
Effect of Summer Typhoon Linfa on the Chlorophyll-a Concentration in the Continental Shelf Region of Northern South China Sea


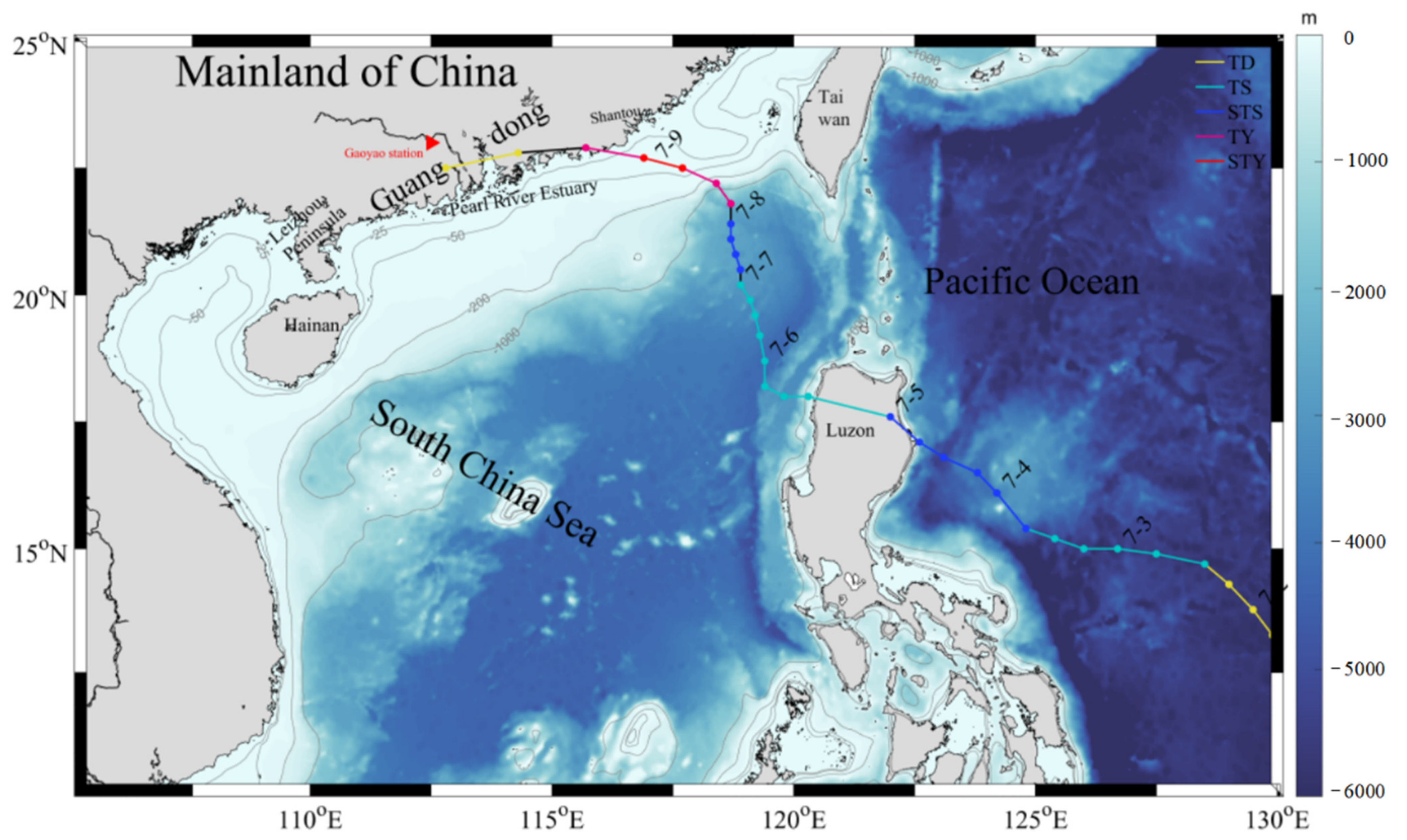{kind=link}
{kind=link}
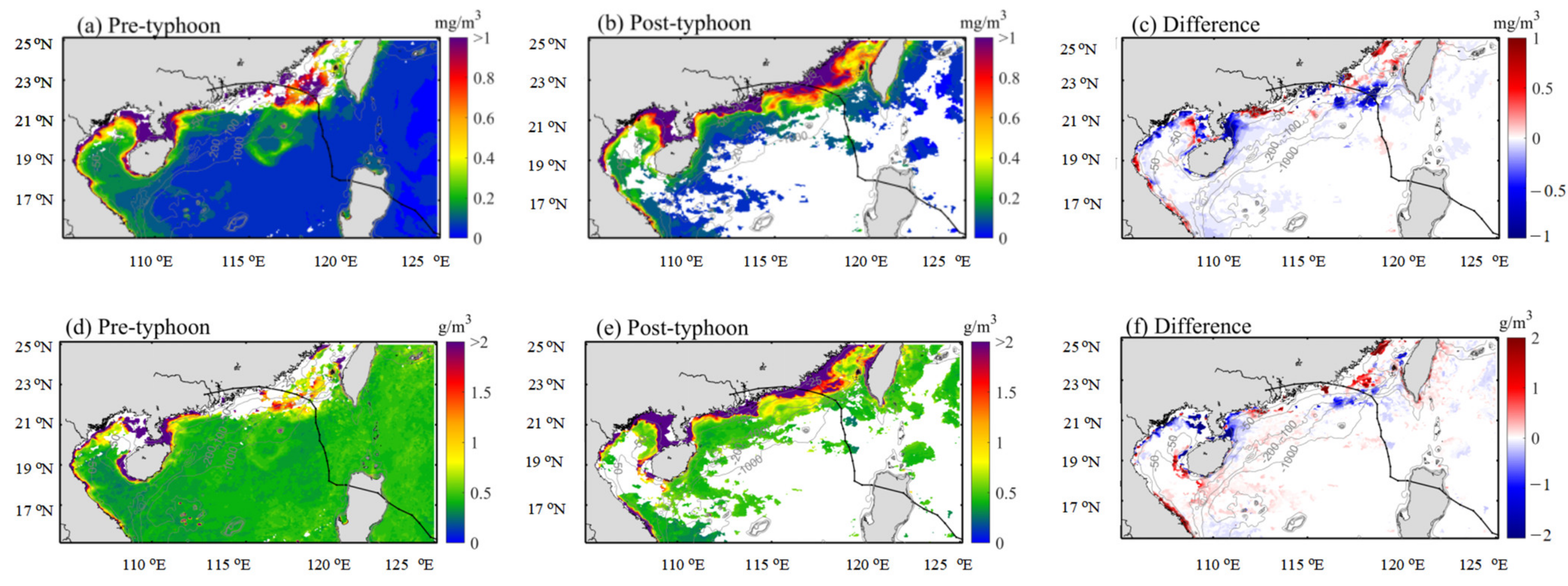{kind=link}
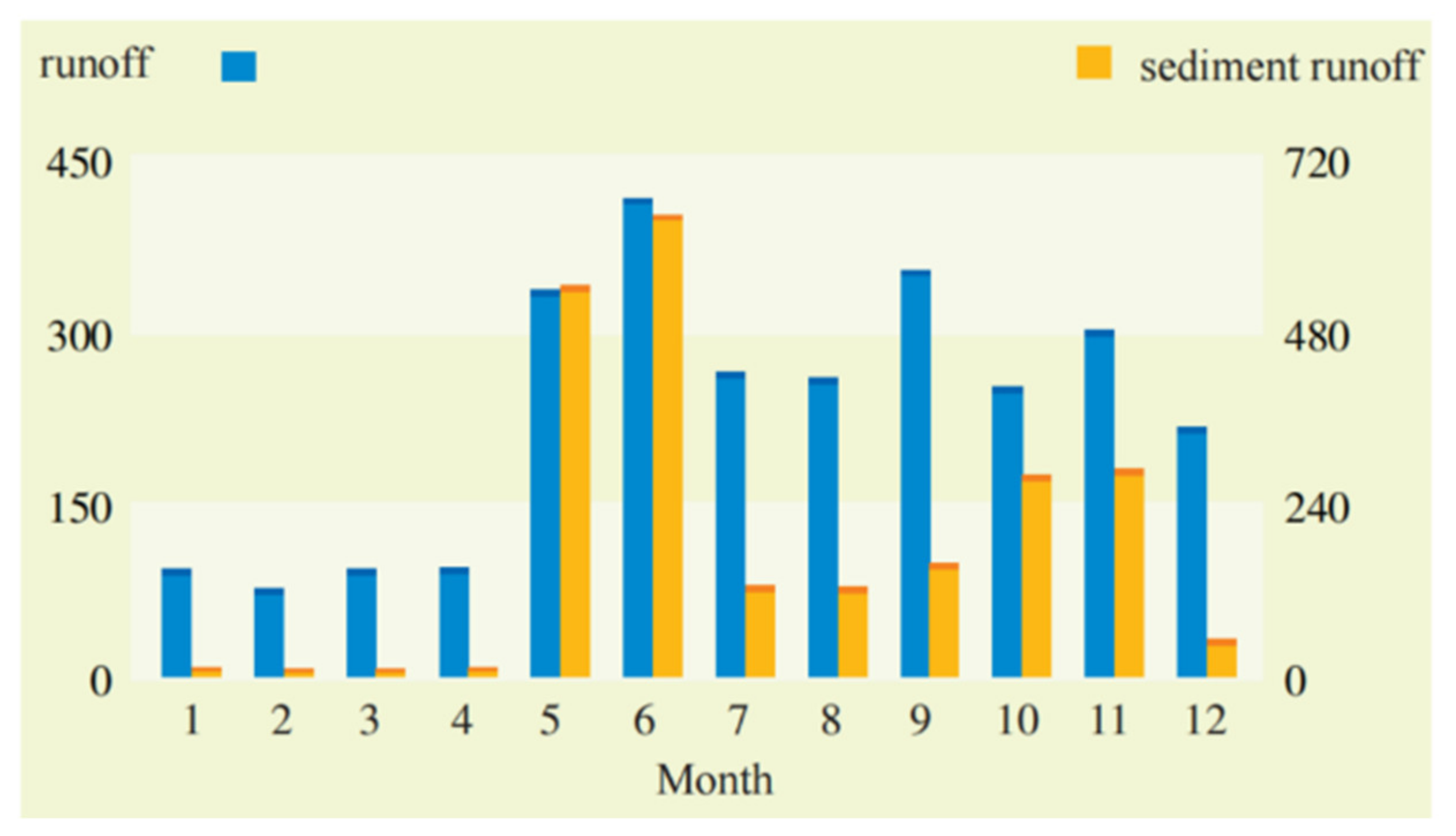{kind=link}
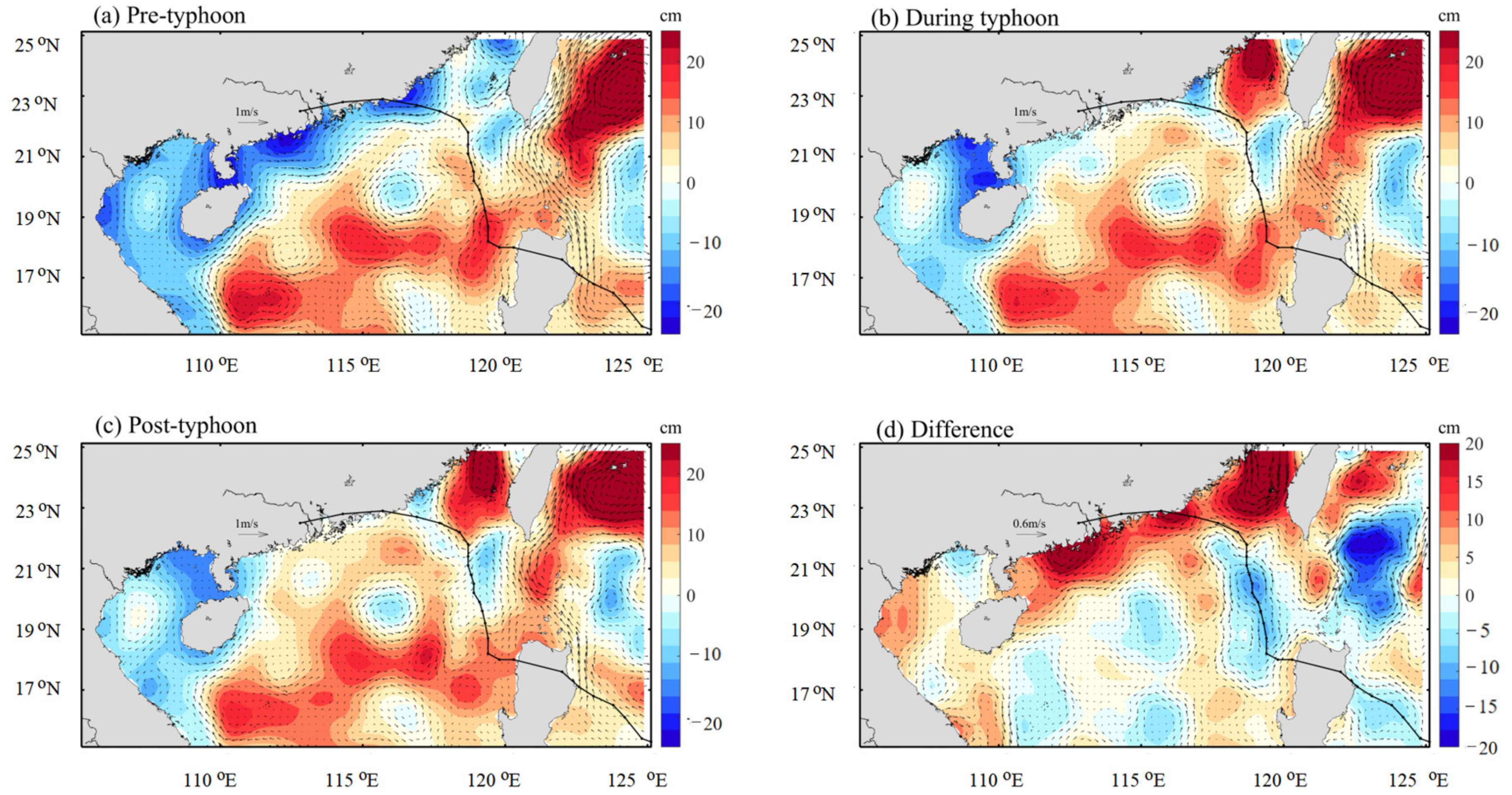{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data
2.2. Method
3. Results
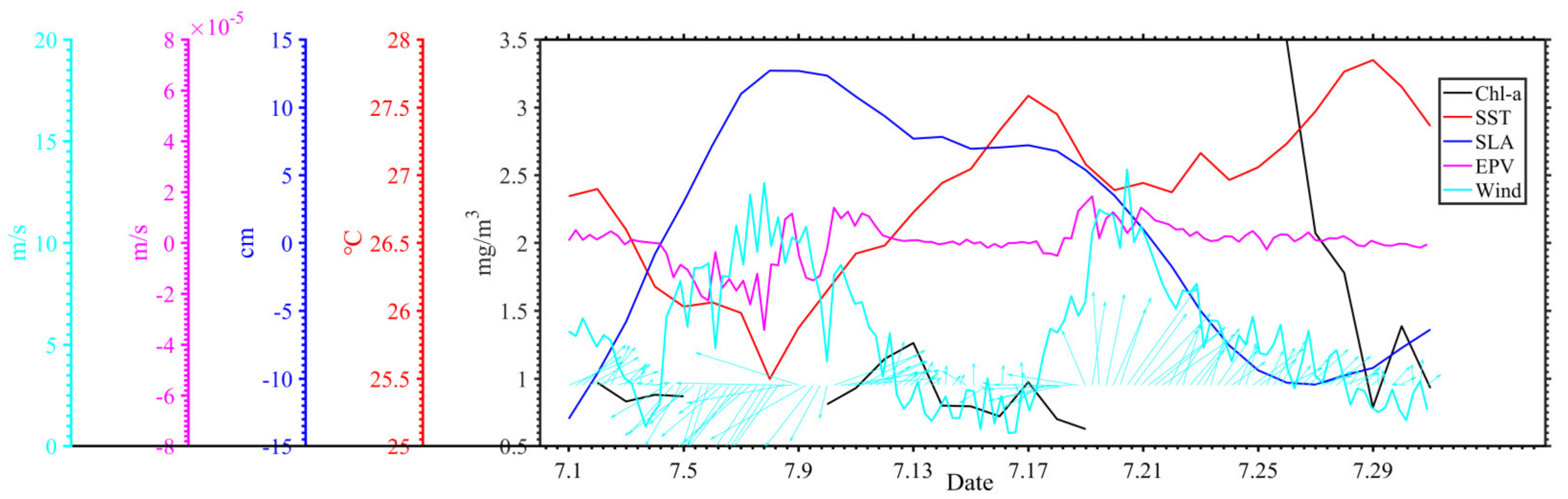
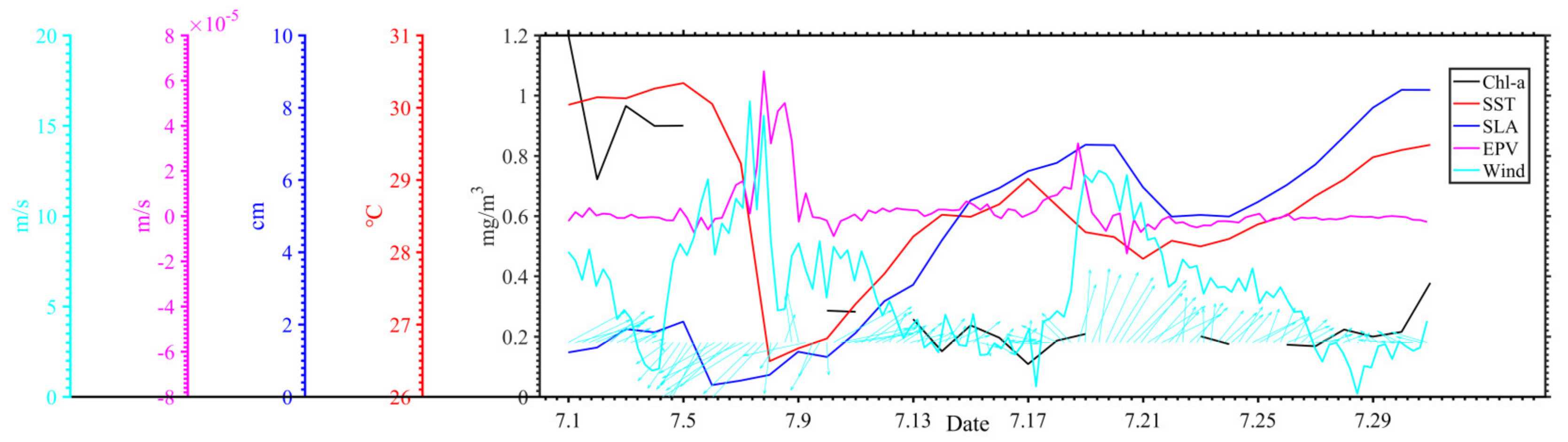
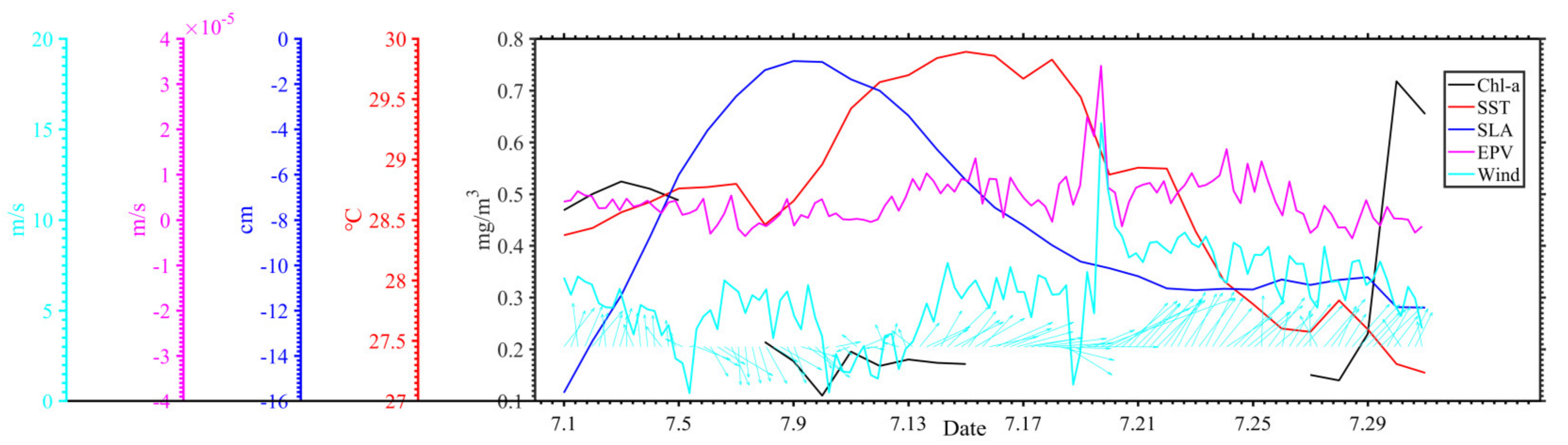
3.1. Distribution of Chlorophyll-a and Surface Suspended Sediment Concentration
3.2. Distribution of Geostrophic Current and SLA
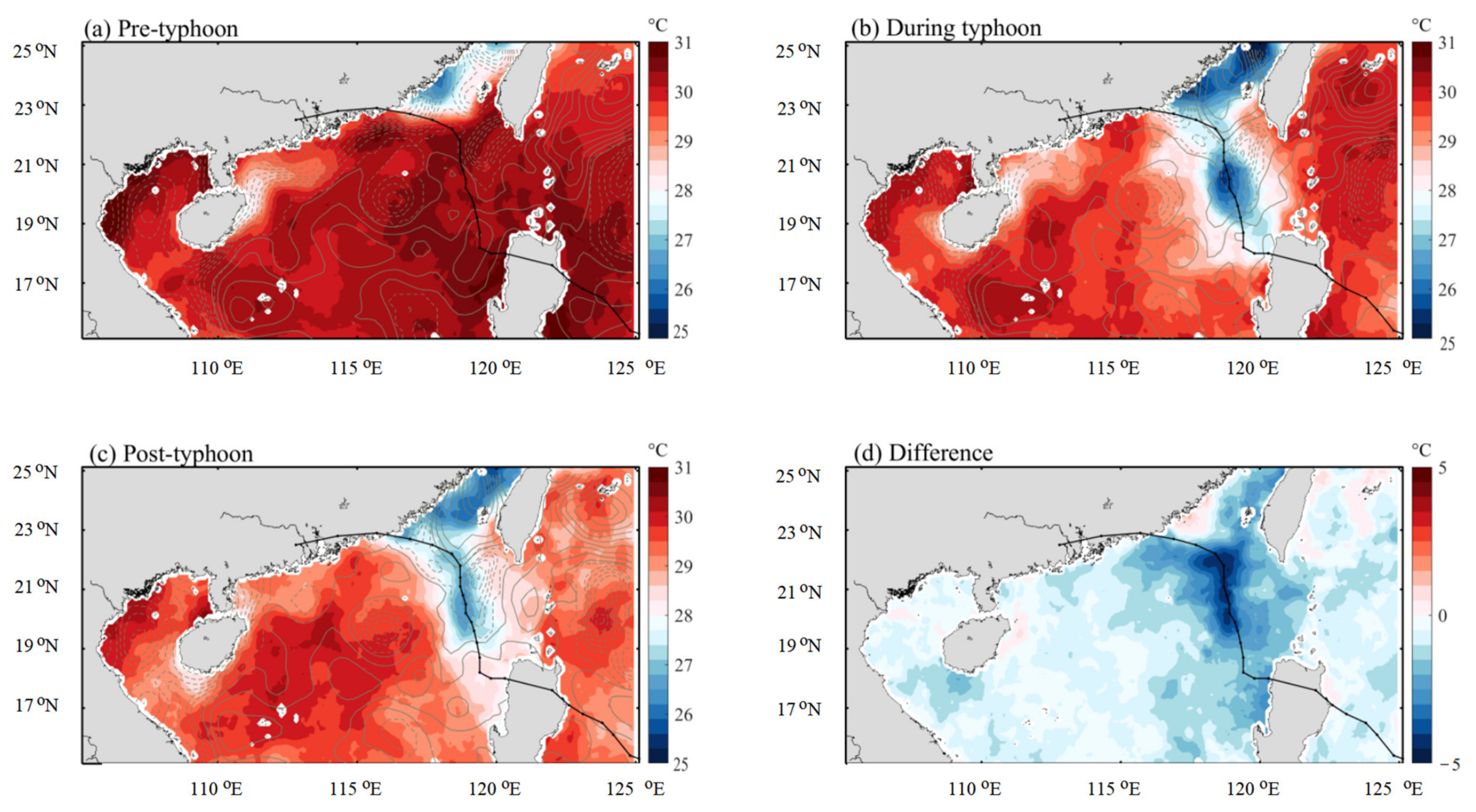
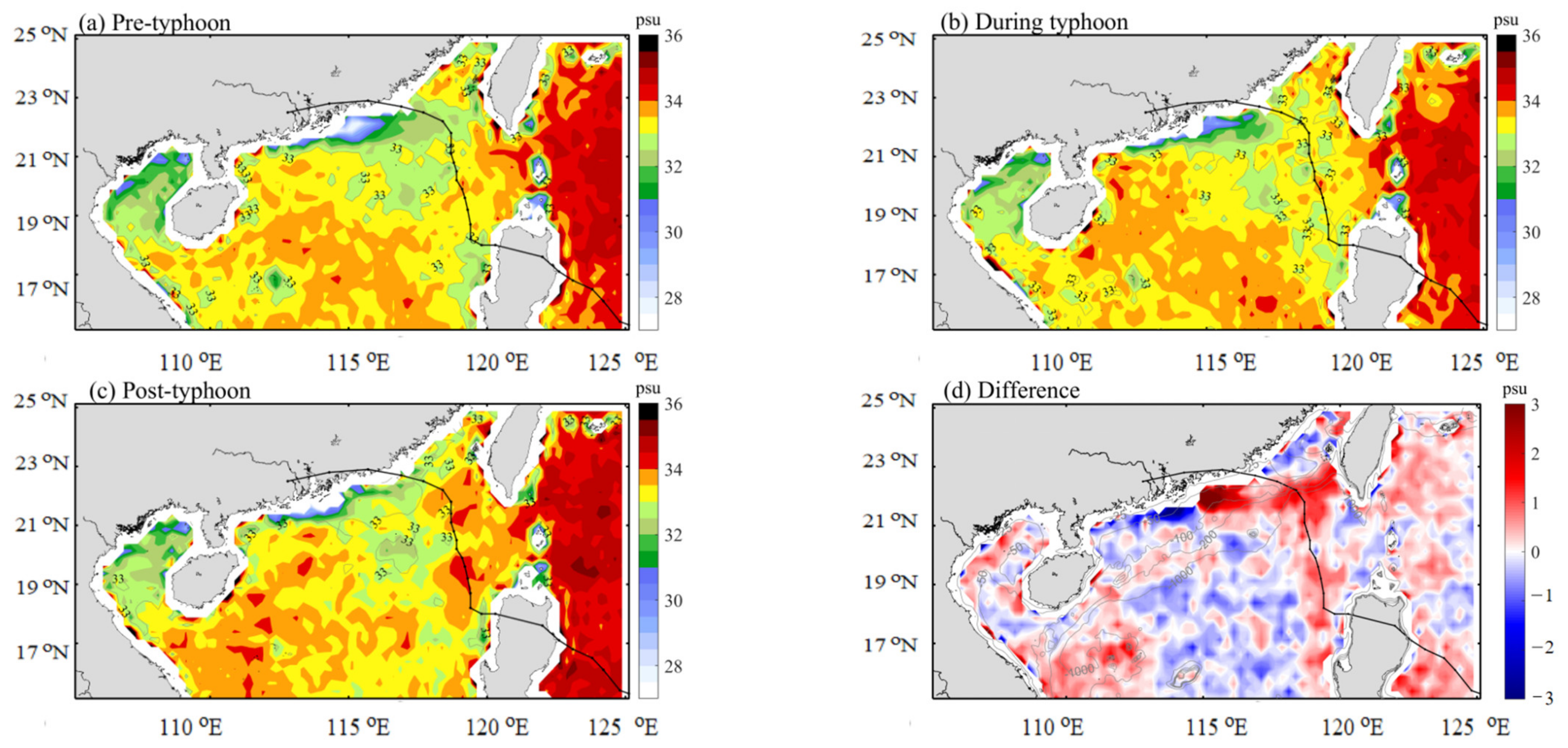
3.3. Distribution of SST and SSS
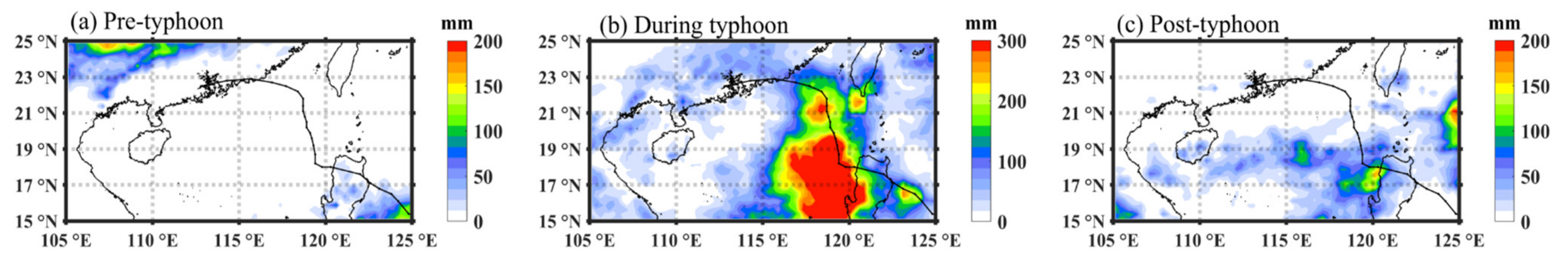
3.4. Distribution of Wind and Rainfall
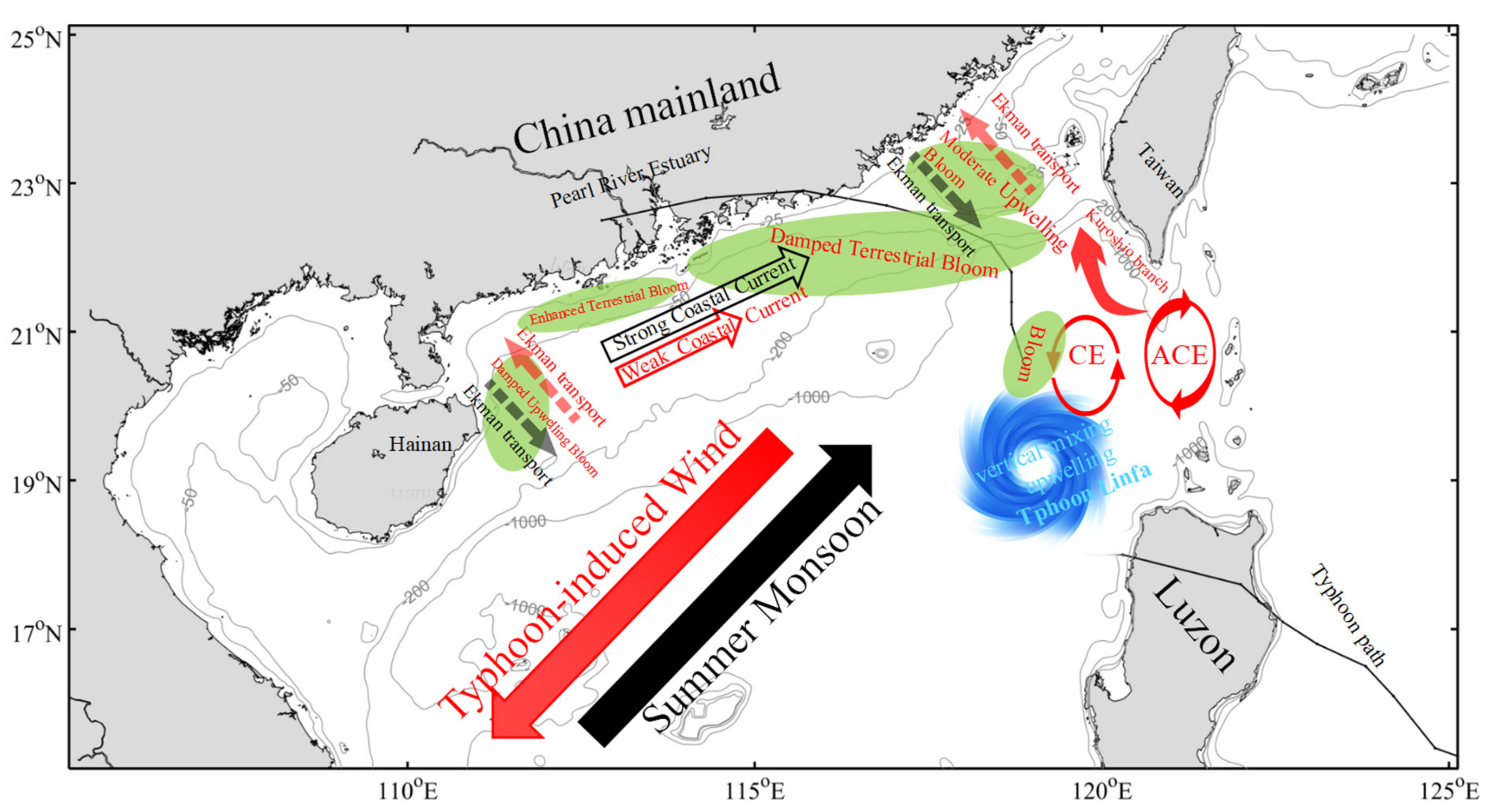
4. Discussion
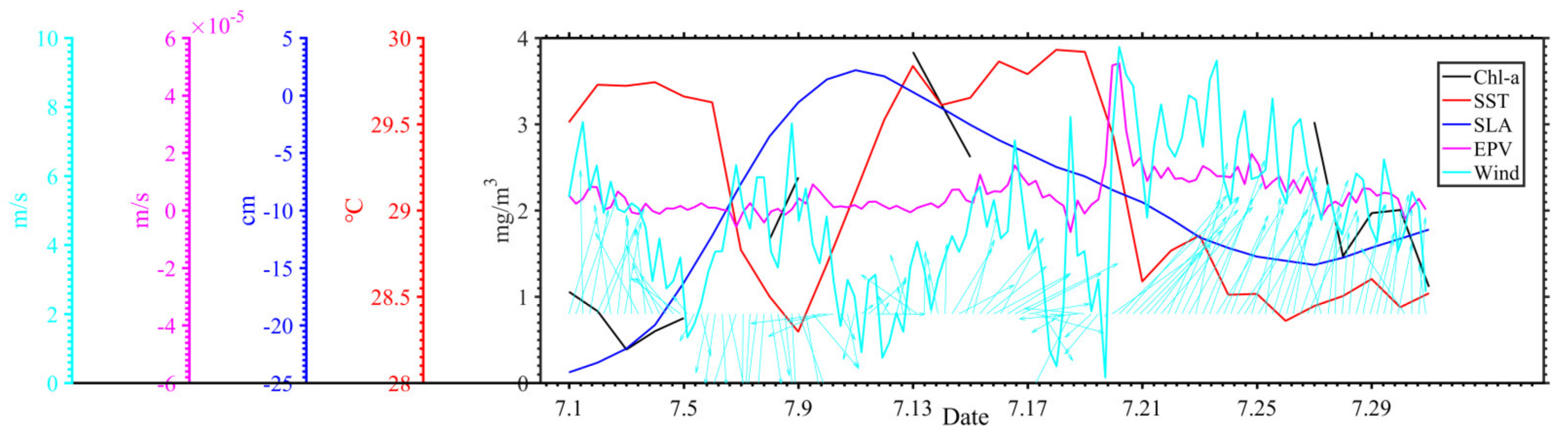
4.1. Typhoon Induced-Upwelling and Mixing
4.2. Phytoplankton Bloom in Two Regions
4.3. Phytoplankton Decline in the Two Regions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, Y.; Sun, J.; Wang, Y.; Li, S.; Xu, T.; Wei, Z.; Qu, T. Overview of the multi-layer circulation in the South China Sea. Prog. Oceanogr. 2019, 175, 171–182. [Google Scholar] [CrossRef]
- Wong, G.T.; Pan, X.; Li, K.Y.; Shiah, F.K.; Ho, T.Y.; Guo, X. Hydrography and nutrient dynamics in the northern South China sea Shelf-sea (NoSoCS). Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 117, 23–40. [Google Scholar] [CrossRef]
- Han, A.; Dai, M.; Kao, S.J.; Gan, J.; Li, Q.; Wang, L.; Zhai, W.; Wang, L. Nutrient dynamics and biological consumption in a large continental shelf system under the influence of both a river plume and coastal upwelling. Limnol. Oceanogr. 2012, 57, 486–502. [Google Scholar] [CrossRef] [Green Version]
- Gan, J.; Lu, Z.; Dai, M.; Cheung, A.Y.; Liu, H.; Harrison, P. Biological response to intensified upwelling and to a river plume in the northeastern South China Sea: A modeling study. J. Geophys. Res. Oceans. 2010, 115, C09001. [Google Scholar] [CrossRef]
- Lu, Z.; Gan, J. Controls of seasonal variability of phytoplankton blooms in the Pearl River Estuary. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 117, 86–96. [Google Scholar] [CrossRef]
- Huang, X.P.; Huang, L.M.; Yue, W.Z. The characteristics of nutrients and eutrophication in the Pearl River estuary, South China. Mar. Pollut. Bull. 2003, 47, 30–36. [Google Scholar] [CrossRef]
- Zhai, W.; Dai, M.; Cai, W.-J.; Wang, Y.; Wang, Z. High partial pressure of CO2 and its maintaining mechanism in a subtropical estuary: The Pearl River estuary, China. Mar. Chem. 2005, 93, 21–32. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, X.X.; Higgitt, D.L.; Chen, C.T.A.; Han, J.; Sun, H. Recent changes of water discharge and sediment load in the Zhujiang (Pearl River) Basin, China. Glob. Environ. Chang. 2008, 60, 365–380. [Google Scholar] [CrossRef]
- Shu, Y.; Wang, D.; Zhu, J.; Peng, S. The 4-D structure of upwelling and Pearl River plume in the northern South China Sea during summer 2008 revealed by a data assimilation model. Ocean Modell. 2011, 36, 228–241. [Google Scholar] [CrossRef]
- Wang, D.; Zhuang, W.; Xie, S.P.; Hu, J.; Shu, Y.; Wu, R. Coastal upwelling in summer 2000 in the northeastern South China Sea. J. Geophys. Res. Oceans. 2012, 117, C04009. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Gan, J.; Dai, M.; Cheung, A.Y. The influence of coastal upwelling and a river plume on the subsurface chlorophyll maximum over the shelf of the northeastern South China Sea. J. Mar. Syst. 2010, 82, 35–46. [Google Scholar] [CrossRef]
- Chen, Z.; Pan, J.; Jiang, Y.; Lin, H. Far-reaching transport of Pearl River plume water by upwelling jet in the northeastern South China Sea. J. Mar. Syst. 2017, 173, 60–69. [Google Scholar] [CrossRef]
- Xue, H.J.; Chai, F. Coupled physical-biological model for the Pearl River Estuary: A phosphate limited subtropical ecosystem. In Estuarine and Coastal Modeling (2001); American Society of Civil Engineers: Reston, VA, USA, 2002; pp. 913–928. [Google Scholar]
- Zheng, G.M.; Tang, D. Offshore and nearshore chlorophyll increases induced by typhoon winds and subsequent terrestrial rainwater runoff. Mar. Ecol Prog. Ser. 2007, 333, 61–74. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Xu, D.; Bai, Y.; Pan, D.; Chen, C.T.A.; Chen, X.; Gong, F. Eddy-entrained Pearl River plume into the oligotrophic basin of the South China Sea. Cont. Shelf. Res. 2016, 124, 117–124. [Google Scholar] [CrossRef]
- Jing, Z.Y.; Qi, Y.Q.; Hua, Z.L.; Zhang, H. Numerical study on the summer upwelling system in the northern continental shelf of the South China Sea. Cont. Shelf. Res. 2009, 29, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Lü, X.; Qiao, F.; Wang, G.; Xia, C.; Yuan, Y. Upwelling off the west coast of Hainan Island in summer: Its detection and mechanisms. Geophys. Res. Lett. 2008, 35, L02604. [Google Scholar] [CrossRef]
- Wang, D.; Shu, Y.; Xue, H.; Hu, J.; Chen, J.; Zhuang, W.; Zu, T.; Xu, J. Relative contributions of local wind and topography to the coastal upwelling intensity in the northern South China Sea. J. Geophys. Res. Oceans. 2014, 119, 2550–2567. [Google Scholar] [CrossRef]
- Forryan, A.; Garabato, A.C.N.; Vic, C.; Nurser, A.G.; Hearn, A.R. Galápagos upwelling driven by localized wind–front interactions. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Liu, K.S.; Chan, J.C. Recent increase in extreme intensity of tropical cyclones making landfall in South China. Clim. Dyn. 2020, 55, 1059–1074. [Google Scholar] [CrossRef]
- Lin, I.; Liu, W.T.; Wu, C.C.; Wong, G.T.; Hu, C.; Chen, Z.; Liang, D.-W.; Yang, Y.; Liu, K.K. New evidence for enhanced ocean primary production triggered by tropical cyclone. Geophys. Res. Lett. 2003, 30, 1718. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.M.; Oey, L.Y. Right-side cooling and phytoplankton bloom in the wake of a tropical cyclone. J. Geophys. Res. Oceans 2015, 120, 5735–5748. [Google Scholar] [CrossRef]
- Lin, Y.C.; Oey, L.Y. Rainfall-enhanced blooming in typhoon wakes. Sci. Rep. 2016, 6, 31310. [Google Scholar] [CrossRef] [Green Version]
- Pan, G.; Chai, F.; Tang, D.; Wang, D. Marine phytoplankton biomass responses to typhoon events in the South China Sea based on physical-biogeochemical model. Ecol. Modell. 2017, 356, 38–47. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, S.; Chen, F.; Ma, Y.; Jiang, C.; Yu, J. Influence of sequential tropical cyclones on phytoplankton blooms in the northwestern South China Sea. J. Oceanol. Limnol. 2021, 39, 14–25. [Google Scholar] [CrossRef]
- Zhao, H.; Tang, D.; Wang, D. Phytoplankton blooms near the Pearl River estuary induced by Typhoon Nuri. J. Geophys. Res. Oceans 2009, 114, C12027. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, D.; Evgeny, M. Chlorophyll concentration response to the typhoon wind-pump induced upper ocean processes considering air–sea heat exchange. Remote Sens. 2019, 11, 1825. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Li, J.; Sun, L.; Wang, G.; Tang, D.; Huang, P.; Yan, H.; Gao, S.; Liu, C.; Gao, Z. Basin-wide responses of the South China sea environment to super typhoon mangkhut (2018). Sci. Total Environ. 2020, 731, 139093. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Composite of typhoon-induced sea surface temperature and chlorophyll-a responses in the South China Sea. J. Geophys. Res. Oceans 2020, 125, e2020JC016243. [Google Scholar] [CrossRef]
- Liu, F.; Tang, S. Influence of the interaction between typhoons and oceanic mesoscale eddies on phytoplankton blooms. J. Geophys. Res. Oceans 2018, 123, 2785–2794. [Google Scholar] [CrossRef]
- Xu, F.; Yao, Y.; Oey, L.; Lin, Y. Impacts of pre-existing ocean cyclonic circulation on sea surface chlorophyll-a concentrations off northeastern Taiwan following episodic typhoon passages. J. Geophys. Res. Oceans 2017, 122, 6482–6497. [Google Scholar] [CrossRef]
- Babin, S.M.; Carton, J.A.; Dickey, T.D.; Wiggert, J.D. Satellite evidence of hurricane-induced phytoplankton blooms in an oceanic desert. J. Geophys. Res. Oceans 2004, 109, C03043. [Google Scholar] [CrossRef]
- Kuttippurath, J.; Sunanda, N.; Martin, M.V.; Chakraborty, K. Tropical storms trigger phytoplankton blooms in the deserts of north Indian Ocean. NPJ Clim. Atmos. Sci. 2021, 4, 1–12. [Google Scholar] [CrossRef]
- Sun, L.; Yang, Y.J.; Xian, T.; Lu, Z.M.; Fu, Y.F. Strong enhancement of chlorophyll a concentration by a weak typhoon. Mar. Ecol Prog. Ser. 2010, 404, 39–50. [Google Scholar] [CrossRef]
- Ye, H.J.; Sui, Y.; Tang, D.L.; Afanasyev, Y.D. A subsurface chlorophyll a bloom induced by typhoon in the South China Sea. J. Mar. Syst. 2013, 128, 138–145. [Google Scholar] [CrossRef]
- Lee, J.H.; Moon, J.H.; Kim, T. Typhoon-triggered phytoplankton bloom and associated upper-ocean conditions in the northwestern Pacific: Evidence from satellite remote sensing, Argo profile, and an ocean circulation model. J. Mar. Sci. Eng. 2020, 8, 788. [Google Scholar] [CrossRef]
- Zhao, H.; Han, G.; Zhang, S.; Wang, D. Two phytoplankton blooms near Luzon Strait generated by lingering Typhoon Parma. J. Geophys. Res. Biogeosci. 2013, 118, 412–421. [Google Scholar] [CrossRef]
- Chai, F.; Wang, Y.; Xing, X.; Yan, Y.; Xue, H.; Wells, M.; Boss, E. A limited effect of sub-tropical typhoons on phytoplankton dynamics. Biogeosciences 2021, 18, 849–859. [Google Scholar] [CrossRef]
- Lin, I.I. Typhoon-induced phytoplankton blooms and primary productivity increase in the western North Pacific subtropical ocean. J. Geophys. Res. Oceans 2012, 117, C03039. [Google Scholar] [CrossRef]
- Li, Y.; Ye, X.; Wang, A.; Li, H.; Chen, J.; Qiao, L. Impact of Typhoon Morakot on chlorophyll a distribution on the inner shelf of the East China Sea. Mar. Ecol Prog. Ser. 2013, 483, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Qiu, D.; Zhong, Y.; Chen, Y.; Tan, Y.; Song, X.; Huang, L. Short-term phytoplankton dynamics during typhoon season in and near the Pearl River Estuary, South China Sea. J. Geophys. Res. Biogeosci. 2019, 124, 274–292. [Google Scholar] [CrossRef]
- Chang, J.; Chung, C.; Gong, G. Influences of cyclones on chlorophyll a concentration and Synechococcus abundance in a subtropical western Pacific coastal ecosystem. Oceanogr. Lit. Rev. 1997, 4, 346. [Google Scholar] [CrossRef] [Green Version]
- Pan, A.; Guo, X.; Xu, J.; Huang, J.; Wan, X. Responses of Guangdong coastal upwelling to the summertime typhoons of 2006. Sci. China Earth Sci. 2012, 55, 495–506. [Google Scholar] [CrossRef]
- Shen, D.; Li, X.; Wang, J.; Bao, S.; Pietrafesa, L.J. Dynamical Ocean responses to Typhoon Malakas (2016) in the vicinity of Taiwan. J. Geophys. Res. Oceans 2021, 126, e2020JC016663. [Google Scholar] [CrossRef]
- Mao, Y.; Sun, J.; Guo, C.; Wei, Y.; Wang, X.; Yang, S.; Wu, C. Effects of typhoon Roke and Haitang on phytoplankton community structure in northeastern South China Sea. Ecosyst. Health Sust. 2019, 5, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Gohin, F. Annual cycles of chlorophyll-a, non-algal suspended particulate matter, and turbidity observed from space and in-situ in coastal waters. Ocean Sci. 2011, 7, 705–732. [Google Scholar] [CrossRef] [Green Version]
- Wentz, F.J.; Gentemann, C.; Smith, D.; Chelton, D. Satellite measurements of sea surface temperature through clouds. Science 2000, 288, 847–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentemann, C.L.; Wentz, F.J.; Mears, C.A.; Smith, D.K. In-Situ validation of Tropical Rainfall Measuring Mission microwave sea surface temperatures. J. Geophys. Res. Oceans 2004, 109, C04021. [Google Scholar] [CrossRef] [Green Version]
- Meissner, T.; Wentz, F.J.; Le Vine, D.M. The Salinity Retrieval Algorithms for the NASA Aquarius Version 5 and SMAP Version 3 Releases. Remote Sens. 2018, 10, 1121. [Google Scholar] [CrossRef] [Green Version]
- Atlas, R.; Hoffman, R.N.; Ardizzone, J.; Leidner, S.M.; Jusem, J.C.; Smith, D.K.; Gombos, D. A cross-calibrated, multiplatform ocean surface wind velocity product for meteorological and oceanographic applications. Bull. Am. Meteorol. Soc. 2011, 92, 157–174. [Google Scholar] [CrossRef]
- Huffman, G.J.; Adler, R.F.; Bolvin, D.T.; Nelkin, E.J. The TRMM multi-satellite precipitation analysis (TMPA). In Satellite Rainfall Applications for Surface Hydrology; Springer: Dordrecht, The Netherlands, 2010; pp. 3–22. [Google Scholar] [CrossRef] [Green Version]
- Price, J.F. Upper Ocean response to a hurricane. J. Phys. Oceanogr. 1981, 11, 153–175. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Li, Y.X.; Yang, Y.J.; Wu, Q.; Chen, X.T.; Li, Q.Y.; Li, Y.-B.; Xian, T. Effects of super typhoons on cyclonic ocean eddies in the western North Pacific: A satellite data-based evaluation between 2000 and 2008. J. Geophys. Res. Oceans 2014, 119, 5585–5598. [Google Scholar] [CrossRef]
- Mei, W.; Pasquero, C.; Primeau, F. The effect of translation speed upon the intensity of tropical cyclones over the tropical ocean. Geophys. Res. Lett. 2012, 39, L07801. [Google Scholar] [CrossRef] [Green Version]
- Gan, J.; Li, L.; Wang, D.; Guo, X. Interaction of a river plume with coastal upwelling in the northeastern South China Sea. Cont. Shelf. Res. 2009, 29, 728–740. [Google Scholar] [CrossRef]
- Bai, Y.; Huang, T.H.; He, X.; Wang, S.L.; Hsin, Y.C.; Wu, C.R.; Zhai, W.; Lui, H.-T.; Chen, C.T.A. Intrusion of the Pearl River plume into the main channel of the Taiwan Strait in summer. J. Sea Res. 2015, 95, 1–15. [Google Scholar] [CrossRef]
- Xu, W.; Jiang, H.; Kang, X. Rainfall asymmetries of tropical cyclones prior to, during, and after making landfall in South China and Southeast United States. Atmos. Res. 2014, 139, 18–26. [Google Scholar] [CrossRef]
- Klotz, B.W.; Jiang, H. Examination of surface wind asymmetries in tropical cyclones. Part I: General structure and wind shear impacts. Mon. Weather Rev. 2017, 145, 3989–4009. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, D.; Tang, S.; Morozov, E.; Liang, W.; Sui, Y. A case study of Chlorophyll a response to tropical cyclone Wind Pump considering Kuroshio invasion and air-sea heat exchange. Sci. Total Environ. 2020, 741, 140290. [Google Scholar] [CrossRef]
- Domingues, R.B.; Guerra, C.C.; Barbosa, A.B.; Galvao, H.M. Are nutrients and light limiting summer phytoplankton in a temperate coastal lagoon? Aquatic Ecol. 2015, 49, 127–146. [Google Scholar] [CrossRef]
- Pennock, J.R.; Sharp, J.H. Temporal alternation between light-and nutrient imitation of phytoplankton production in a coastal plain estuary. Mar. Ecol. Prog. Ser. 1994, 111, 275–288. [Google Scholar] [CrossRef]
- Teixeira, I.G.; Arbones, B.; Froján, M.; Nieto-Cid, M.; Álvarez-Salgado, X.A.; Castro, C.G.; Fernández, E.; Sobrino, C.; Teira, E.; Figueiras, F.G. Response of phytoplankton to enhanced atmospheric and riverine nutrient inputs in a coastal upwelling embayment. Estuar. Coast. Shelf Sci. 2018, 210, 132–141. [Google Scholar] [CrossRef]
- Dokulil, M.T. Environmental control of phytoplankton productivity in turbulent turbid systems. In Phytoplankton in Turbid Environments: Rivers and Shallow Lakes; Springer: Dordrecht, The Netherlands, 1994; pp. 65–72. [Google Scholar] [CrossRef]

















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Zhang, S. Effect of Summer Typhoon Linfa on the Chlorophyll-a Concentration in the Continental Shelf Region of Northern South China Sea. J. Mar. Sci. Eng. 2021, 9, 794. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080794
Wang T, Zhang S. Effect of Summer Typhoon Linfa on the Chlorophyll-a Concentration in the Continental Shelf Region of Northern South China Sea. Journal of Marine Science and Engineering. 2021; 9(8):794. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080794
Chicago/Turabian StyleWang, Tongyu, and Shuwen Zhang. 2021. "Effect of Summer Typhoon Linfa on the Chlorophyll-a Concentration in the Continental Shelf Region of Northern South China Sea" Journal of Marine Science and Engineering 9, no. 8: 794. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080794