Exploratory Sampling of Spalting Fungi in the Southern Peruvian Amazon Forest

 ,
,
Abstract
:1. Introduction
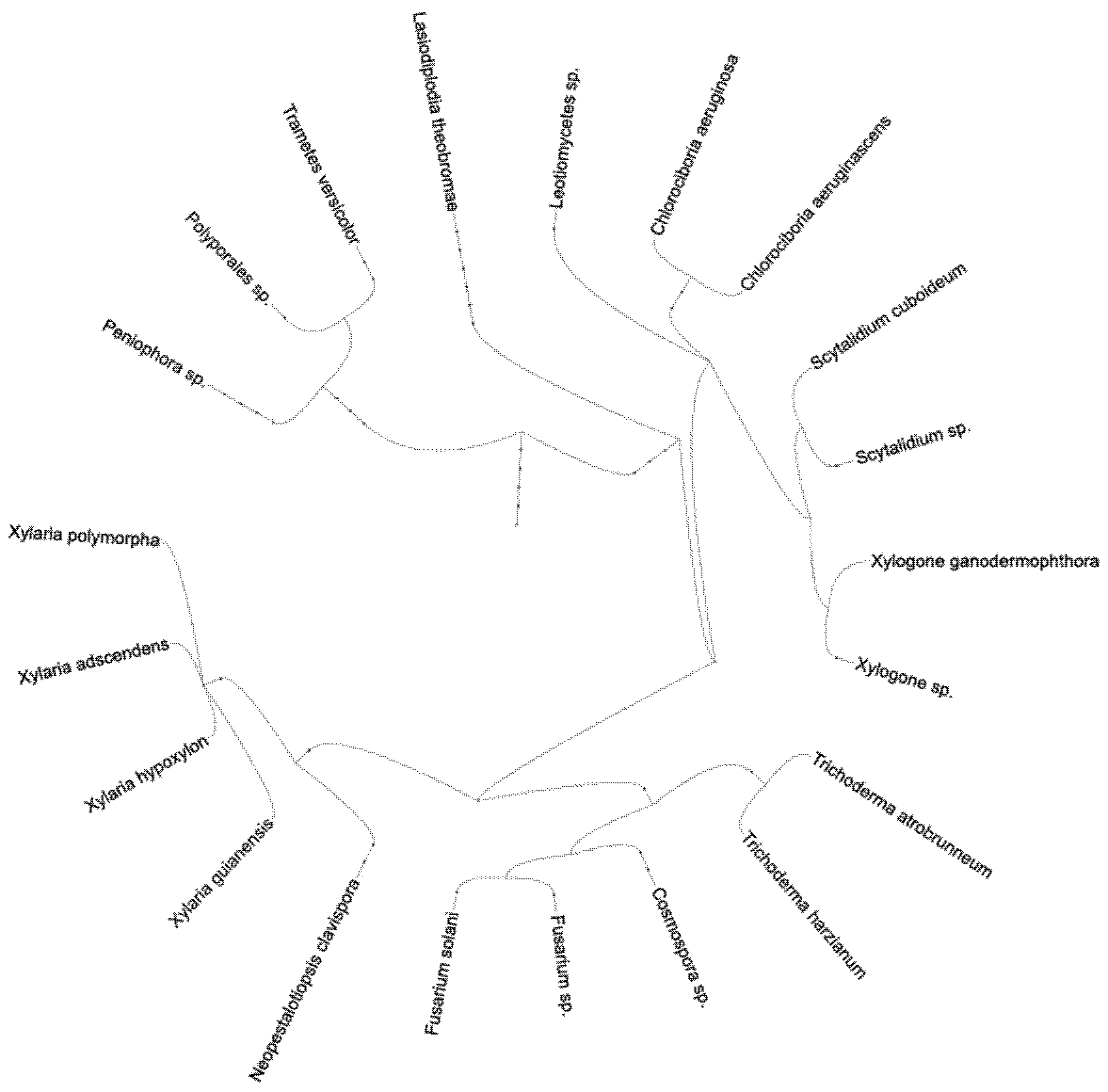
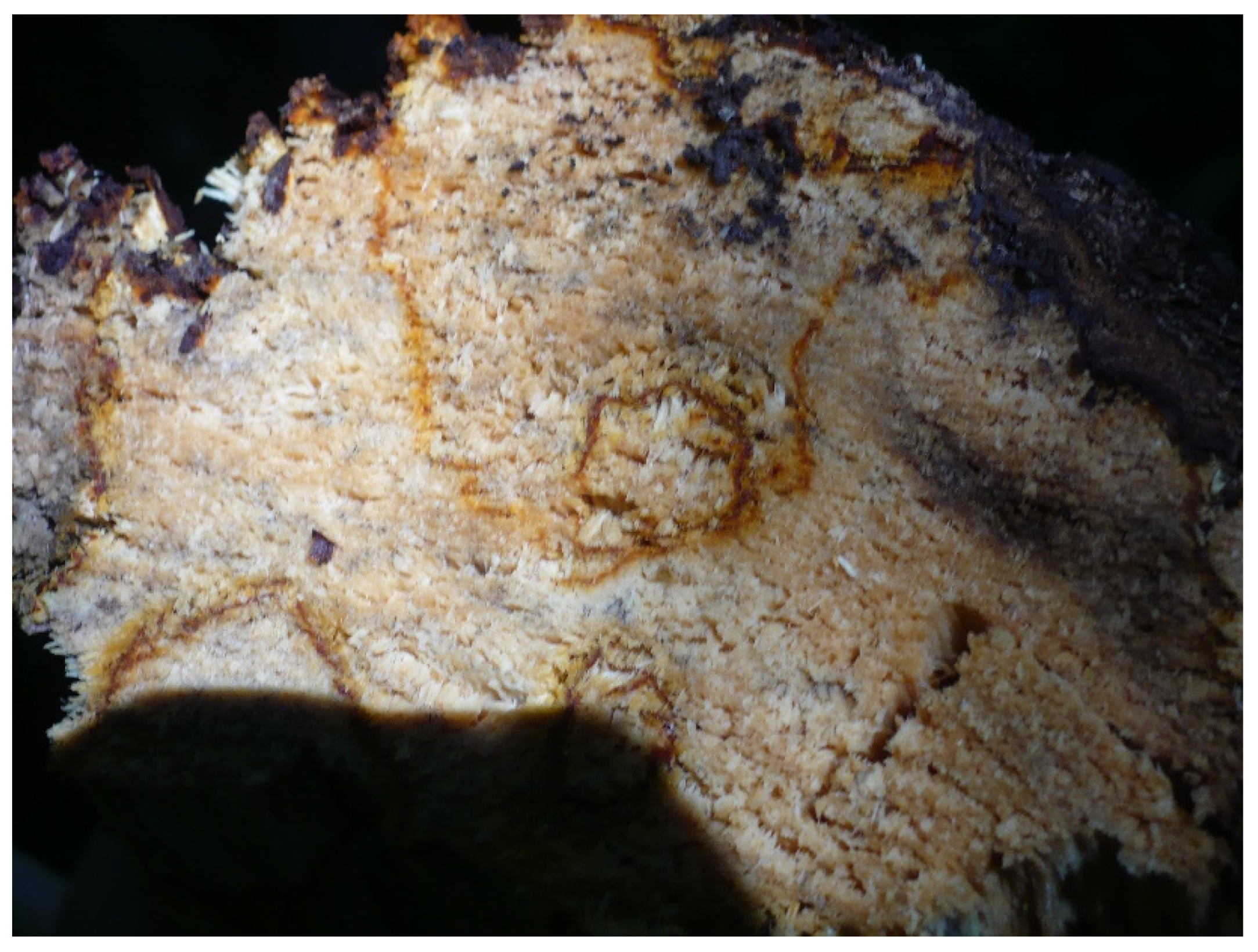
2. Results
3. Discussion
4. Materials and Methods
4.1. Collection
4.2. Fungal Isolation
4.3. DNA Sequencing
4.4. Pairwise Sequence Alignment
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sánchez, S.; Llosa, G.; Pulido, V.; Del Rio, M.L.; Rueda, D. Perú: Estrategia Nacional Sobre Diversidad Biológica; Consejo Nacional del Ambiente (CONAM): Lima, Peru, 2001. [Google Scholar]
- Escobal, J.; Aldana, U. Are nontimber forest products the antidote to rainforest degradation? Brazil nut extraction in Madre De Dios, Peru. World Dev. 2003, 31, 1873–1887. [Google Scholar] [CrossRef]
- Scullion, J.J.; Vogt, K.A.; Sienkiewicz, A.; Gmur, S.J.; Trujillo, C. Assessing the influence of land-cover change and conflicting land-use authorizations on ecosystem conversion on the forest frontier of Madre de Dios, Peru. Biol. Conserv. 2014, 171, 247–258. [Google Scholar] [CrossRef]
- Swenson, J.J.; Carter, C.E.; Domec, J.-C.; Delgado, C.I. Gold mining in the Peruvian Amazon: Global prices, deforestation, and mercury imports. PLoS ONE 2011, 6, e18875. [Google Scholar] [CrossRef] [Green Version]
- Puhakka, L.; Salo, M.; Saaksjarvi, I. Bird Diversity, Birdwatching Tourism and Conservation in Peru: A Geographic Analysis. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Duellman, W.E. The Reserva Cuzco Amazonico, Peru: Biological Investigations, Conservation, and Ecotourism; Museum of Natural History, University of Kansas: Lawrence, KS, USA, 1991. [Google Scholar]
- Mori del Aguila, T.; Bendayan Acosta, M.E.; Tresierra Ayala, A.; Mildred, G.D.; Ruiz Sanchez, E.; Bardales Farcia, J.; Reategui Amasifuen, R.; Espinoza Campos, F.; Davila Flores, C. Acomicetes y Basidiomicetes Macroscopicos en Bosques de Puerto Almendras (Loreto, Peru). Folia Amaz. 2011, 20, 7–14. [Google Scholar] [CrossRef]
- Vega Gutiérrez, S.; Robinson, S. Potential of spalting moderate value wood species in Peru. Int. Wood Prod. J. 2015, 6, 165–168. [Google Scholar] [CrossRef]
- Robinson, S.C.; Richter, D.L.; Laks, P.E. Colonization of sugar maple by spalting fungi. For. Prod. J. 2007, 57, 24–32. [Google Scholar]
- Robinson, S.C.; Richter, D.L.; Laks, P.E. Inducing and Stimulating Spalting in Sugar Maple; IRG 08-10652; International Research Group on Wood Protection: Stockholm, Sweden, 2008. [Google Scholar]
- Robinson, S.C.; Weber, G.; Hinsch, E. Inducing zone lines and melanin formation for decorative purposes on North American western wood species, with emphasis on conifers. Int. Wood Prod. J. 2014, 5, 196–199. [Google Scholar] [CrossRef]
- Robinson, S.C.; Richter, D.L.; Laks, P.E. Effects of substrate on laboratory spalting of sugar maple. Holzforschung 2009, 63, 491–495. [Google Scholar] [CrossRef]
- Shigo, A.L.; Sharon, E.M. Mapping columns of discolored and decayed tissues in sugar maple, Acer saccharum. Phytopathology 1970, 60, 232–237. [Google Scholar] [CrossRef]
- Rayner, A.D.M. Interactions between fungi colonising hardwood stumps and their possible role in determining patterns of colinisation and succession. Ann. Appl. Biol. 1978, 89, 131–134. [Google Scholar] [CrossRef]
- Campbell, A.H. Zone lines in plant tissues.1. The black lines formed by Xylaria polymorpha (pers.) grev. in hardwoods. Ann. Appl. Biol. 1933, 20, 123–145. [Google Scholar] [CrossRef]
- Robinson, S.C.; Tudor, D.; Zhang, W.R.; Ng, S.; Cooper, P.A. Ability of three yellow pigment producing fungi to colour wood under controlled conditions. Int. Wood Prod. J. 2014, 5, 103–107. [Google Scholar] [CrossRef]
- Maeda, M.; Yamauchi, T.; Oshima, K.; Shimomura, M.; Miyauchi, S.; Mukae, K.; Sakaki, T.; Shibata, M.; Wakamatsu, K. Extraction of xylindein from Chlorociboria aeruginosa complex and its biological characteristics. Bull. Nagaoka Univ. Technol. 2003, 25, 105–111. [Google Scholar]
- Vega Gutierrez, S.M.; Hazell, K.K.; Simonsen, J.; Robinson, S.C. Description of a Naphthoquinonic Crystal Produced by the Fungus Scytalidium cuboideum. Molecules 2018, 23, 1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Sigler, L.; Lee, J.; Gibas, C.; Yun, S.; Lee, Y. Xylogone ganodermophthora sp. nov., an ascomycetous pathogen causing yellow rot on cultivated mushroom Ganoderma lucidum in Korea. Mycologia 2010, 102, 1167–1184. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, G.M.; Ekong, D.E.; Nielson, A.H.; Todd, L. Xylindein. Chimia 1965, 19, 208–212. [Google Scholar]
- Otterstedt, A. Investigating Green Marquetry on Bowed-String Instruments. The Leaves Be Greene. Galpin Soc. J. 2001, 54, 330–338. [Google Scholar] [CrossRef]
- Blanchette, R.A.; Wilmering, A.M.; Baumeister, M. The use of green-stained wood caused by the fungus Chlorociboria in Intarsia masterpieces from the 15th century. Holzforschung 1992, 46, 225–232. [Google Scholar] [CrossRef]
- Michaelsen, H.; Unger, A.; Fischer, C.-H. Blaugrüne Färbung an Intarsienhölzern des 16. bis 18. Jahrhunderts. Restauro 1992, 98, 17–25. [Google Scholar]
- Gutierrez, P.T.V.; Robinson, S.C. Determining the Presence of Spalted Wood in Spanish Marquetry Woodworks of the 1500s through the 1800s. Coatings 2017, 7, 188. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, D.L. Evaluation of the Retail Market Potential for Locally Produced Paper Birch Lumber in Alaska; USDA: Washington, DC, USA, 2002; pp. 1–19. [Google Scholar]
- Vega Gutierrez, P.T.; Robinson, S.C. Complexity of biodegradation patterns in spalted wood and its influense on the perception of US woodturners. Eur. J. Wood Wood Prod. 2020, 172–183. [Google Scholar] [CrossRef]
- Ramamurthi, C.S.; Korf, R.P.; Batra, L.R. A revision of the North American species of Chlorociboria (Sclerotiniaceae). Mycologia 1958, 49, 854–863. [Google Scholar] [CrossRef]
- Robinson, S.C. Developing fungal pigments for “painting” vascular plants. Appl. Microbiol. Biotechnol. 2012, 93, 1389–1394. [Google Scholar] [CrossRef]
- Vega Gutierrez, S.M. Spalting Fungi: Genetic Identification, Material Interactions and Microscopical Characteristics of Extracted Pigments. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2017. [Google Scholar]
- Rogers, J.D.; Calla, B.E. Xylaria polymorpha and its allies in the continental United States. Mycologia 1986, 78, 391–400. [Google Scholar] [CrossRef]
- Worrall, J.T.; Anagnost, S.E.; Zabel, R.A. Comparison of wood decay among diverse lignicolous fungi. Mycologia 1997, 89, 199–219. [Google Scholar] [CrossRef]
- Phillips, L.W. The nature of spalted wood: Analysis of zone line formation between six white rot fungi. Ph.D. Thesis, Brigham Young University, Provo, UT, USA, 1987. [Google Scholar]
- Robinson, S.C.; Michaelsen, H.; Robinson, J.C. Spalted Wood. The History, Science and Art of a Unique Material, 1st ed.; Schiffer Publishing, Ltd.: Atglen, PA, USA, 2016; p. 287. [Google Scholar]
- Thomas, D.C.; Vandegrift, R.; Ludden, A.; Carroll, G.C.; Roy, B.A. Spatial Ecology of the Fungal Genus Xylaria in a Tropical Cloud Forest. Biotropica 2016, 48, 381–393. [Google Scholar] [CrossRef]
- González, P.; Alaniz, S.; Montelongo, M.; Rauduviniche, L.; Rebellato, J.; Silvera-Pérez, E.; Mondino, P. First report of Pestalotiopsis clavispora causing dieback on blueberry in Uruguay. Plant Dis. 2012, 96, 914. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [Green Version]
- Seaver, F.G. Photographs and descriptions of cup fungi –XXIV. Chlorociboria. Mycologia 1936, 28, 309–394. [Google Scholar]
- Robinson, S.C.; Hinsch, E.; Weber, G.; Freitas, S. Method of extraction and resolubilisation of pigments from Chlorociboria aeruginosa and Scytalidium cuboideum, two prolific spalting fungi. Coloration Technol. 2014, 130, 221–225. [Google Scholar] [CrossRef]
- Weber, G.L.; Boonloed, A.; Naas, K.M.; Koesdjojo, M.T.; Remcho, V.T.; Robinson, S.C. A method to stimulate production of extracellular pigments from wood-degrading fungi using a water carrier. Curr. Res. Environ. Appl. Mycol. 2016, 6, 218–230. [Google Scholar] [CrossRef]
- Robinson, S.C.; Laks, P.E. Wood species affects laboratory colonization rates of Chlorociboria sp. Int. Biodeterior. Biodegrad. 2010, 64, 305–308. [Google Scholar] [CrossRef]
- Etebarian, H.; Scott, E.; Wicks, T. Trichoderma harzianum T39 and T. virens DAR 74290 as Potential Biological Control Agents for Phytophthora erythroseptica. Eur. J. Plant Pathol. 2000, 106, 329–337. [Google Scholar] [CrossRef]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Genetic and metabolic biodiversity of Trichoderma from Colombia and adjacent neotropic regions. Fungal Genet. Biol. 2009, 46, 615–631. [Google Scholar] [CrossRef]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef] [Green Version]
- Chaverri, P.; Castlebury, L.A.; Samuels, G.J.; Geiser, D.M. Multilocus phylogenetic structure within the Trichoderma harzianum/Hypocrea lixii complex. Mol. Phylogenetics Evol. 2003, 27, 302–313. [Google Scholar] [CrossRef]
- Nagia, F.; El-Mohamedy, R.S.R. Dyeing of wool with natural anthraquinone dyes from Fusarium oxysporum. Dye. Pigment. 2007, 75, 550–555. [Google Scholar] [CrossRef]
- Robinson, S.C.; Tudor, D.; Cooper, P.A. Feasibility of using red pigment producing fungi to stain wood for decorative applications. Can. J. For. Res. 2011, 41, 1722–1728. [Google Scholar] [CrossRef]
- Lozovaya, V.V.; Lygin, A.V.; Zernova, O.V.; Li, S.; Widholm, J.M. Lignin degradation by Fusarium solani f. sp. glyines. Plant Dis. 2006, 90, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, S.C.; Laks, P.E.; Richter, D.L. Stimulating spalting in sugar maple using sub-lethal doses of copper. Eur. J. Wood Wood Prod. 2011, 69, 527–532. [Google Scholar] [CrossRef]
- Milus, E.A.; Parson, C.E. Evaluation of foliar fungicides for controlling Fusarium head blight of wheat. Plant Dis. 1994, 78, 697–699. [Google Scholar] [CrossRef]
- Gazis, R.; Rehner, S.; Chaverri, P. Species delimitation in fungal endophyte diversity studies and its implications in ecological and biogeographic inferences. Mol. Ecol. 2011, 20, 3001–3013. [Google Scholar] [CrossRef]
- Herrera, C.S.; Rossman, A.Y.; Samuels, G.J.; Pereira, O.L.; Chaverri, P. Systematics of the Cosmospora viliuscula species complex. Mycologia 2015, 107, 532–557. [Google Scholar] [CrossRef]
- Weber, G.; Chen, H.-L.; Hinsch, E.; Freitas, S.; Robinson, S. Pigments extracted from the wood-staining fungi Chlorociboria aeruginosa, Scytalidium cuboideum, and S. ganodermophthorum show potential for use as textile dyes. Coloration Technol. 2014, 130, 445–452. [Google Scholar] [CrossRef]
- Floudas, D.; Binder, M.; Riley, R.; Barry, K.; Blanchette, R.A.; Henrissat, B.; Martínez, A.T.; Otillar, R.; Spatafora, J.W.; Yadav, J.S. The Paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes. Science 2012, 336, 1715–1719. [Google Scholar] [CrossRef] [Green Version]
- Kotlaba, F. Geographical distribution and ecology of polypores/Polyporales sl. in Czechoslovakia. Academia Praha 1984, 180–184. [Google Scholar]
- Mendil, D.; Uluözlü, Ö.D.; Hasdemir, E.; Çaǧlar, A. Determination of trace elements on some wild edible mushroom samples from Kastamonu, Turkey. Food Chem. 2004, 88, 281–285. [Google Scholar] [CrossRef]
- Hubert, E.E. The Diagnosis of Decay in Wood; US Government Printing Office: Washington, DC, USA, 1925.
- Bannwarth, M.; Bastian, S.; Heckmann-Pohl, D.; Giffhorn, F.; Schulz, G.E. Crystal structure of pyranose 2-oxidase from the white-rot fungus Peniophora sp. Biochemistry 2004, 43, 11683–11690. [Google Scholar] [CrossRef]
- Okamoto, K.; Imashiro, K.; Akizawa, Y.; Onimura, A.; Yoneda, M.; Nitta, Y.; Maekawa, N.; Yanase, H. Production of ethanol by the white-rot basidiomycetes Peniophora cinerea and Trametes suaveolens. Biotechnol. Lett. 2010, 32, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Martin, F. The Ecological Genomics of Fungi; Wiley Blackwell: Ames, IA, USA, 2014. [Google Scholar]
- Sexton, C.M.; Smith, S.M.; Morrell, J.J.; Kropp, B.R.; Corden, M.E.; Graham, R.D. Identity and distribution of Basidiomycotina colonizing Douglas-fir poles during three years of air-seasoning. Mycol. Res. 1992, 96, 321–330. [Google Scholar] [CrossRef]
- Mohali, S.; Burgess, T.; Wingfield, M. Diversity and host association of the tropical tree endophyte Lasiodiplodia theobromae revealed using simple sequence repeat markers. For. Pathol. 2005, 35, 385–396. [Google Scholar] [CrossRef]
- Encinas, O.; Daniel, G. Decay capacity of different strains of the blue stain fungus Lasiodiploida theobromae on various wood species. Mater. Org. 1996, 30, 239–258. [Google Scholar]
- Zabel, R.A.; Morrell, J.J. Wood Microbiology. Decay and Its Prevention; Harcourt Brace Jovanovich, Academic Press, Inc.: New York, NY, USA, 1992. [Google Scholar]
- Louman, B. Silvicultura de Bosques Latifoliados Húmedos con Énfasis en América Central; CATIE: Turrialba, Costa Rica, 2001; Volume 46. [Google Scholar]
- Pennington, T.D.; Reynel, C.; Daza, A. Illustrated Guide to the Trees of Peru; David Hunt: St. Louis, MO, USA, 2004. [Google Scholar]
- Schweingruber, F.H. Tree Species in North America. In Trees and Wood in Dendrochronology; Springer: Berlin/Heidelberg, Germany, 1993; pp. 208–342. [Google Scholar]
- Vega Gutierrez, S.M.; He, Y.; Cao, Y.; Stone, D.; Walsh, Z.; Malhotra, R.; Chen, H.-L.; Chang, C.-H.; Robinson, S.C. Feasibility and Surface Evaluation of the Pigment from Scytalidium cuboideum for Inkjet Printing on Textiles. Coatings 2019, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.C.; Vega Gutierrez, S.M.; Cespedes Garcia, R.A.; Iroume, N.; Vorland, N.R.; McClelland, A.; Huber, M.; Stanton, S. Potential for carrying dyes derived from spalting fungi in natural oils. J. Coat. Technol. Res. 2017, 14, 1107–1113. [Google Scholar] [CrossRef]
- Palomino Agurto, M.E. Wood-Rotting Fungal Pigments as Colorant Coatings on Oil-Based Textile Dyes: A Detailed View of the Interaction between Fungal Pigments and Some Commercial Fabrics. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2018. [Google Scholar]
- Giesbers, G.; Van Schenck, J.; Vega Gutierrez, S.; Robinson, S.; Ostroverkhova, O. Fungi-Derived Pigments for Sustainable Organic (Opto) Electronics. MRS Adv. 2018, 1–6, 2059–8521. [Google Scholar] [CrossRef] [Green Version]
- Vega Gutierrez, S.M.; Kramer, O.E.; Andersen, C.C.; Vega Gutierrez, P.T.; Robinson, S.C. A Method for Citizen Scientists to Catalogue Worldwide Chlorociboria spp. Distribution. Challenges 2018, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Vega Gutierrez, S.M.; Robinson, S.C. Initial investigations into the spalting potential of Peruvian hardwoods. In Proceedings of the IRG45—International Research Group on Wood Protection, St. George, Utah, 11–15 May 2014. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 1990, 18, 315–322. [Google Scholar]
- Tudor, D.; Margaritescu, S.; Sánchez-Ramírez, S.; Robinson, S.C.; Cooper, P.A.; Moncalvo, J.M. Morphological and molecular characterization of the two known North American Chlorociboria species and their anamorphs. Fungal Biol. 2014, 118, 732–742. [Google Scholar] [CrossRef]
- Dugan, K.A.; Lawrence, H.S.; Hares, D.R.; Fisher, C.L.; Budowle, B. An improved method for post-PCR purification for mtDNA sequence analysis. J. Forensic Sci. 2002, 47, 1–8. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Spalting Type Observed | Order | Genus | Species | GenBank Accession Number | GenBank Accession Number-Compare | ID % Similarity |
|---|---|---|---|---|---|---|
| Brown pigment | Incertae sedis | Xylogone | Xylogone sp1. Arx and T. Nilsson | MW340804 | KT264505.1 | 96% |
| Yellow pigment | Helotiales | Scytalidium | Scytalidium sp1. Pesante | MW340805 | HQ631037.1 | 96% |
| Orange pigment | Helotiales | Scytalidium | Scytalidium sp2. Pesante | MW340806 | KR093920.1 | 96% |
| Black pigment | Botryosphaeriales | Lasiodiplodia | Lasiodiplodia theobromae (Pat.) Griffon and Maubl. | MW340807 | KU507483.1 | 100% |
| Purple pigment | Hypocreales | Fusarium | Fusarium solani (Mart.) Sacc. | MW340808 | JX282606.1 | 100% |
| Yellow pigment | Hypocreales | Trichoderma | Trichoderma atrobrunneum F.B. Rocha, P. Chaverri and W. Jaklitsch | MW340833 | FJ442677.1 | 100% |
| Purple pigment | Helotiales | Scytalidium | Scytalidium sp2. Pesante | MW340806 | KR093920.1 | 97% |
| Orange pigment | Xylariales | Neopestalotiopsis | Neopestalotiopsis clavispora (G.F. Atkinson) Maharachch, K.D. Hyde and Crous | MW340817 | KX721071.1 | 100% |
| Lime green pigment | Incertae sedis | Fungal sp1. | MW340811 | KT996091.1 | 100% | |
| Lilac pigment | Incertae sedis | Xylogone | Xylogone sp2. Arx and T. Nilsson | MW340823 | KU512708.1 | 92% |
| Yellow pigment | Incertae sedis | Xylogone | Xylogone sp2. Arx and T. Nilsson | MW340823 | KU512708.1 | 92% |
| Orange pigment | Helotiales | Scytalidium | Scytalidium sp4. Pesante | MW340832 | KR093920.1 | 92% |
| Green and purple pigment | Hypocreales | Fusarium | Fusarium sp. Link | MW340819 | KU950729.1 | 100% |
| Yellow pigment | Leotiomycetes | Leotiomycete | Leotiomycetes sp1. O.E. Erikss. and Winka | MW340814 | KF638554.1 | 100% |
| Purple pigment | Leotiomycetes | Leotiomycete | Leotiomycetes sp2. O.E. Erikss. and Winka | MW340816 | KF638554.1 | 96% |
| Black zone line | Xylariales | Xylaria | Xylaria guianensis (Mont.) Fr. | MW340809 | AM993100.1 | 99% |
| Orange zone line | Incertae sedis | Fungal sp2. | MW340824 | KT996094.1 | 99% | |
| Black zone line | Xylariales | Xylaria | Xylaria hypoxylon (L.) Grev. | MW340812 | KP143687.1 | 99% |
| Orange zone line | Hypocreales | Trichoderma | Trichoderma harzianum Rifai | MW340813 | KC576745.1 | 99% |
| Black zone line | Russulales | Peniophora | Peniophora sp. Cooke | MW340815 | KJ832046.1 | 99% |
| Orange zone line | Hypocreales | Hypocrea | Hypocrea lixii Pat. | MW340829 | FJ442252.1 | 100% |
| Black zone line | Incertae sedis | Fungal sp2. | MW340824 | KT996094.1 | 99% | |
| Brown zone line | Hypocreales | Cosmospora | Cosmospora sp. Rabenh | MW340827 | KJ676175.1 | 100% |
| Black zone line | Xylariales | Xylaria | Xylaria adscendens (Fr.) Fr. | MW340826 | KP133288.1 | 96% |
| Orange zone line | Polyporales | Polyporales sp. Gäum | MW340822 | LN997757.1 | 91% | |
| Black zone line | Hypocreales | Bionectria | Bionectria sp. Speg | MW340821 | HM770964.1 | 99% |
| Black zone line | Xylariales | Neopestalotiopsis | Neopestalotiopsis clavispora (G.F. Atkinson) Maharachch, K.D. Hyde and Crous | MW340831 | KX721071.1 | 100% |
| Black zone line | Incertae sedis | Fugal sp3. | MW340828 | KM265525.1 | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vega Gutierrez, S.M.; Illescas Guevara, J.F.; Andersen, C.C.; Koechlin von Stein, J.; Robinson, S.C. Exploratory Sampling of Spalting Fungi in the Southern Peruvian Amazon Forest. Challenges 2020, 11, 32. https://0-doi-org.brum.beds.ac.uk/10.3390/challe11020032
Vega Gutierrez SM, Illescas Guevara JF, Andersen CC, Koechlin von Stein J, Robinson SC. Exploratory Sampling of Spalting Fungi in the Southern Peruvian Amazon Forest. Challenges. 2020; 11(2):32. https://0-doi-org.brum.beds.ac.uk/10.3390/challe11020032
Chicago/Turabian StyleVega Gutierrez, Sarath M., Javier F. Illescas Guevara, Claudia C. Andersen, Jose Koechlin von Stein, and Seri C. Robinson. 2020. "Exploratory Sampling of Spalting Fungi in the Southern Peruvian Amazon Forest" Challenges 11, no. 2: 32. https://0-doi-org.brum.beds.ac.uk/10.3390/challe11020032