
Factors Influencing Habitat Selection of Three Cryptobenthic Clingfish Species in the Shallow North Adriatic Sea


{kind=link}
{kind=link}
{kind=link}
{kind=link}
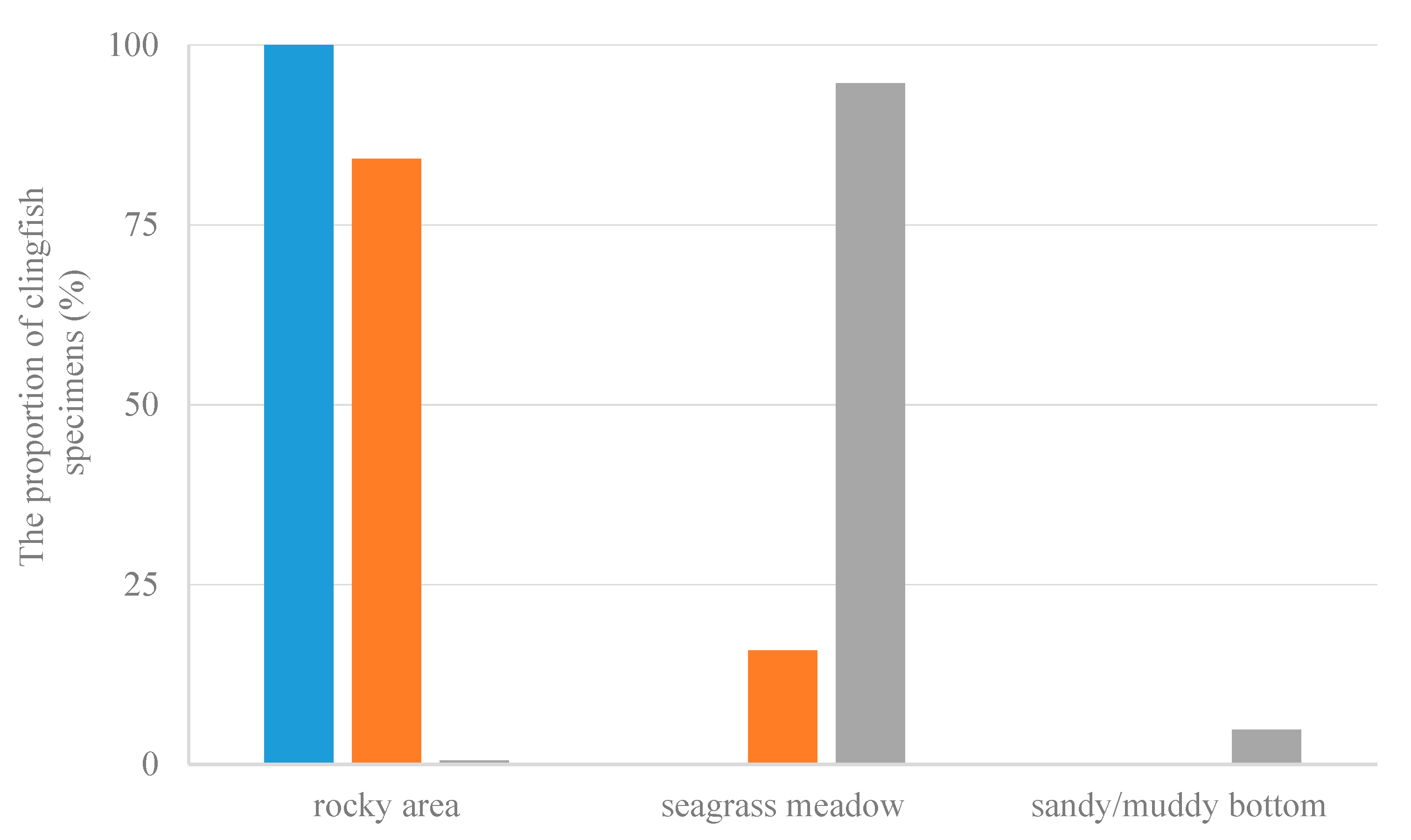{kind=link}
{kind=link}
{kind=link}
{kind=link}
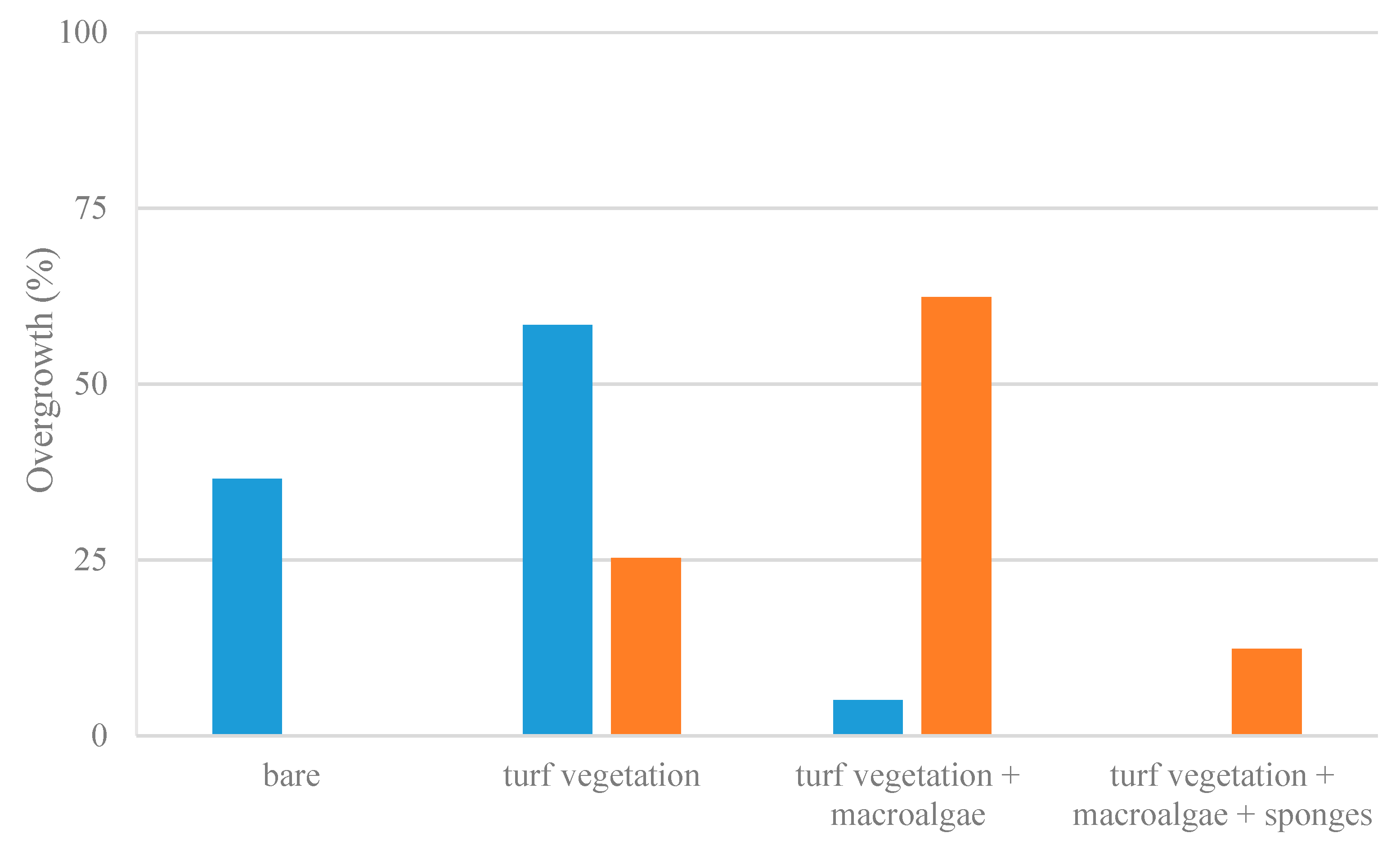{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
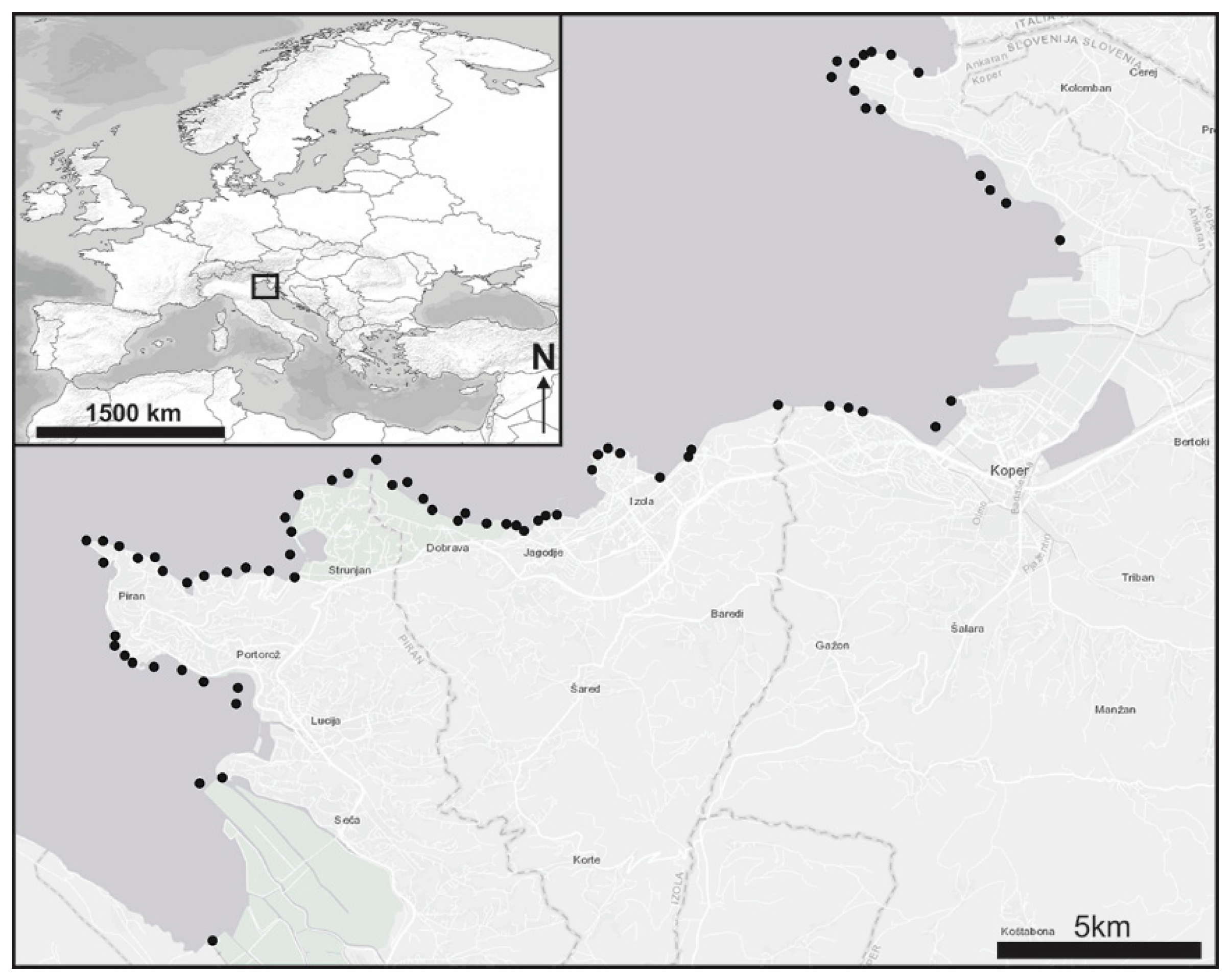
2.1. Study Area
2.2. Data Collection
2.2.1. Field Work
2.2.2. Fish Collection
2.2.3. Habitat Survey
2.2.4. Laboratory Work
2.2.5. Data Analysis
3. Results
3.1. Size Distribution and Sex Ratio of Three Species
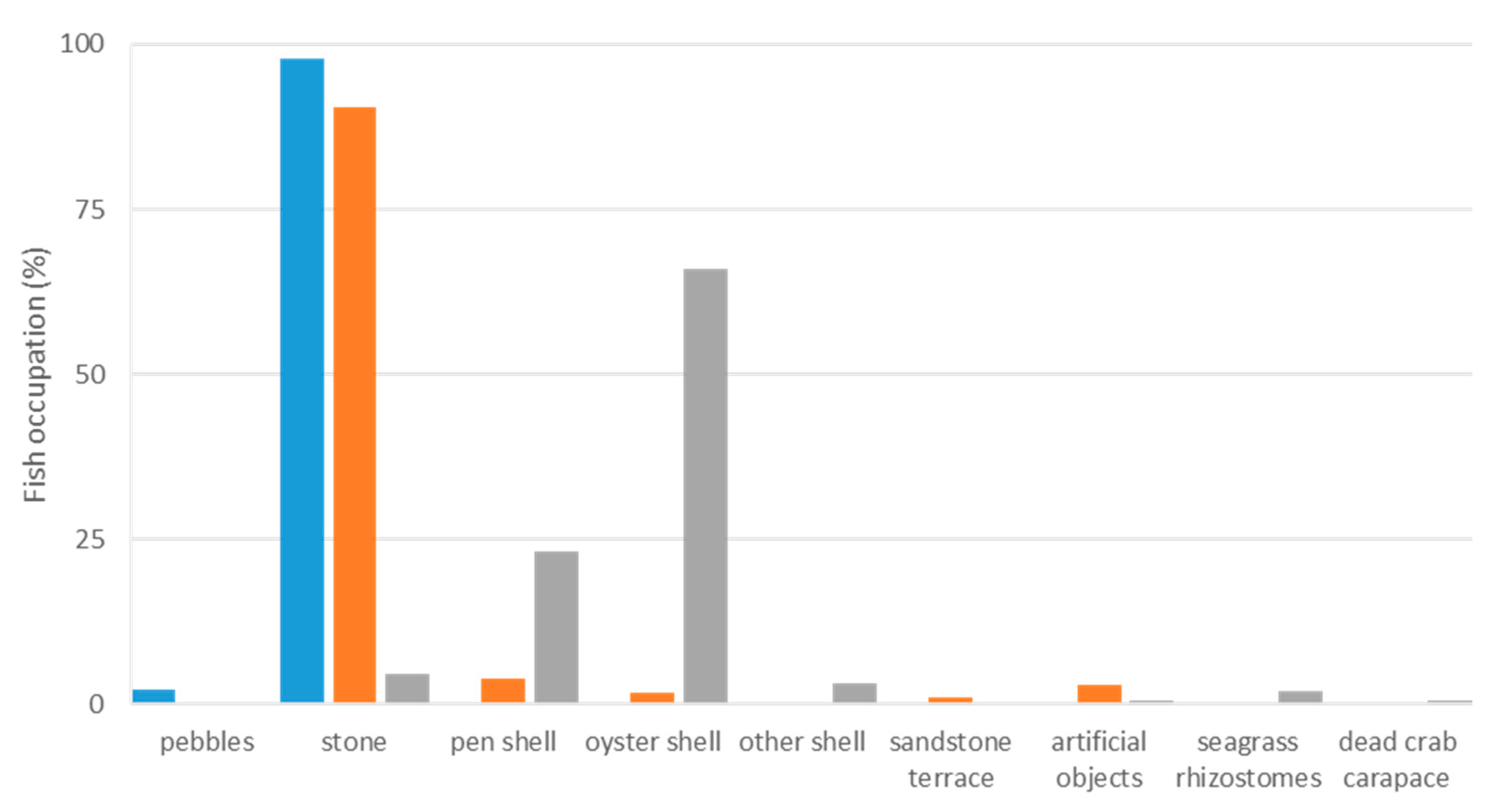
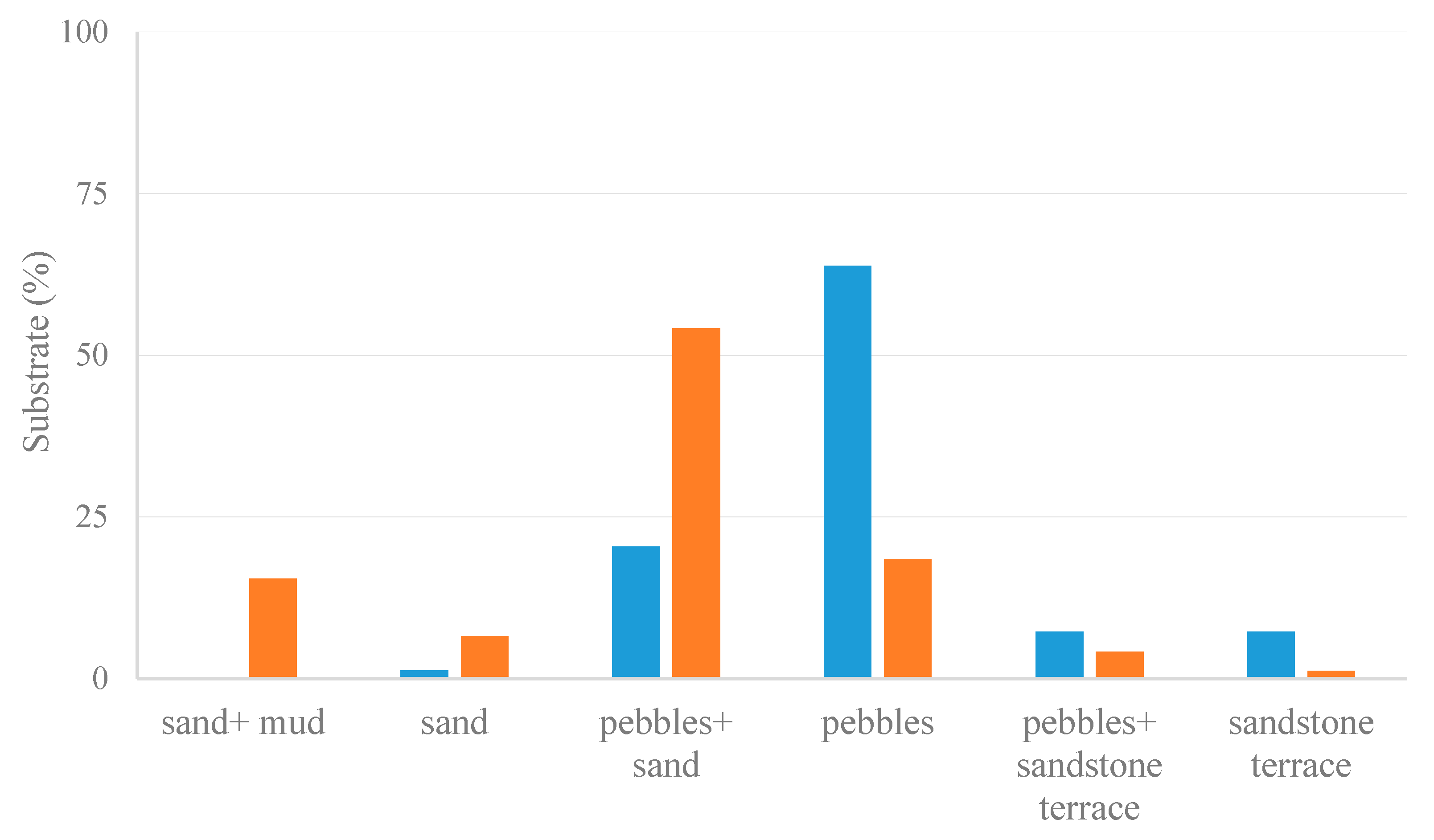
3.2. Habitat Preferences of Lepadogaster lepadogaster
3.3. Habitat Preferences of Lepadogaster candolii
3.4. Habitat Preferences of Apletodon incognitus
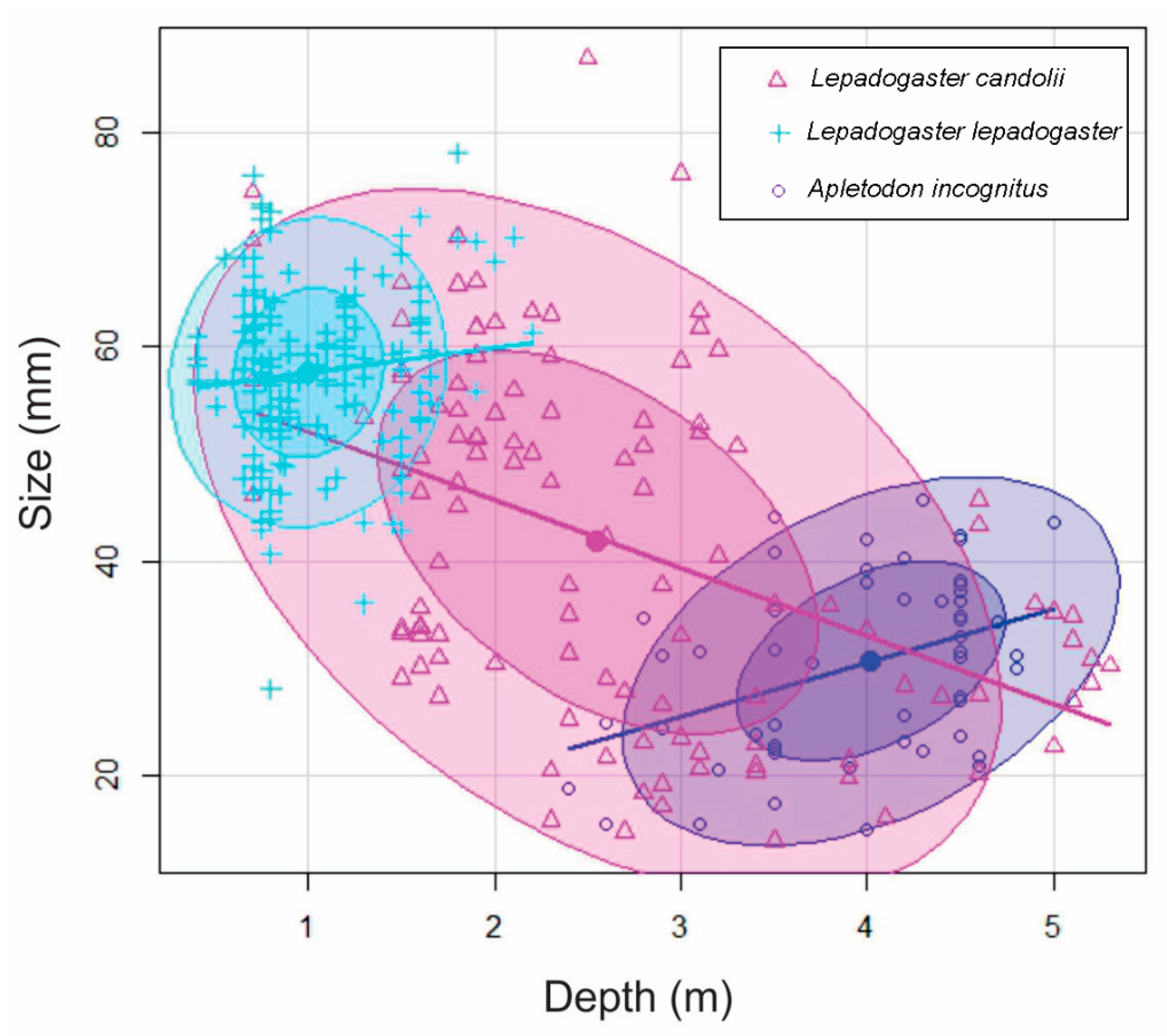
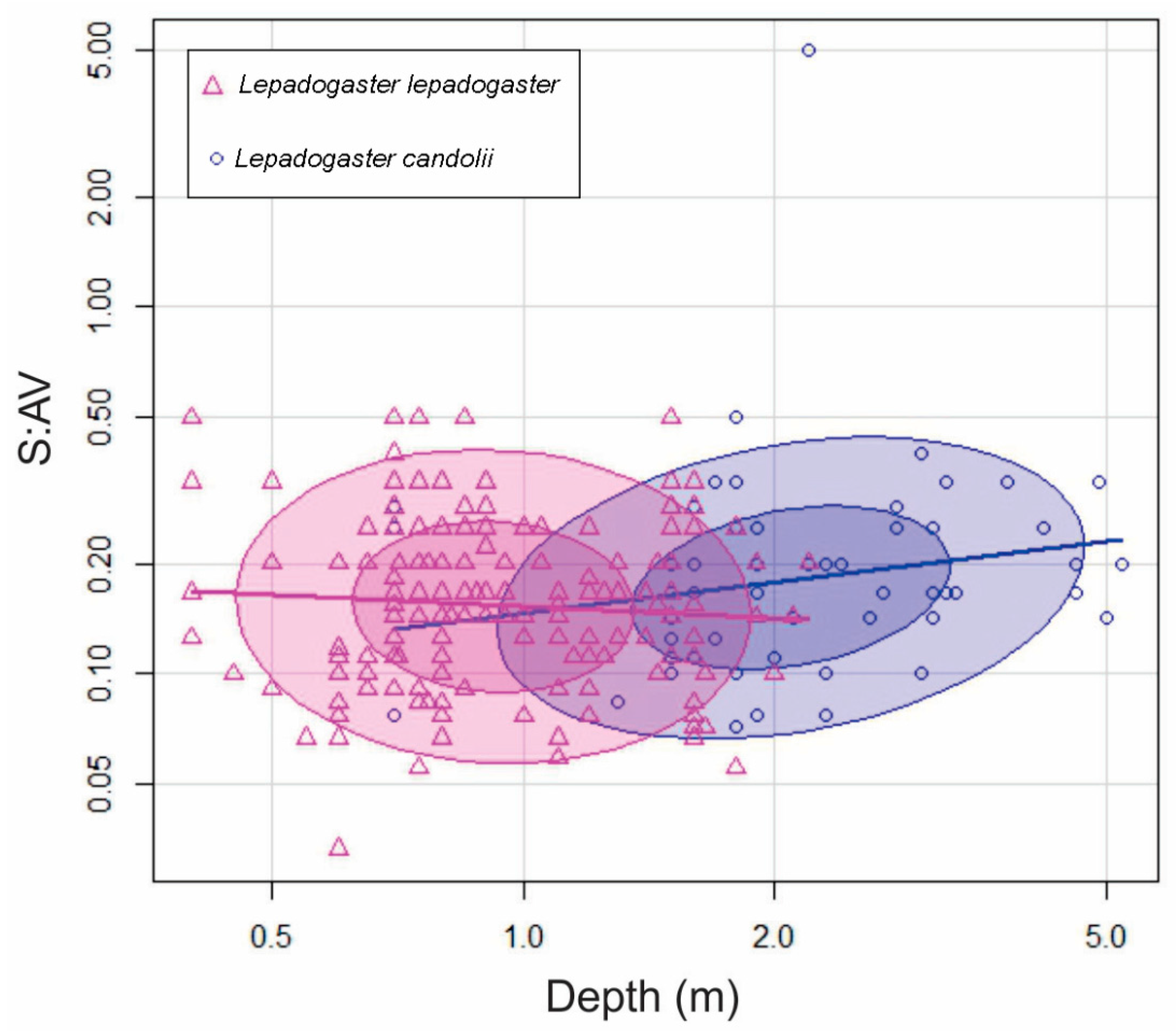
3.5. Comparison of Three Clingfish Species
3.5.1. Size Distribution and Sex Ratio
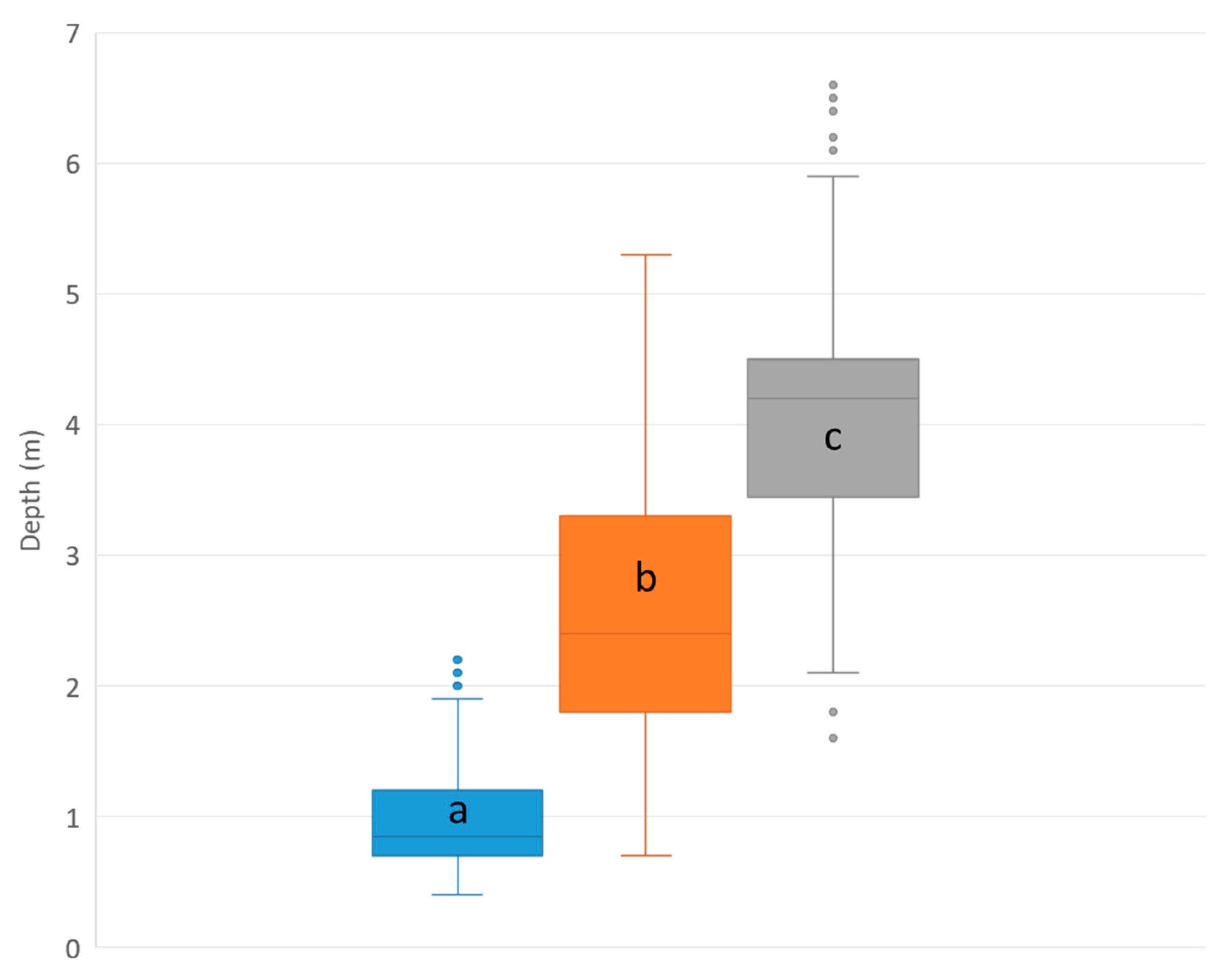
3.5.2. Depth Distribution
3.5.3. Habitat Selection
4. Discussion
4.1. Size Distribution and Sex Ratio
4.2. Habitat Selection Patterns in Three Clingfish Species
4.2.1. Species Occurrence and Abundance
4.2.2. Depth Segregation
4.2.3. Habitat Selection
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cañadas, A.; Sagarminaga, R.; De Stephanis, R.; Urquiola, E.; Hammond, P.S. Habitat Preference Modelling as a Conservation Tool: Proposals for Marine Protected Areas for Cetaceans in Southern Spanish Waters. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 495–521. [Google Scholar] [CrossRef]
- Huey, R.B. Physiological Consequences of Habitat Selection. Am. Nat. 1991, 137, S91–S115. [Google Scholar] [CrossRef] [Green Version]
- Franklin, A.B.; Anderson, D.R.; Gutiérrez, R.J.; Burnham, K.P. Climate, Habitat Quality, and Fitness in Northern Spotted Owl Populations in Northwestern California. Ecol. Monogr. 2000, 70, 539–590. [Google Scholar] [CrossRef]
- Crowe, T.P.; Underwood, A.J. Testing Behavioural “preference” for Suitable Microhabitat. J. Exp. Mar. Biol. Ecol. 1998, 225, 1–11. [Google Scholar] [CrossRef]
- Brooker, R.M.; Munday, P.L.; Mcleod, I.M.; Jones, G.P. Habitat Preferences of a Corallivorous Reef Fish: Predation Risk versus Food Quality. Coral Reefs 2013, 32, 613–622. [Google Scholar] [CrossRef]
- Mittelbach, G.G. Foraging Efficiency and Body Size: A Study of Optimal Diet and Habitat Use by Bluegills. Ecology 1981, 62, 1370–1386. [Google Scholar] [CrossRef]
- Gilliam, J.F.; Fraser, D.F. Habitat Selection Under Predation Hazard: Test of a Model with Foraging Minnows. Ecology 1987, 68, 1856–1862. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J. Adaptiveness and Implications of Small Size in Teleosts. Symp. Zool. Soc. Lond. 1979, 44, 263–306. [Google Scholar]
- Henriques, M.; Lourenco, R.; Almada, F.; Calado, G.; Goncalves, D.; Guillemaud, T.; Cancela, M.L.; Almada, V.C. A Revision of the Status of Lepadogaster lepadogaster (Teleostei: Gobiesocidae): Sympatric Subspecies or a Long Misunderstood Blend of Species? Biol. J. Linn. Soc. 2002, 76, 327–338. [Google Scholar] [CrossRef]
- Smith-Vaniz, W.F.; Jelks, H.L.; Rocha, L.A. Relevance of Crypticfishes in the Biodiversity Assessments: A Case Study at Buck Island Reef National Monument, St. Croix. Bull. Mar. Sci. 2006, 79, 17–48. [Google Scholar]
- Miller, P.J. Gobiidae. In Fishes of the North-Eastern Atlantic and the Mediterranean; Whitehead, P.J.P., Bauchot, M.-L.H., Nielsen, J.-C., Tortonese, J.E., Eds.; UNESCO: Paris, France, 1986; pp. 1019–1085. [Google Scholar]
- Patzner, R.A. Habitat Utilization and Depth Distribution of Small Cryptobenthic Fishes (Blenniidae, Gobiesocidae, Gobiidae, Tripterygiidae) in Ibiza (Western Mediterranean Sea). Environ. Biol. Fishes 1999, 55, 207–214. [Google Scholar] [CrossRef]
- Hofrichter, R.; Patzner, R.A. Habitat and Microhabitat of Mediterranean Clingfishes (Teleostei: Gobiesociformes: Gobiesocidae). Mar. Ecol. 2000, 21, 41–53. [Google Scholar] [CrossRef]
- Glavičić, I.; Paliska, D.; Soldo, A.; Kovačić, M. A Quantitative Assessment of the Cryptobenthic Fish Assemblage at Deep Littoral Cliffs in the Mediterranean. Sci. Mar. 2016, 80, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Ackerman, J.L.; Bellwood, D.R. Reef Fish Assemblages: A Re-Evaluation Using Enclosed Rotenone Stations. Mar. Ecol. Prog. Ser. 2000, 206, 227–237. [Google Scholar] [CrossRef]
- Ackerman, J.L.; Bellwood, D.R. Comparative Efficiency of Clove Oil and Rotenone for Sampling Tropical Reef Fish Assemblages. J. Fish Biol. 2002, 60, 893–901. [Google Scholar] [CrossRef]
- Depczynski, M.; Bellwood, D.R. The Role of Cryptobenthic Reef Fishes in Coral Reef Trophodynamics. Mar. Ecol. Prog. Ser. 2003, 256, 183–191. [Google Scholar] [CrossRef]
- Hofrichter, R. Taxonomie, Verbreitung und Ökologie von Schildfischen der Unterfamilie Lepadogastrinae (Gobiesocidae, Teleostei). Ph.D. Thesis, University of Salzburg, Salzburg, Austria, 1995. [Google Scholar]
- Brandl, S.J.; Wagner, M.; Hofrichter, R.; Patzner, R.A. First Record of the Clingfish Apletodon dentatus (Gobiesocidae) in the Adriatic Sea and a Description of a Simple Method to Collect Clingfishes. Bull. Fish Biol. 2011, 13, 65–69. [Google Scholar]
- Wheeler, A. The Fishes of the British Isles and North-West Europe; Macmillan: London, UK, 1969. [Google Scholar]
- Briggs, J.C. Gobiesocidae. In Check List of the North-Eastern Atlantic and of the Mediterranean; Hureau, J.C., Monod, T., Eds.; UNESCO: Paris, France, 1986; pp. 651–656. [Google Scholar]
- Briggs, J.C. Gobiesocidae. In Check List of the Fisches of the Eastern Tropical Atlantic; Metivier, C., Sanchez-Jaimes, J., Eds.; UNESCO: Lisbon, Portugal, 1990; pp. 474–478. [Google Scholar]
- Dunne, J. Gobiesocidae Occuring in the Coastal Waters of Connemara. Ir. Fish. Invest. 1983, 23, 32–36. [Google Scholar]
- Hofrichter, R. Beitrag zur Kenntnis der Mediterranen Schildfische (Teleostei, Gobiesocidae) mit Besonderer Berücksichtigung der Fortpflanzung von Lepadogaster lepadogaster. Master’s Thesis, University of Salzburg, Salzburg, Austria, 1993. [Google Scholar]
- Illich, I.P.; Kotrschal, K. Depth Distribution and Abundance of Northern Adriatic Littoral Rocky Reef Blennioid Fishes (Blenniidae and Tripterygion). Mar. Ecol. 1990, 11, 277–289. [Google Scholar] [CrossRef]
- King, P.A. Littoral and Benthic Investigations on the West Coast of Ireland: XXII, The Biology of a Population of Shore Clingfish Lepadogaster lepadogaster (Bonnaterre, 1788) at Inishbofin, Co. Galway. Proc. R. Ir. Acad. B. 1989, 89, 47–58. [Google Scholar]
- Wilson, J.P.F. A Note on the Biology of the Cornish Clingfish Lepadogaster lepadogaster (Bonnaterre). Irish Nat. J. 1981, 20, 209–210. [Google Scholar]
- Kovačić, M.; Patzner, R.A.; Schliewen, U. A First Quantitative Assessment of the Ecology of Cryptobenthic Fishes in the Mediterranean Sea. Mar. Biol. 2012, 159, 2731–2742. [Google Scholar] [CrossRef]
- Hofrichter, R.; Patzner, R.A. A New Species of Apletodon from the Mediterranean Sea and the Eastern Atlantic with Notes on the Differentiation between Apletodon and Diplecogaster Species (Pisces: Teleostei: Gobiesociformes: Gobiesocidae). Senckenb. Biol. 1997, 77, 15–22. [Google Scholar]
- Patzner, R.A.; Santo, R.S. Field Observations on the Association Between the Clingfish Diplecogaster bimaculata pectoralis Briggs, 1955 and Different Species of Sea Urchins at the Azores. Z. Fischk. 1992, 1, 157–161. [Google Scholar]
- Bilecenoglu, M.; Kaya, M. The Occurrence of Apletodon incognitus Hofrichter & Patzner, 1997 (Gobiesocidae) in the Eastern Mediterranean Sea. Acta Ichthyol. Piscat. 2006, 36, 143–145. [Google Scholar]
- Louisy, P. Europe and Mediterranean Marine Fish Identification Guide; Les Éditions Eugen Ulmer: Paris, France, 2015. [Google Scholar]
- Ogorelec, B.; Mišič, M.; Faganeli, J. Marine Geology of the Gulf of Trieste (Northern Adriatic): Sedimentological Aspects. Mar. Geol. 1991, 99, 79–92. [Google Scholar] [CrossRef]
- Lipej, L.; Ivajnšič, D.; Makovec, T.; Mavrič, B.; Šiško, M.; Trkov, D.; Orlando-Bonaca, M. Raziskava z Oceno Stanja Morskih Travnikov v Krajinskem Parku Strunjan; Marine Biology Station Piran, National Institute of Biology: Piran, Slovenia, 2018. [Google Scholar]
- Gibson, R.N. The Use of the Anaesthetic Quinaldine in Fish Ecology. J. Anim. Ecol. 1967, 36, 301. [Google Scholar] [CrossRef]
- Wagner, M.; Bračun, S.; Skofitsch, G.; Kovačić, M.; Zogaris, S.; Iglésias, S.P.; Sefc, K.M.; Koblmüller, S. Diversification in Gravel Beaches: A Radiation of Interstitial Clingfish (Gouania, Gobiesocidae) in the Mediterranean Sea. Mol. Phylogenet. Evol. 2019, 139, 106525. [Google Scholar] [CrossRef] [PubMed]
- Ličer, M.; Fettich, A.; Jeromel, M. Prognozirano Plimovanje Morja Tide Tables 2018; Ministry of the Environment and Spatial Planning, Slovenian Environment Agency: Ljubljana, Slovenia, 2018.
- Ličer, M.; Fettich, A.; Jeromel, M. Prognozirano Plimovanje Morja Tide Tables 2019; Ministry of the Environment and Spatial Planning, Slovenian Environment Agency: Ljubljana, Slovenia, 2019.
- Ličer, M.; Fettich, A.; Jeromel, M. Prognozirano Plimovanje Morja Tide Tables 2017; Ministry of the Environment and Spatial Planning, Slovenian Environment Agency: Ljubljana, Slovenia, 2017.
- Jardas, I. Jadranska Ihtiofauna; Školska knjiga: Zagreb, Croatia, 1996. [Google Scholar]
- Marčeta, B. Osteichthyes. In Key for Determination of Vertebrates in SLOVENIA; Kryštufek, B., Janžekovič, F., Eds.; DZS: Ljubljana, Slovenia, 1999; pp. 47–210. [Google Scholar]
- Wagner, M.; Bračun, S.; Kovačić, M.; Iglésias, S.P.; Sellos, D.Y.; Zogaris, S.; Koblmüller, S. Lepadogaster purpurea (Actinopterygii: Gobiesociformes: Gobiesocidae) from the Eastern Mediterranean Sea: Significantly Extended Distribution Range. Acta Ichthyol. Piscat. 2017, 47, 417–421. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org (accessed on 10 April 2021).
- Abel, E.F. Freiwasserbeobachtungen an Fischen Im Golf von Neapel als Beitrag zur Kenntnis Ihrer Ökologie und Ihres Verhaltens. Int. Rev. Der Gesamten Hydrobiol. Und Hydrogr. 1962, 47, 219–290. [Google Scholar] [CrossRef]
- Gibson, R.N. The Biology and Behaviour of Littoral Fish. Ocean. Mar. Biol. Ann. Rev. 1969, 7, 367–410. [Google Scholar]
- Orlando-Bonaca, M.; Lipej, L. Utilization of Empty Holes by Two Adriatic Endolithic Blennies under Experimental Conditions—Preliminary Results. Ann. Ser. Hist. Nat. 2008, 18, 179–190. [Google Scholar]
- Trkov, D. Znotrajvrstni in Medvrstni Odnosi...Mediolitoralnih Babic (Blenniidae) Tržaškega Zaliva. Master’s Thesis, Univerza na Primorskem, Fakulteta za Matematiko, Naravoslovje in Informacijske Tehnologije, Koper, Slovenia, 2014. [Google Scholar]
- Lengkeek, W. Dynamic Reproductive Behaviour and the Evolution of Sex Size Dimorphism in Mediterranean Blennies. Ph.D. Thesis, University of East Anglia, Norwich, England, 2006. [Google Scholar]
- Zander, C.D.; Berg, J. Feeding Ecology of Littoral Gobiid and Blennioid Fishes of the Banyuls Area (Mediterranean Sea). II: Prey Selection and Size Preference. Vie Milieu 1984, 34, 149–157. [Google Scholar]
- Gibson, R.N. The Vertical Distribution and Feeding Relationships of Intertidal Fish on the Atlantic Coast of France. J. Anim. Ecol. 1972, 41, 189–207. [Google Scholar] [CrossRef]
- Brandstätter, R.; Körbek, R. Biologie Der Saugfische: 1. Habitat, Verbreitung und Nahrung. BUFUS Info. Zool. Salzbg. 1989, 5, 27–29. [Google Scholar]
- Guttridge, T.L.; Gruber, S.H.; Franks, B.R.; Kessel, S.T.; Gledhill, K.S.; Uphill, J.; Krause, J.; Sims, D.W. Deep Danger: Intra-Specific Predation Risk Influences Habitat Use and Aggregation Formation of Juvenile Lemon Sharks Negaprion brevirostris. Mar. Ecol. Prog. Ser. 2012, 445, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Freeman, M.C.; Stouder, D.J. Intraspecific Interactions Influence Size Specific Depth Distribution in Cottus bairdi. Environ. Biol. Fishes 1989, 24, 231–236. [Google Scholar] [CrossRef]
- Johnson, C.R. Notes on the Intertidal Life History of the Northern Clingfish, Gobiesox maeandricus (Girard). Am. Midl. Nat. 1970, 83, 625. [Google Scholar] [CrossRef]
- Pires, T.H.S.; Gibran, F.Z. Intertidal Life: Field Observations on the Clingfish Gobiesox barbatulus in Southeastern Brazil. Neotrop. Ichthyol. 2011, 9, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Trkov, D.; Lipej, L. A Non-Destructive Method for Assessing the Feeding Habits of Coastal Fish. Mediterr. Mar. Sci. 2019, 20, 453–459. [Google Scholar] [CrossRef]
- Holthuis, L.B. Report on a Collection of Crustacea Decapoda and Stomatopoda from Turkey and the Balkans. Zool. Verh. Leiden. 1961, 47, 1–67. [Google Scholar]
- Melzer, R.; Ceseña, F.; Buršić, M.; Lehmann, T.; Mayer, R.; Mavrič, B.; Makovec, T.; Pfannkuchen, M.; McHenry, J.; Heß, M. Knights, Ballerinas and Invisibles: The Decapod Crustaceans of the Brijuni Marine Protected Area = Vitezovi, Balerine i Nevidljivi: Rakovi Deseteronošci Zaštićenog Morskog Područja Nacionalnog Parka Brijuni; Javna ustanova Nacionalni park Brijuni: Pula, Croatia, 2019. [Google Scholar]
- Gonclalves, E.J.; Almada, V.C.; Almeida, S.P.; Gonclalves, D.M.; Repas, M.; Simoes, N. Observations on the Agonistic Behaviour of Lepadogaster lepadogaster purpurea (Pisces: Gobiesocidae). J. Fish Biol. 1996, 49, 367–369. [Google Scholar] [CrossRef]
- Paine, R.T.; Palmer, A.R. Syciases Sanguineus: A Unique Trophic Generalist from the Chilean Intertidal Zone. Copeia 1978, 1, 75–81. [Google Scholar] [CrossRef]
- Gonçalves, D.M.; Gonçalves, E.J.; Almada, V.C.; Almeida, S.P. Comparative Behaviour of Two Species of Lepadogaster (Pisces: Gobiesocidae) Living at Different Depth. J. Fish Biol. 1998, 53, 447–450. [Google Scholar] [CrossRef]
- Kovačić, M. Cryptobenthic Gobies (Pisces, Perciformes, Gobiidae) and Clingfishes (Pisces, Gobiesociformes, Gobiesocidae) in the Kvarner Area, Adriatic Sea. Nat. Croat. 1997, 6, 423–435. [Google Scholar]
- Pallas, A.; Garcia-Calvo, B.; Corgos, A.; Bernardez, C.; Freire, J. Distribution and Habitat Use Patterns of Benthic Decapod Crustaceans in Shallow Waters: A Comparative Approach. Mar. Ecol. Prog. Ser. 2006, 324, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Almada, V.C.; Santos, R.S. Parental Care in the Rocky Intertidal: A Case Study of Adaptation and Exaptation in Mediterranean and Atlantic Blennies. Rev. Fish Biol. Fish. 1995, 5, 23–37. [Google Scholar] [CrossRef]
- Velasco, E.M.; Gómez-Cama, M.C.; Hernando, J.A.; Soriguer, M.C. Trophic Relationships in an Intertidal Rockpool Fish Assemblage in the Gulf of Cádiz (NE Atlantic). J. Mar. Syst. 2010, 80, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Mazé, R.A. Estudio de La Dieta de Las Poblaciones Intermareales de Dos Especies de Lepadogaster (Teleostei, Gobiesocidae) En La Costa Cantábrica, España. Boletín La Real Soc. Española Hist. Nat. Sección Biológica. 2007, 102, 85–92. [Google Scholar]
- Koppel, V.H. Habitat Selection and Space Partitioning among Two Mediterranean Blenniid Species. Mar. Ecol. 1988, 9, 329–346. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M. Izbira Mikrohabitatov pri Obrežni Favni Babic v Tržaškem Zalivu. Ph.D. Thesis, Univerza v Mariboru, Maribor, Slovenia, 2006. [Google Scholar]
- Kotrschal, K.; Goldschmid, A. Morphological Evidence for the Biological Role of Caniniform Teeth in Combtooth Blennies (Blenniidae, Teleostei). J. Fish Biol. 1992, 41, 983–991. [Google Scholar] [CrossRef]
- Mora, C. Ecology of Fishes on Coral Reefs; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Fahrig, L. Relative Effects of Habitat Loss and Fragmentation on Population Extinction. J. Wildl. Manag. 1997, 61, 610. [Google Scholar] [CrossRef]
- Brandl, S.J.; Goatley, C.H.R.; Bellwood, D.R.; Tornabene, L. The Hidden Half: Ecology and Evolution of Cryptobenthic Fishes on Coral Reefs. Biol. Rev. 2018, 93, 1846–1873. [Google Scholar] [CrossRef]
- Taylor, M.S.; Van Tassell, J.L. Observations on Microhabitat Utilization by Three Widely Distributed Neotropical Gobies of the Genus Elacatinus. Copeia 2002, 2002, 1134–1136. [Google Scholar] [CrossRef]
- Luiz, O.J.; Allen, A.P.; Robertson, D.R.; Floeter, S.R.; Kulbicki, M.; Vigliola, L.; Becheler, R.; Madin, J.S. Adult and Larval Traits as Determinants of Geographic Range Size among Tropical Reef Fishes. Proc. Natl. Acad. Sci. USA 2013, 110, 16498–16502. [Google Scholar] [CrossRef] [Green Version]
- Turk, R. An Assessment of the Vulnerability of the Slovene Coastal Belt and Its Categorisation in View of (in) Admissible Human Pressure, Various Activities, and Land Use. Ann. Ser. Hist. Nat. 1999, 15, 37–50. [Google Scholar]
- Pergent, G.; Bazairi, H.; Bianchi, C.N.; Boudouresque, C.F.; Buia, M.C.; Calvo, S.; Clabaut, P.; Harmelin-Vivien, M.; Angel Mateo, M.; Montefalcone, M.; et al. Climate Change and Mediterranean Seagrass Meadows: A Synopsis for Environmental Managers. Mediterr. Mar. Sci. 2014, 15, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Telesca, L.; Belluscio, A.; Criscoli, A.; Ardizzone, G.; Apostolaki, E.T.; Fraschetti, S.; Gristina, M.; Knittweis, L.; Martin, C.S.; Pergent, G.; et al. Seagrass Meadows (Posidonia oceanica) Distribution and Trajectories of Change. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Cabanellas-Reboredo, M.; Vázquez-Luis, M.; Mourre, B.; Álvarez, E.; Deudero, S.; Amores, Á.; Addis, P.; Ballesteros, E.; Barrajón, A.; Coppa, S.; et al. Tracking a Mass Mortality Outbreak of Pen Shell Pinna nobilis Populations: A Collaborative Effort of Scientists and Citizens. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersting, D.; Mouloud, B.; Cizmek, H.; Grau, A.; Jimenez, C.; Katsanevakis, S.; Oztürk, B.; Tuncer, S.; Tunesi, L.; Vázquez-Luis, M.; et al. Pinna Nobilis. IUCN Red List Threat. Species 2019; IUCN Red List: London, UK, 2019. [Google Scholar] [CrossRef]
- Lipej, L.; Mavrič, B.; Orlando-Bonaca, M. Analiza Kriptobentoških Mikrohabitatov V Slovenskem Morju in Opredelitev Njihove Vloge Pri Ocenjevanju Stanja Biotske Raznovrstnosti Morskega Obrežnega Pasu; Marine Biology Station Piran, National Institute of Biology: Piran, Slovenia, 2012. [Google Scholar]
- Giacobbe, S. Epibiontic Mollusc Communities on Pinna nobilis L. (Bivalvia, Mollusca). J. Nat. Hist. 2002, 36, 1385–1396. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trkov, D.; Ivajnšič, D.; Kovačić, M.; Lipej, L. Factors Influencing Habitat Selection of Three Cryptobenthic Clingfish Species in the Shallow North Adriatic Sea. J. Mar. Sci. Eng. 2021, 9, 789. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080789
Trkov D, Ivajnšič D, Kovačić M, Lipej L. Factors Influencing Habitat Selection of Three Cryptobenthic Clingfish Species in the Shallow North Adriatic Sea. Journal of Marine Science and Engineering. 2021; 9(8):789. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080789
Chicago/Turabian StyleTrkov, Domen, Danijel Ivajnšič, Marcelo Kovačić, and Lovrenc Lipej. 2021. "Factors Influencing Habitat Selection of Three Cryptobenthic Clingfish Species in the Shallow North Adriatic Sea" Journal of Marine Science and Engineering 9, no. 8: 789. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080789