
Culture Growth of the Cyanobacterium Phormidium sp. in Various Salinity and Light Regimes and Their Influence on Its Phycocyanin and Other Pigments Content


Abstract
:1. Introduction
2. Materials and Methods
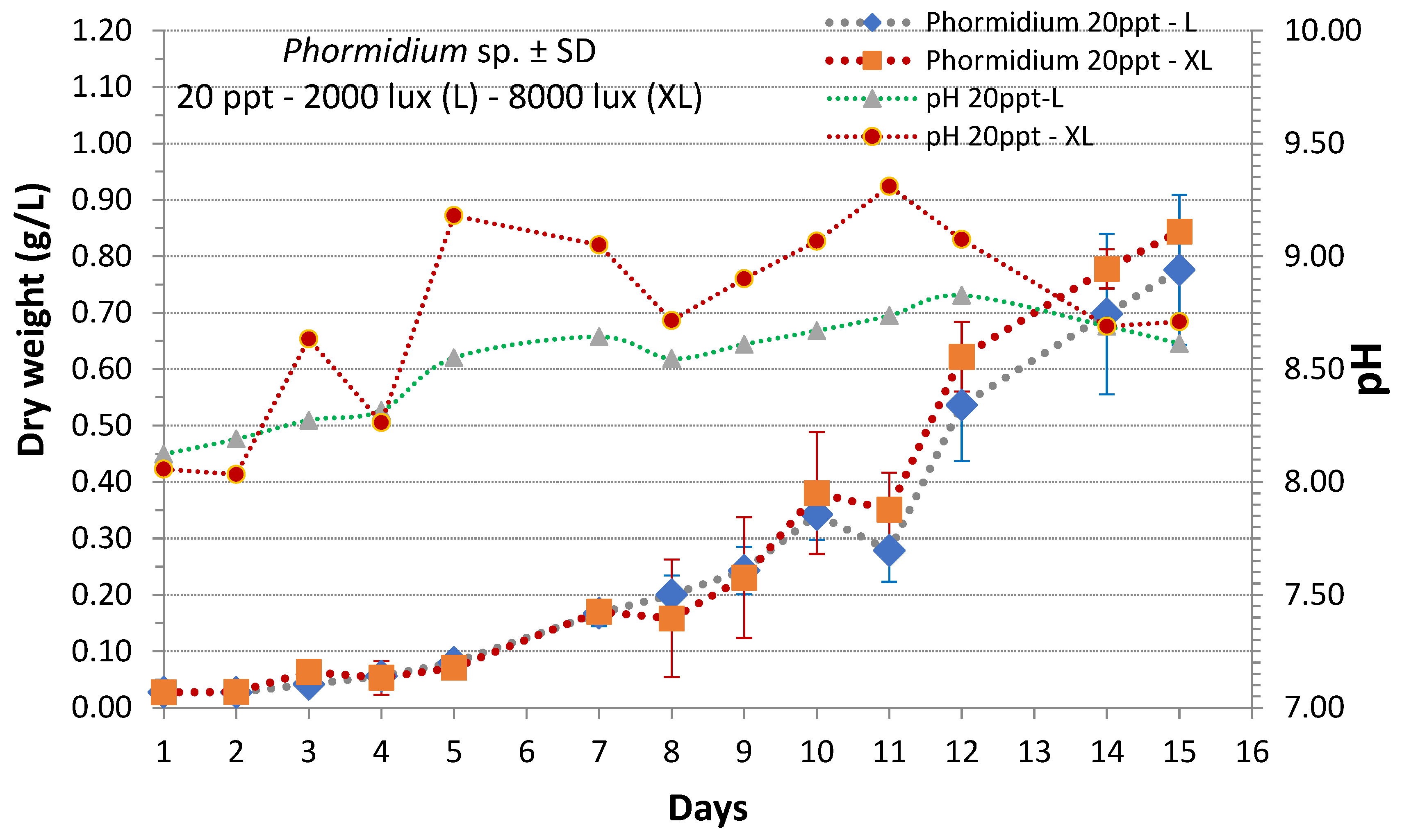
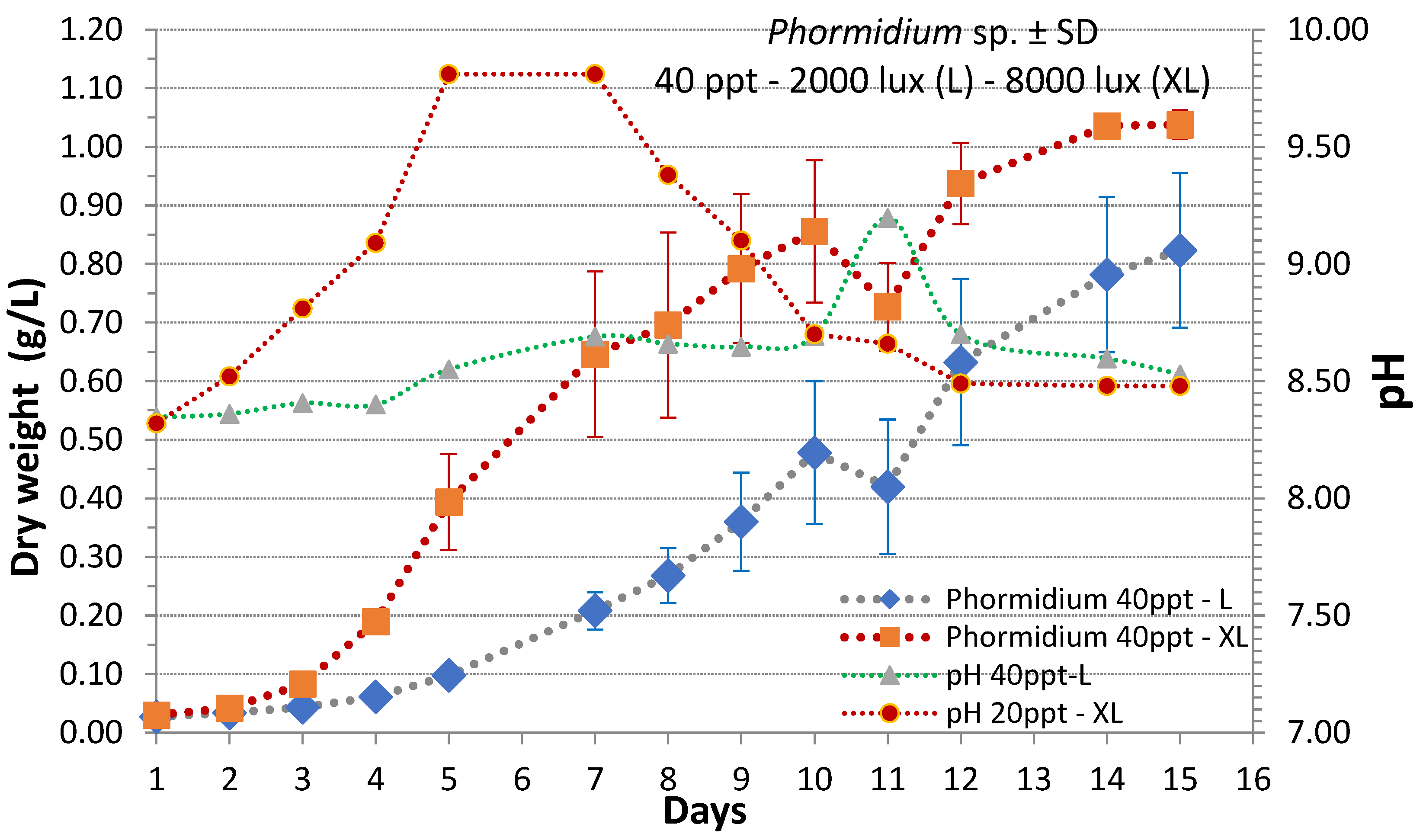
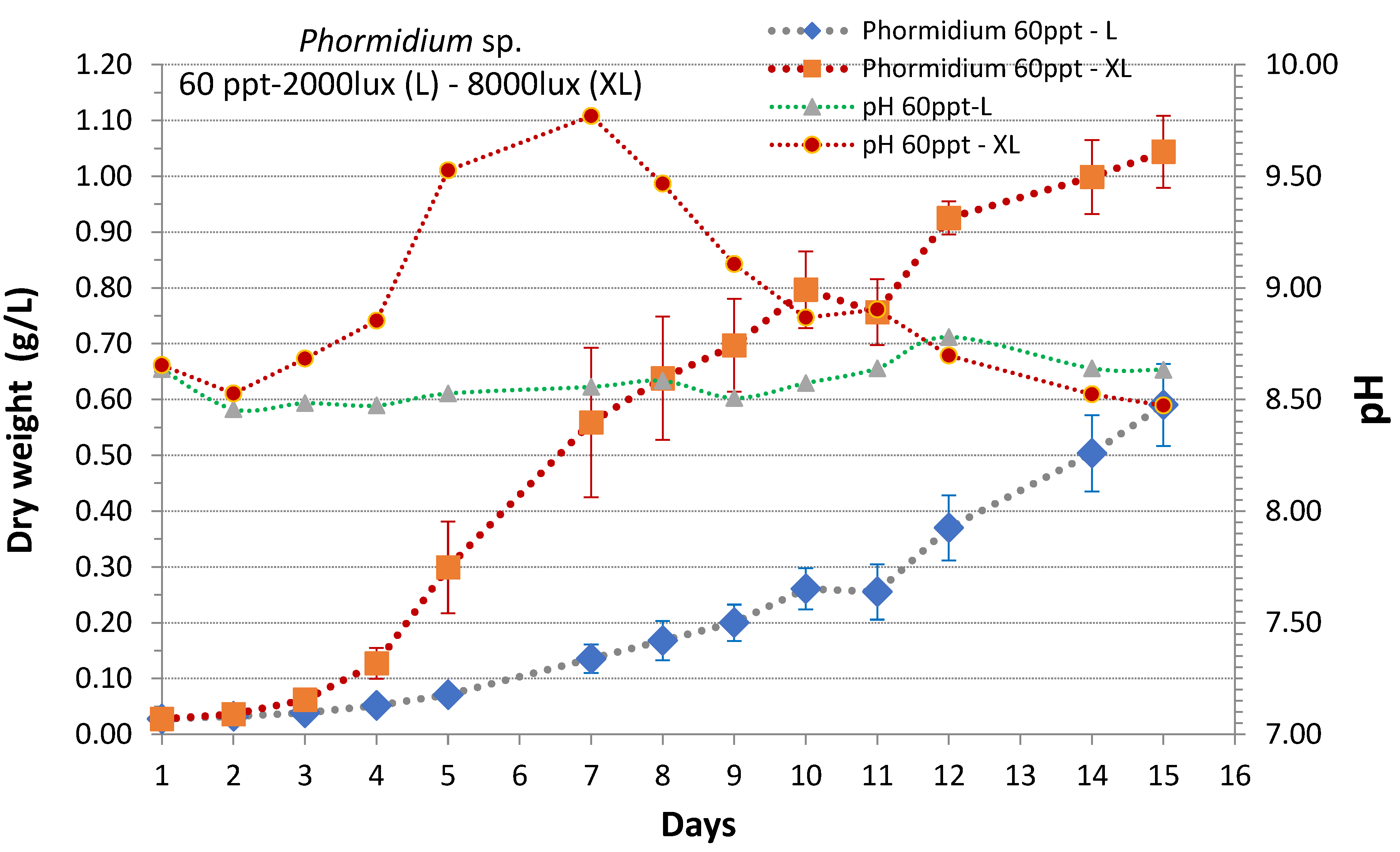
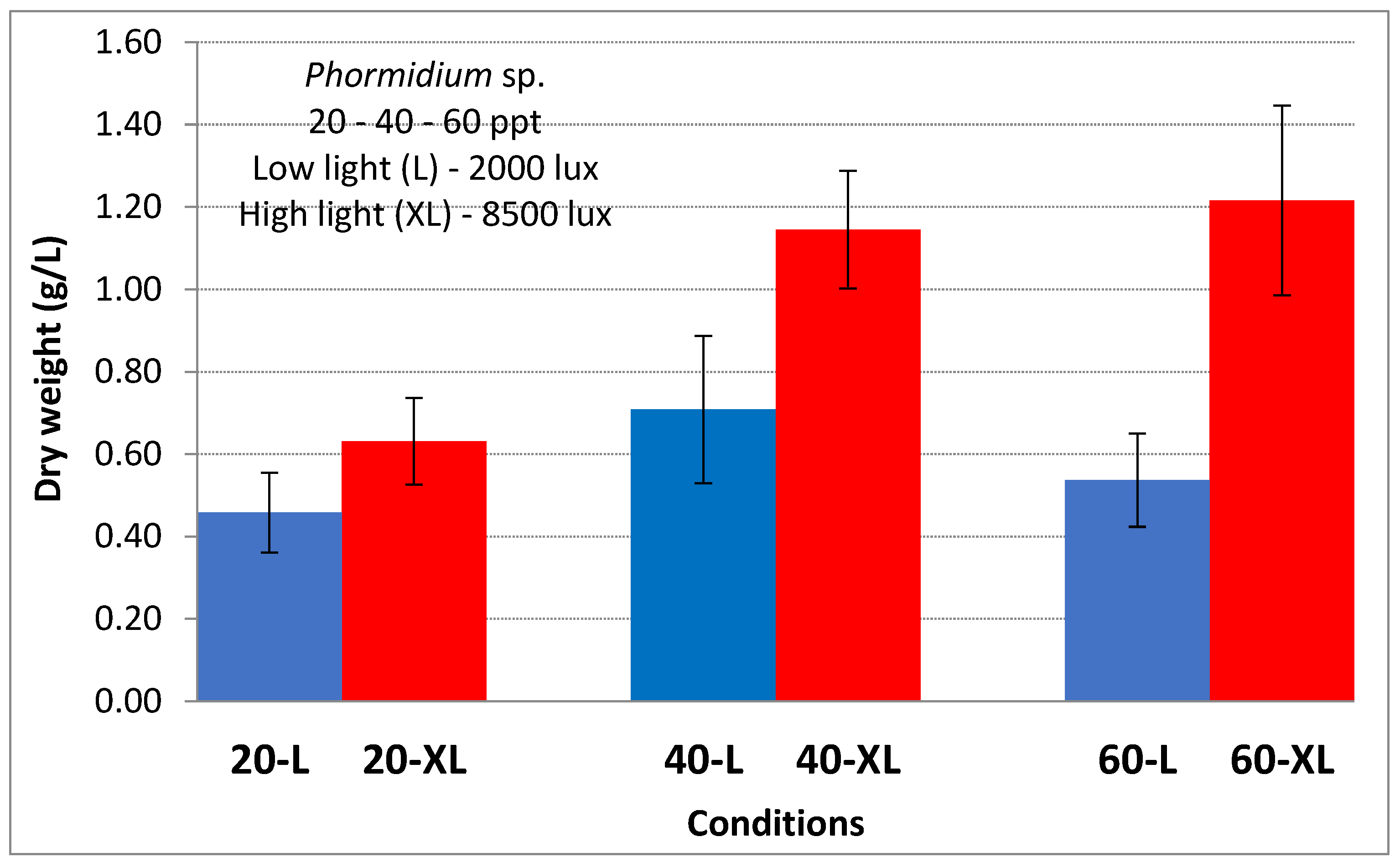
3. Results
4. Discussion
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae—A review. J. Algal Biomass Utln. 2012, 3, 89–100. [Google Scholar]
- Sun, H.; Zhao, W.; Mao, X.; Li, Y.; Wu, T.; Chen, F. High-value biomass from microalgae production platforms: Strategies and progress based on carbon metabolism and energy conversion. Biotechnol. Biofuels 2018, 11, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Pathak, J.; Rajneesh, M.P.K.; Singh, S.P.; Häder, D.P.; Sinha, R.P. Cyanobacterial Farming for Environment Friendly Sustainable Agriculture Practices: Innovations and Perspectives. Front. Environ. Sci. 2018, 6, 1–13. [Google Scholar] [CrossRef]
- Saranraj, P.; Sivasakthi, S. Spirulina platensis-Food for future: A review. Asian J. Pharm. Sci. Technol. 2014, 4, 26–33. [Google Scholar]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Hotos, G.N. A Preliminary Survey on the Planktonic Biota in a Hypersaline Pond of Messolonghi Saltworks (W. Greece). Diversity 2021, 13, 270. [Google Scholar] [CrossRef]
- Eriksen, N.T. Production of phycocyanin-a pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef]
- Hotos, G.N.; Avramidou, D.; Bekiari, V. Calibration Curves of Culture Density Assessed by Spectrophotometer for Three Microalgae (Nephroselmis sp., Amphidinium carterae and Phormidium sp.). Eur. J. Biol. Biotechnol. 2020, 1, 1–7. [Google Scholar] [CrossRef]
- Griffiths, M.J.; Garcin, C.; van Hille, R.P.; Harrison, S.T. Interference by pigment in the estimation of microalgal biomass concentration by optical density. J. Microbiol. Methods 2011, 85, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Zavřel, T.; Sinetova, M.A.; Červený, J. Measurement of Chlorophyll a and Carotenoids Concentration in Cyanobacteria. Bio-Protocol 2015, 5, e1467. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313, ISSN 1618–1328. [Google Scholar] [CrossRef]
- Arashiro, L.T.; Boto-Ordóñez, M.; Van Hulle, S.W.H.; Ferrer, I.; Garfí, M.; Rousseau, D.P.L. Natural pigments from microalgae grown in industrial wastewater. Bioresour. Technol. 2020, 303, 122894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraes, C.C.; Sala, L.; Cerveira, P.G.; Kalil, J.S. C-Phycocyanin extraction from Spirulina platensis wet biomass. Braz. J. Chem. Eng. 2011, 28, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.R.; Dayananda, C.; Sarada, R.; Shamala, T.R.; Ravishankar, G.A. Effect of salinity on growth of green alga Botryococcus braunii and its constituents. Bioresour. Technol. 2007, 98, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Groom, M.J.; Gray, E.M.; Townsend, P.A. Biofuels and biodiversity: Principles for creating better policies for biofuel production. Conserv. Biol. 2008, 22, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Kitaya, Y.; Xiao, L.; Masuda, A.; Ozawa, T.; Tsuda, M.; Omasa, K. Effects of temperature, photosynthetic photon flux density, photoperiod and O2 and CO2 concentrations on growth rates of the symbiotic dinoflagellate, Amphidinium sp. J. Appl. Phycol. 2008, 20, 737–742. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Xue, S.; Su, Z.; Cong, W. Growth of Spirulina platensis enhanced under intermittent illumination. J. Biotechnol. 2011, 151, 271–277. [Google Scholar] [CrossRef]
- Parmar, A.; Singh, N.K.; Pandey, A.; Gnansounou, E.; Madamwar, D. Cyanobacteria and microalgae: A positive prospect for biofuels. Bioresour. Technol. 2011, 102, 10163–10172. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.H.; Huang, S.W.; Chen, C.Y.; Hasunuma, T.; Kondo, A.; Chang, J.S. Characterization and optimization of carbohydrate production from an indigenous microalga Chlorella vulgaris FSP-E. Bioresour. Technol. 2013, 135, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Wahidin, S.; Idris, A.; Shaleh, S.R. The influence of light intensity and photoperiod on the growth and lipid content of microalgae Nannochloropsis sp. Bioresour. Technol. 2013, 129, 7–11. [Google Scholar] [CrossRef]
- Marquardt, J.; Palinska, K.A. Genotypic and phenotypic diversity of cyanobacteria assigned to the genus Phormidium (Oscillatoriales) from different habitats and geographical sites. Arch. Microbiol. 2007, 187, 397–413. [Google Scholar] [CrossRef]
- Kumar, J.; Singh, V.P.; Prasad, S.M. NaCl-induced physiological and biochemical changes in two cyanobacteria Nostoc muscorum and Phormidium foveolarum acclimatized to different photosynthetically active radiation. J. Photochem. Photobiol. B 2015, 151, 221–232. [Google Scholar] [CrossRef]
- Bhandari, R.; Sharma, P.K. High-light-induced changes on photosynthesis, pigments, sugars, lipids and antioxidant enzymes in freshwater (Nostoc spongiaeforme) and marine (Phormidium corium) cyanobacteria. Photochem. Photobiol. 2006, 82, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Nagasathya, A.; Thajuddin, N. Cyanobacterial Diversity in the Hypersaline Environment of the Saltpans of Southeastern Coast of India. Asian J. Plant Sci. 2008, 7, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Rosales, N.; Ortega, J.; Mora, R.; Morales, E. Influence of salinity on the growth and biochemical composition of the cyanobacterium Synechococcus sp. Cienc. Mar. 2005, 31, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Dias, F.; Antunes, J.T.; Ribeiro, T.; Azevedo, J.; Vasconcelos, V.; Leão, P.N. Cyanobacterial Allelochemicals But Not Cyanobacterial Cells Markedly Reduce Microbial Community Diversity. Front. Microbiol. 2017, 8, 1495. [Google Scholar] [CrossRef]
- Silveira, S.T.; Burkert, J.F.; Costa, J.A.; Burkert, C.A.; Kalil, S.J. Optimization of phycocyanin extraction from Spirulina platensis using factorial design. Bioresour. Technol. 2007, 98, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Göksan, T.; Zekeriyaoğlu, A.; Ak, I. The Growth of Spirulina platensis in Different Culture Systems under Greenhouse Condition. Turk. J. Biol. 2006, 31, 47–52. [Google Scholar]
- Pandey, J.P.; Pathak, N.; Tiwari, A. Standardization of pH and Light Intensity for the Biomass Production of Spirulina platensis. J. Algal Biomass Util. 2010, 1, 93–102. [Google Scholar]
- Walter, A.; de Carvalho, J.C.; Soccol, V.T.; Bisinella de Faria, A.B.; Ghiggi, V.; Soccol, C.R. Study of Phycocyanin Production from Spirulina platensis Under Different Light Spectra. Brasilian Arch. Biol. Technol. 2011, 54, 675–682. [Google Scholar] [CrossRef]
- Singh, N.K.; Parmar, A.; Madamwar, D. Optimization of medium components for increased production of C-phycocyanin from Phormidium ceylanicum and its purification by single step process. Bioresour. Technol. 2009, 100, 1663–1669. [Google Scholar] [CrossRef]
- Rodrigues, D.B.; Menezes, C.R.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Bioactive pigments from microalgae Phormidium autumnale. Food Res. Int. 2015, 77, 273–279. [Google Scholar] [CrossRef]
- Healey, F.P. Physiological indicators of nutrient deficiency in algae. Fish. Mar. Serv. Res. Dev. Tech. Rep. 1975, 585, 30. [Google Scholar] [CrossRef]
- Nicholls, K.H.; Dillon, P.J. An Evaluation of Phosphorus-Chlorophyll-Phytoplankton Relationships for Lakes. Nternationale Rev. Der Gesamten Hydrobiol. Hydrogr. 1978, 63, 141–154. [Google Scholar] [CrossRef]
- Walsby, A. Microalgae: Biotechnology and Microbiology. Exp. Agric. 1995, 31, 112. [Google Scholar] [CrossRef]
- Fresnedo, O.; Gomez, R.; Serra, J.L. Carotenoid composition in the cyanobacterium Phormidium laminosum. Effect of nitrogen starvation. FEBS Lett. 1991, 282, 300–304. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Yu, H.; Brown, M.T. Solar PAR and UV radiation affects the physiology and morphology of the cyanobacterium Anabaena sp. PCC 7120. J. Photochem. Photobiol. B 2007, 89, 117–124. [Google Scholar] [CrossRef]
- Schubert, H.; Fulda, S.; Hagemann, M. Effects of adaptation to different salt concentrations on photosynthesis and pigmentation of the cyanobacterium Synechocystis sp. PCC 6803. J. Plant Physiol. 1993, 142, 291–295. [Google Scholar] [CrossRef]
- Lu, C.; Vonshak, A. Effects of salinity stress on photosystem II function in cyanobacterial Spirulina platensis cells. Physiol. Plant. 2002, 114, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Murata, N.; Miyao, M. Extrinsic membrane proteins in the photosynthetic oxygen-evolving complex. Trends Biochem. Sci. 1985, 10, 122–124. [Google Scholar] [CrossRef]
- Parida, A.; Das, A.; Mittra, B. Effects of NaCl Stress on the Structure, Pigment Complex Composition, and Photosynthetic Activity of Mangrove Bruguiera parviflora Chloroplasts. Photosynthetica 2003, 41, 191–200. [Google Scholar] [CrossRef]
- Zerveas, S.; Mente, M.S.; Tsakiri, D.; Kotzabasis, K. Microalgal photosynthesis induces alkalization of aquatic environment as a result of H+ uptake independently from CO2 concentration—New perspectives for environmental applications. J. Environ. Manag. 2021, 289, 112546. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Var. | 20 ppt-L | 20 ppt-XL | 40 ppt-L | 40 ppt-XL | 60 ppt-L | 60 ppt-XL |
|---|---|---|---|---|---|---|
| μmax | a 0.363 ± 0.03 | b 0.203 ± 0.012 | c 0.404 ± 0.045 | a,c 0.401 ± 0.010 | d 0.317 ± 0.038 | e 0.491 ± 0.025 |
| tg | a 1.922 ± 0.169 | b 3.426 ± 0.199 | a,c 1.739 ± 0.207 | a,c 1.791 ± 0.359 | d 2.222 ± 0.290 | e 1.415 ± 0.070 |
| days | 4th–7th | 4th–7th | 4th–7th | 4th–7th | 4th–7th | 4th–7th |
| n | 27 | 27 | 27 | 27 | 27 | 27 |
| Var. | 20 ppt-L | 20 ppt-XL | 40 ppt-L | 40 ppt-XL | 60 ppt-L | 60 ppt-XL |
|---|---|---|---|---|---|---|
| PC | a 0.277 ± 0.061 | b 0.0011 ± 0.000 | c 0.054 ± 0.0192 | d 0.0037 ± 0.004 | e 0.107 ± 0.039 | d,f 0.0086 ± 0.09 |
| min-max | 0.216–0.350 | 0.001–0.0013 | 0.033–0.074 | 0.0002–0.0075 | 0.069–0.146 | 0.0005–0.0175 |
| PCyield | a 22.658 ± 1.44 | b 0.084 ± 0.012 | c 5.750 ± 2.40 | d 0.393 ± 0.413 | e 9.631 ± 0.362 | f 0.645 ± 0.664 |
| min-max | 21.30–24.06 | 0.075–0.098 | 3.428–8.001 | 0.014–0.837 | 9.284–10.006 | 0.030–1.276 |
| PE | a 0.0462 ± 0.029 | b 0.0004 ± 0.00 | c 0.0056 ± 0.0015 | d 0.0013 ± 0.0009 | e 0.0107 ± 0.0056 | d,f 0.0024 ± 0.002 |
| min-max | 0.0193–0.0744 | 0.0004 | 0.0032–0.0073 | 0.0002–0.0023 | 0.005–0.0162 | 0.0003–0.0053 |
| PEyield | a 3.6030 ± 1.803 | b 0.0307 ± 0.001 | c 0.575 ± 0.1641 | d 0.1392 ± 0.1221 | e 0.9662 ± 0.1141 | f 0.1787 ± 0.173 |
| min-max | 1.9325–5.2468 | 0.0291–0.0319 | 0.4101–0.7311 | 0.0148–0.2588 | 0.8188–1.0324 | 0.0202–0.3640 |
| AllPC | 0.1055 ± 0.053 | 0.0004 ± 0.00 | 0.0144 ± 0.0035 | 0.0028 ± 0.0025 | 0.0654 ± 0.0641 | 0.0058 ± 0.006 |
| Chl-a | a 8.146 ± 0.298 | b 3.457 ± 0.136 | c 6.008 ± 0.387 | d 4.808 ± 0.378 | d,e 4.272 ± 0.855 | f 1.438 ± 0.225 |
| min-max | 7.793–8.433 | 3.334–3.599 | 5.482–6.306 | 4.274–5.164 | 3.179–5.002 | 1.169–1.670 |
| Carot | a 2.032 ± 0.183 | b 1.210 ± 0.026 | c 1.567 ± 0.090 | d 1.976 ± 0.114 | b,c,e 1.445 ± 0.304 | b,f 1.153 ± 0.274 |
| min-max | 1.760–2.156 | 1.186–1.245 | 1.433–1.624 | 1.825–2.100 | 0.993–1.653 | 0.747–1.331 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hotos, G.N. Culture Growth of the Cyanobacterium Phormidium sp. in Various Salinity and Light Regimes and Their Influence on Its Phycocyanin and Other Pigments Content. J. Mar. Sci. Eng. 2021, 9, 798. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080798
Hotos GN. Culture Growth of the Cyanobacterium Phormidium sp. in Various Salinity and Light Regimes and Their Influence on Its Phycocyanin and Other Pigments Content. Journal of Marine Science and Engineering. 2021; 9(8):798. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080798
Chicago/Turabian StyleHotos, George N. 2021. "Culture Growth of the Cyanobacterium Phormidium sp. in Various Salinity and Light Regimes and Their Influence on Its Phycocyanin and Other Pigments Content" Journal of Marine Science and Engineering 9, no. 8: 798. https://0-doi-org.brum.beds.ac.uk/10.3390/jmse9080798