18, 1692–1706. [Google Scholar] [CrossRef]
- Wang, L. Isotopic signals in two morphotypes of Globigerinoides ruber (white) from the South China Sea: Implications for monsoon climate change during the last glacial cycle. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 161, 381–394. [Google Scholar] [CrossRef]
- Kuroyanagi, A.; Kawahata, H. Vertical distribution of living planktonic foraminifera in the seas around Japan. Mar. Micropaleontol. 2004, 53, 173–196. [Google Scholar] [CrossRef]
- Antonarakou, A.; Kontakiotis, G.; Mortyn, P.G.; Drinia, H.; Sprovieri, M.; Besiou, E.; Tripsanas, E. Biotic and geochemical (δ18O, δ13C, Mg/Ca, Ba/Ca) responses of Globigerinoides ruber morphotypes to upper water column variations during the last deglaciation, Gulf of Mexico. Geochim. Cosmochim. Acta 2015, 170. [Google Scholar] [CrossRef]
- Kuroyanagi, A.; Tsuchiya, M.; Kawahata, H.; Kitazato, H. The occurrence of two genotypes of the planktonic foraminifer Globigerinoides ruber (white) and paleo-environmental implications. Mar. Micropaleontol. 2008, 68, 236–243. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Antonarakou, A.; Zachariasse, W.J. Late Quaternary palaeoenvironmental changes in the Aegean Sea: Interrelations and interactions between North and South Aegean Sea. Bull. Geol. Soc. Greece 2013, 47, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Lirer, F.; Sprovieri, M.; Vallefuoco, M.; Ferraro, L.; Pelosi, N.; Giordano, L.; Capotondi, L. Planktonic foraminifera as bio-indicators for monitoring the climatic changes that have occurred over the past 2000 years in the southeastern Tyrrhenian Sea. Integr. Zool. 2014, 9, 542–554. [Google Scholar] [CrossRef]
- Rohling, E.J.; Jorissen, F.J.; Grazzini, C.V.; Zachariasse, W.J. Northern Levantine and Adriatic Quaternary planktic foraminifera; Reconstruction of paleoenvironmental gradients. Mar. Micropaleontol. 1993, 21, 191–218. [Google Scholar] [CrossRef]
- Marshall, B.J.; Thunell, R.C.; Henehan, M.J.; Astor, Y.; Wejnert, K.E. Planktonic foraminiferal area density as a proxy for carbonate ion concentration: A calibration study using the Cariaco Basin ocean time series. Paleoceanography 2013, 28, 363–376. [Google Scholar] [CrossRef]
- Kucera, M.; Kennett, J. Causes and consequences of a Middle Pleistocene origin of the modern planktonic foraminifer Neogloboquadrina pachyderma sinistral. Geology 2002, 30, 539–542. [Google Scholar] [CrossRef]
- Schneider, A.; Wallace, D.W.R.; Körtzinger, A. Alkalinity of the Mediterranean Sea. Geophys. Res. Lett. 2007, 34, L15608. [Google Scholar] [CrossRef] [Green Version]
- Ujiié, Y.; Asami, T.; de Garidel-Thoron, T.; Liu, H.; Ishitani, Y.; de Vargas, C. Longitudinal differentiation among pelagic populations in a planktic foraminifer. Ecol. Evol. 2012, 2, 1725–1737. [Google Scholar] [CrossRef] [Green Version]
- Kontakiotis, G. Palaeoceanographic and Palaeoclimatic Study of Eastern Mediterranean During Late Quaternary, Based on Planktonic Foraminiferal Assemblages. Ph.D. Thesis, National and Kapodistrian University of Athens, Athens, Greece, 2012. (In Greek, with English extended abstract). [Google Scholar]
- Rigual-Hernández, A.S.; Sierro, F.J.; Bárcena, M.A.; Flores, J.A.; Heussner, S. Seasonal and interannual changes of planktic foraminiferal fluxes in the Gulf of Lions (NW Mediterranean) and their implications for paleoceanographic studies: Two 12-year sediment trap records. Deep Sea Res. Part I Oceanogr. Res. Pap. 2012, 66, 26–40. [Google Scholar] [CrossRef]
- Tolderlund, D.S.; Allan, W.H.B. Seasonal Distribution of Planktonic Foraminifera in the Western North Atlantic. Micropaleontology 1971, 17, 297–329. [Google Scholar] [CrossRef]
- Numberger, L.; Hemleben, C.; Hoffmann, R.; Mackensen, A.; Schulz, H.; Wunderlich, J.-M.; Kucera, M. Habitats, abundance patterns and isotopic signals of morphotypes of the planktonic foraminifer Globigerinoides ruber (d’Orbigny) in the eastern Mediterranean Sea since the Marine Isotopic Stage 12. Mar. Micropaleontol. 2009, 73, 90–104. [Google Scholar] [CrossRef]
- Richey, J.N.; Poore, R.Z.; Flower, B.P.; Hollander, D.J. Ecological controls on the shell geochemistry of pink and white Globigerinoides ruber in the northern Gulf of Mexico: Implications for paleoceanographic reconstruction. Mar. Micropaleontol. 2012, 82–83, 28–37. [Google Scholar] [CrossRef]
- Wejnert, K.E.; Pride, C.J.; Thunell, R.C. The oxygen isotope composition of planktonic foraminifera from the Guaymas Basin, Gulf of California: Seasonal, annual, and interspecies variability. Mar. Micropaleontol. 2010, 74, 29–37. [Google Scholar] [CrossRef]
- D’Ortenzio, F.; Ragni, M.; Marullo, S.; Ribera d’Alcalà, M. Did biological activity in the Ionian Sea change after the Eastern Mediterranean Transient? Results from the analysis of remote sensing observations. J. Geophys. Res. Ocean. 2003, 108. [Google Scholar] [CrossRef] [Green Version]
- D’Ortenzio, F.; Ribera d’Alcalà, M. On the trophic regimes of the Mediterranean Sea: A satellite analysis. Biogeosciences 2009, 6, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Lavigne, H.; D’Ortenzio, F.; Ribera D’Alcalà, M.; Claustre, H.; Sauzède, R.; Gacic, M. On the vertical distribution of the chlorophyll a concentration in the Mediterranean Sea: A basin-scale and seasonal approach. Biogeosciences 2015, 12, 5021–5039. [Google Scholar] [CrossRef] [Green Version]
- Gotsis-Skretas, O.; Pagou, K.; Moraitou-Apostolopoulou, M.; Ignatiades, L. Seasonal horizontal and vertical variability in primary production and standing stocks of phytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc (March 1994–January 1995). Prog. Oceanogr. 1999, 44, 625–649. [Google Scholar] [CrossRef]
- Malinverno, E.; Maffioli, P.; Corselli, C.; De Lange, G.J. Present-day fluxes of coccolithophores and diatoms in the pelagic Ionian Sea. J. Mar. Syst. 2014, 132, 13–27. [Google Scholar] [CrossRef]
- Grilli, F.; Marini, M.; Book, J.W.; Campanelli, A.; Paschini, E.; Russo, A. Flux of nutrients between the middle and southern Adriatic Sea (Gargano-Split section). Mar. Chem. 2013, 153, 1–14. [Google Scholar] [CrossRef]
- Takagi, H.; Kimoto, K.; Fujiki, T.; Saito, H.; Schmidt, C.; Kucera, M.; Moriya, K. Characterizing photosymbiosis in modern planktonic foraminifera. Biogeosciences 2019, 16, 3377–3396. [Google Scholar] [CrossRef] [Green Version]
- Rohling, E.J.; Jorissen, F.J.; De Stigter, H.C. 200 Year interruption of Holocene sapropel formation in the Adriatic Sea. J. Micropalaeontol. 1997, 16, 97–108. [Google Scholar] [CrossRef]
- Žarić, S.; Donner, B.; Fischer, G.; Mulitza, S.; Wefer, G. Sensitivity of planktic foraminifera to sea surface temperature and export production as derived from sediment trap data. Mar. Micropaleontol. 2005, 55, 75–105. [Google Scholar] [CrossRef]
- Rohling, E.J.; Sprovieri, M.; Cane, T.; Casford, J.S.L.; Cooke, S.; Bouloubassi, I.; Emeis, K.C.; Schiebel, R.; Rogerson, M.; Hayes, A.; et al. Reconstructing past planktic foraminiferal habitats using stable isotope data: A case history for Mediterranean sapropel S5. Mar. Micropaleontol. 2004, 50, 89–123. [Google Scholar] [CrossRef]
- Rasmussen, T.L.; Thomsen, E. Changes in planktic foraminiferal faunas, temperature and salinity in the Gulf Stream during the last 30,000 years: Influence of meltwater via the Mississippi River. Quat. Sci. Rev. 2012, 33, 42–54. [Google Scholar] [CrossRef]
- Schiebel, R.; Hemleben, C. Interannual variability of planktic foraminiferal populations and test flux in the eastern North Atlantic Ocean (JGOFS). Deep Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 1809–1852. [Google Scholar] [CrossRef]
- Machain, M.; Monreal-Gomez, M.; Arellano-Torres, E.; Merino-Ibarra, M.; Gonzalez-Chavez, G. Recent planktonic foraminiferal distribution patterns and their relation to hydrographic conditions of the Gulf of Tehuantepec, Mexican Pacific. Mar. Micropaleontol. 2008, 66, 103–119. [Google Scholar] [CrossRef]
- Casford, J.S.L.; Rohling, E.J.; Abu-Zied, R.; Cooke, S.; Fontanier, C.; Leng, M.; Lykousis, V. Circulation changes and nutrient concentrations in the late Quaternary Aegean Sea: A nonsteady state concept for sapropel formation. Paleoceanography 2002, 17, 14-1–14-11. [Google Scholar] [CrossRef] [Green Version]
- Hecht, A.D. Size variations in planktonic foraminifera: Implications for quantitative paleoclimatic analysis. Science 1976, 192, 1330–1332. [Google Scholar] [CrossRef]
- Schmidt, D.N.; Lazarus, D.; Young, J.R.; Kucera, M. Biogeography and evolution of body size in marine plankton. Earth-Sci. Rev. 2006, 78, 239–266. [Google Scholar] [CrossRef]
- Schmidt, D.N.; Thierstein, H.R.; Bollmann, J.; Schiebel, R. Abiotic forcing of plankton evolution in the Cenozoic. Science 2004, 303, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Bé, A.W.H.; Harrison, S.M.; Lott, L. Orbulina universa d’Orbigny in the Indian Ocean. Micropaleontology 1973, 19, 150–192. [Google Scholar] [CrossRef]
- Hecht, A.D. An ecologic model for test size variation in recent planktonic foraminifera: Applications to the fossil record. J. Foraminifer. Res. 1976, 6, 295–311. [Google Scholar] [CrossRef]
- Kennett, J.P. Phenotypic variation in some recent and late Cenozoic planktonic foraminifera. In Foraminifera; Hedley, R.H., Adams, C.G., Eds.; Academic Press: New York, NY, USA, 1976; Volume 2, pp. 111–170. [Google Scholar]
- Malmgren, B.A.; Kennett, J.P. Size variations in Globigerina bulloides d’Orbigny as a Quaternary paleoclimatic index in the Southern Ocean. Antarct. J. 1976, 177–178. [Google Scholar]
- Malmgren, B.A.; Kennett, J.P. Biometric differentiation between recent Globigerina bulloides and Globigerina falconensis in the southern Indian Ocean. J. Foraminifer. Res. 1977, 7, 130–148. [Google Scholar] [CrossRef]
- Rillo, M.C.; Miller, C.G.; Kučera, M.; Ezard, T.H.G. Predictability of intraspecific size variation in extant planktonic foraminifera. bioRxiv 2018, 468165. [Google Scholar] [CrossRef] [Green Version]
- Peter, K.; Sommer, U. Phytoplankton Cell Size Reduction in Response to Warming Mediated by Nutrient Limitation. PLoS ONE 2013, 8, e71528. [Google Scholar] [CrossRef] [Green Version]
- Bé, A.W.H.; Caron, D.A.; Anderson, O.R. Effects of feeding frequency on life processes of the planktonic foraminifer Globigerinoides sacculifer in laboratory culture. J. Mar. Biol. Assoc. UK 1981, 61, 257–277. [Google Scholar] [CrossRef]
- Takagi, H.; Kimoto, K.; Fujiki, T.; Moriya, K. Effect of nutritional condition on photosymbiotic consortium of cultured Globigerinoides sacculifer (Rhizaria, Foraminifera). Symbiosis 2018, 76, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Stambler, N. The Mediterranean Sea–Primary Productivity. In The Mediterranean Sea: Its History and Present Challenges; Goffredo, S., Dubinsky, Z., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 113–121. [Google Scholar]
- Bé, A.W.H.; Hutson, W.H. Ecology of Planktonic Foraminifera and Biogeographic Patterns of Life and Fossil Assemblages in the Indian Ocean. Micropaleontology 1977, 23, 369. [Google Scholar] [CrossRef]
- Darling, K.F.; Wade, C.M. The genetic diversity of planktic foraminifera and the global distribution of ribosomal RNA genotypes. Mar. Micropaleontol. 2008, 67, 216–238. [Google Scholar] [CrossRef]
- de Vargas, C.; Zaninetti, L.; Hilbrecht, H.; Pawlowski, J. Phylogeny and rates of molecular evolution of planktonic Foraminifera: SSU rDNA sequences compared to the fossil record. J. Mol. Evol. 1997, 45, 285–294. [Google Scholar] [CrossRef]
- Kucera, M.; Darling, K.F. Cryptic species of planktonic foraminifera: Their effect on palaeoceanographic reconstructions. Philos. Trans. Ser. A Math. Phys. Eng. Sci. 2002, 360, 695–718. [Google Scholar] [CrossRef] [PubMed]
- Parker, F.L. Planktonic Foraminiferal Species in Pacific Sediments. Micropaleontology 1962, 8, 219–254. [Google Scholar] [CrossRef]
- Darling, K.F.; Wade, C.M.; Kroon, D.; Brown, A.J.L.; Bijma, J. The Diversity and Distribution of Modern Planktic Foraminiferal Small Subunit Ribosomal RNA Genotypes and their Potential as Tracers of Present and Past Ocean Circulations. Paleoceanography 1999, 14, 3–12. [Google Scholar] [CrossRef] [Green Version]
- de Vargas, C.; Bonzon, M.; Rees, N.W.; Pawlowski, J.; Zaninetti, L. A molecular approach to biodiversity and biogeography in the planktonic foraminifer Globigerinella siphonifera (d’Orbigny). Mar. Micropaleontol. 2002, 45, 101–116. [Google Scholar] [CrossRef]
- Huber, R.; van Staaden, M.J.; Kaufman, L.S.; Liem, K.F. Microhabitat use, trophic patterns, and the evolution of brain structure in African cichlids. Brain Behav. Evol. 1997, 50, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Darling, K.F.; Wade, C.M.; Kroon, D.; Brown, A.J.L. Planktic foraminiferal molecular evolution and their polyphyletic origins from benthic taxa. Mar. Micropaleontol. 1997, 30, 251–266. [Google Scholar] [CrossRef]
- Morard, R.; Quillévéré, F.; Escarguel, G.; Ujiie, Y.; de Garidel-Thoron, T.; Norris, R.D.; de Vargas, C. Morphological recognition of cryptic species in the planktonic foraminifer Orbulina universa. Mar. Micropaleontol. 2009, 71, 148–165. [Google Scholar] [CrossRef]
- Vargas, C.d.; Norris, R.; Zaninetti, L.; Gibb, S.; Pawlowski, J. Molecular evidence of cryptic speciation in planktonic foraminifers and their relation to oceanic provinces. Proc. Natl. Acad. Sci. USA 1999, 96, 2864–2868. [Google Scholar] [CrossRef] [Green Version]
- Morard, R.; Quillévéré, F.; Douady, C.J.; de Vargas, C.; de Garidel-Thoron, T.; Escarguel, G. Worldwide genotyping in the planktonic foraminifer Globoconella inflata: Implications for life history and paleoceanography. PLoS ONE 2011, 6, e26665. [Google Scholar] [CrossRef] [Green Version]
- Aurahs, R.; Treis, Y.; Darling, K.; Kucera, M. A revised taxonomic and phylogenetic concept for the planktonic foraminifer species Globigerinoides ruber based on molecular and morphometric evidence. Mar. Micropaleontol. 2011, 79, 1–14. [Google Scholar] [CrossRef]
- Seears, H.A.; Darling, K.F.; Wade, C.M. Ecological partitioning and diversity in tropical planktonic foraminifera. BMC Evol. Biol. 2012, 12, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, A.K.; Weinkauf, M.F.; Kurasawa, A.; Darling, K.F.; Kucera, M.; Grimm, G.W. Phylogeography of the tropical planktonic foraminifera lineage globigerinella reveals isolation inconsistent with passive dispersal by ocean currents. PLoS ONE 2014, 9, e92148. [Google Scholar] [CrossRef] [PubMed]
- de Vargas, C.; Renaud, S.; Hilbrecht, H.; Pawlowski, J. Pleistocene Adaptive Radiation in Globorotalia truncatulinoides: Genetic, Morphologic, and Environmental Evidence. Paleobiology 2001, 27, 104–125. [Google Scholar] [CrossRef]
- Quillévéré, F.; Morard, R.; Escarguel, G.; Douady, C.J.; Ujiié, Y.; de Garidel-Thoron, T.; de Vargas, C. Global scale same-specimen morpho-genetic analysis of Truncorotalia truncatulinoides: A perspective on the morphological species concept in planktonic foraminifera. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 391, 2–12. [Google Scholar] [CrossRef]
- Darling, K.F.; Wade, C.M.; Stewart, I.A.; Kroon, D.; Dingle, R.; Brown, A.J. Molecular evidence for genetic mixing of Arctic and Antarctic subpolar populations of planktonic foraminifers. Nature 2000, 405, 43–47. [Google Scholar] [CrossRef]
- Stewart, I.A.; Darling, K.F.; Kroon, D.; Wade, C.M.; Troelstra, S.R. Genotypic variability in subarctic Atlantic planktic foraminifera. Mar. Micropaleontol. 2001, 43, 143–153. [Google Scholar] [CrossRef]
- André, A.; Quillévéré, F.; Schiebel, R.; Morard, R.; Howa, H.; Meilland, J.; Douady, C.J. Disconnection between genetic and morphological diversity in the planktonic foraminifer Neogloboquadrina pachyderma from the Indian sector of the Southern Ocean. Mar. Micropaleontol. 2018, 144, 14–24. [Google Scholar] [CrossRef]
- Bauch, D.; Darling, K.; Simstich, J.; Bauch, H.A.; Erlenkeuser, H.; Kroon, D. Palaeoceanographic implications of genetic variation in living North Atlantic Neogloboquadrina pachyderma. Nature 2003, 424, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Darling, K.F.; Kucera, M.; Kroon, D.; Wade, C.M. A resolution for the coiling direction paradox in Neogloboquadrina pachyderma. Paleoceanography 2006, 21, PA2011. [Google Scholar] [CrossRef] [Green Version]
- Darling, K.F.; Kucera, M.; Pudsey, C.J.; Wade, C.M. Molecular evidence links cryptic diversification in polar planktonic protists to Quaternary climate dynamics. Proc. Natl. Acad. Sci. USA 2004, 101, 7657. [Google Scholar] [CrossRef] [Green Version]
- Darling, K.F.; Kucera, M.; Wade, C.M. Global molecular phylogeography reveals persistent Arctic circumpolar isolation in a marine planktonic protist. Proc. Natl. Acad. Sci. USA 2007, 104, 5002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morard, R.; Vollmar, N.M.; Greco, M.; Kucera, M. Unassigned diversity of planktonic foraminifera from environmental sequencing revealed as known but neglected species. PLoS ONE 2019, 14, e0213936. [Google Scholar] [CrossRef] [PubMed]
- Beer, C.J.; Schiebel, R.; Wilson, P.A. Testing planktic foraminiferal shell weight as a surface water [CO32-] proxy using plankton net samples. Geology 2010, 38, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Deuser, W.G.; Ross, E.H.; Hemleben, C.; Spindler, M. Seasonal changes in species composition, numbers, mass, size, and isotopic composition of planktonic foraminifera settling into the deep sargasso sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1981, 33, 103–127. [Google Scholar] [CrossRef]
- Ortiz, J.D.; Mix, A.C. The spatial distribution and seasonal succession of planktonic foraminifera in the California Current off Oregon, September 1987–September 1988. Geol. Soc. Lond. Spec. Publ. 1992, 64, 197–213. [Google Scholar] [CrossRef]
- Salmon, K.H.; Anand, P.; Sexton, P.F.; Conte, M. Upper ocean mixing controls the seasonality of planktonic foraminifer fluxes and associated strength of the carbonate pump in the oligotrophic North Atlantic. Biogeosciences 2015, 12, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Poore, R.Z.; Spear, J.W.; Tedesco, K.A. Seasonal flux and assemblage composition of planktic foraminifers from a sediment-trap study in the northern Gulf of Mexico. J. Coast. Res. 2013, 6–19. [Google Scholar] [CrossRef]
- Chernihovsky, N.; Torfstein, A.; Almogi-Labin, A. Seasonal flux patterns of planktonic foraminifera in a deep, oligotrophic, marginal sea: Sediment trap time series from the Gulf of Aqaba, northern Red Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2018, 140, 78–94. [Google Scholar] [CrossRef]
- Jonkers, L.; Kučera, M. Global analysis of seasonality in the shell flux of extant planktonic Foraminifera. Biogeosciences 2015, 12, 2207–2226. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cruise | Station | Latitude | Longitude | Basin (Region) | Water Depth | Lithology | SST | SSS | Ch–a | Density σθ |
|---|---|---|---|---|---|---|---|---|---|---|
| (°N) | (°E) | (m) | °C | psu | kgCm−2yr−1 | (kg/m3) | ||||
| Poseidon P339 | #17. GeoB 10725 | 42.001 | 16.217 | Gargano Promontory (S. Adriatic Sea) | 94 | Mud | 14.40 | 38.29 | 0.42 | 28.65 |
| Poseidon P339 | #16. GeoB 10729 | 41.647 | 17.191 | Gargano Promontory (S. Adriatic Sea) | 708 | Sandy mud | 13.40 | 38.60 | 0.19 | 29.10 |
| Poseidon P339 | #15. GeoB 10730 | 41.500 | 17.050 | Gargano Promontory (S. Adriatic Sea) | 179 | Sandy mud | 13.90 | 38.70 | 0.21 | 29.07 |
| Poseidon P339 | #14. GeoB 10739 | 40.500 | 18.642 | Strait of Otranto (S. Adriatic Sea) | 561 | Mud | 13.80 | 38.73 | 0.24 | 29.12 |
| Poseidon P339 | #13. GeoB 10741 | 40.234 | 18.667 | Strait of Otranto (S. Adriatic Sea) | 286 | Sandy mud | 14.00 | 38.67 | 0.28 | 29.03 |
| Poseidon P339 | #12. GeoB 10718 | 39.693 | 18.058 | Gulf of Taranto (NW Ionian Sea) | 214 | Sandy mud | 14.20 | 38.32 | 0.27 | 28.71 |
| Poseidon P339 | #11. GeoB 10748 | 39.667 | 17.050 | Strait of Otranto (S. Adriatic Sea) | 284 | Mud | 13.80 | 38.73 | 0.25 | 29.12 |
| Poseidon P339 | #10. GeoB 10720 | 39.507 | 17.979 | Gulf of Taranto (NW Ionian Sea) | 1384 | Mud | 13.60 | 38.69 | 0.19 | 29.13 |
| Meteor M 71-3 | #9. H-11 | 39.283 | 19.333 | Offshore Kerkyra basin (NE. Ionian Sea) | 1032 | Muddy sand | 13.60 | 38.70 | 0.15 | 29.14 |
| Meteor M 71-3 | #8. H-07 | 39.167 | 17.750 | External Calabrian Arc (NW. Ionian Sea) | 1663 | Mud | 13.60 | 38.69 | 0.18 | 29.13 |
| Meteor M 71-3 | #7. H-12 | 38.833 | 19.750 | Offshore Kerkyra basin (NE. Ionian Sea) | 1459 | Clay | 13.60 | 38.70 | 0.14 | 29.14 |
| Meteor M 71-3 | #6. H-06 | 38.500 | 18.500 | Ionian Bathyal Plain (N. Ionian Sea) | 3018 | Clay | 13.80 | 38.71 | 0.14 | 29.10 |
| Meteor M 71-3 | #5. H-05 | 37.500 | 18.500 | Ionian Bathyal Plain (C. Ionian Sea) | 3157 | Mud | 13.90 | 38.72 | 0.12 | 29.09 |
| Meteor M 71-3 | #4. H-04 | 35.917 | 16.000 | External Calabrian Arc (W. Ionian Sea) | 3747 | Muddy sand | 13.80 | 38.67 | 0.14 | 29.07 |
| Meteor M 71-3 | #3. H-01 | 35.750 | 23.000 | Offshore Cretan basin (E. Ionian Sea) | 2121 | Clay | 14.10 | 38.83 | 0.11 | 29.13 |
| Meteor M 71-3 | #2. H-02 | 35.750 | 21.000 | Mediterranean Ridge (C. Ionian Sea) | 3005 | Clay | 13.90 | 38.74 | 0.09 | 29.10 |
| Meteor M 71-3 | #1. H-03 | 35.750 | 18.500 | Ionian Bathyal Plain (C. Ionian Sea) | 4088 | Clay | 13.80 | 38.67 | 0.10 | 29.07 |
| Species | ECD Range | Average ECD | ||
|---|---|---|---|---|
| Adriatic Sea | Ionian Sea | Adriatic Sea | Ionian Sea | |
| Globigerinoides ruber s.s. | 125–419 | 130–377 | 229 | 223 |
| Globigerinoides ruber s.l. | 162–417 | 157–469 | 308 | 269 |
| Globigerinoides ruber rosea | 135–574 | 126–579 | 344 | 302 |
| Trilobatus trilobus | 129–561 | 127–569 | 273 | 284 |
| Globigerinella siphonifera | 125–544 | 125–689 | 273 | 306 |
| Globigerina bulloides | 135–414 | 126–364 | 227 | 217 |
| Neogloboquadrina pachyderma | 130–392 | 127–400 | 213 | 234 |
| Globorotalia inflata | 162–551 | 171–612 | 303 | 357 |
| Globorotalia truncatulinoides | nd | 153–818 | nd | 435 |
| Globigerina rubescens | 130–207 | 128–259 | 169 | 191 |
| Orbulina universa | 153–814 | 174–905 | 414 | 496 |
| Turborotalita quinqueloba | 126–199 | 127–252 | 166 | 175 |
| Globigerinita glutinata | 126–211 | 125–315 | 165 | 197 |
| Species | ECD (μm) | St. Dev. (%) | No of Sites | No of Specimens | |
|---|---|---|---|---|---|
| Central Mediterranean | Eastern Mediterranean | ||||
| Globigerinoides ruber s.s. | 224 | 218 | 8 | 17 | 897 |
| Globigerinoides ruber s.l. | 279 | 259 | 11 | 17 | 592 |
| Globigerinoides ruber rosea | 313 | 338 | 15 | 16 | 192 |
| Trilobatus trilobus | 280 | 315 | 18 | 16 | 122 |
| Globigerinella siphonifera | 292 | 305 | 15 | 17 | 260 |
| Globigerina bulloides | 218 | 207 | 9 | 17 | 1463 |
| Neogloboquadrina pachyderma | 227 | 220 | 14 | 12 | 211 |
| Globorotalia inflata | 337 | 266 | 17 | 11 | 90 |
| Globorotalia truncatulinoides | 435 | nd | 20 | 5 | 20 |
| Globigerina rubescens | 184 | 189 | 11 | 13 | 140 |
| Orbulina universa | 471 | 427 | 19 | 17 | 180 |
| Turborotalita quinqueloba | 171 | 192 | 14 | 12 | 79 |
| Globigerinita glutinata | 188 | 189 | 13 | 13 | 118 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kontakiotis, G.; Efstathiou, E.; Zarkogiannis, S.D.; Besiou, E.; Antonarakou, A. Latitudinal Differentiation among Modern Planktonic Foraminiferal Populations of Central Mediterranean: Species–Specific Distribution Patterns and Size Variability. J. Mar. Sci. Eng. 2021, 9, 551. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9050551
Kontakiotis G, Efstathiou E, Zarkogiannis SD, Besiou E, Antonarakou A. Latitudinal Differentiation among Modern Planktonic Foraminiferal Populations of Central Mediterranean: Species–Specific Distribution Patterns and Size Variability. Journal of Marine Science and Engineering. 2021; 9(5):551. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9050551
Chicago/Turabian StyleKontakiotis, George, Eirini Efstathiou, Stergios D. Zarkogiannis, Evangelia Besiou, and Assimina Antonarakou. 2021. "Latitudinal Differentiation among Modern Planktonic Foraminiferal Populations of Central Mediterranean: Species–Specific Distribution Patterns and Size Variability" Journal of Marine Science and Engineering 9, no. 5: 551. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9050551