Understanding the Mechanism of Formation of a Response to Juglone for Intact and Immobilized Bacterial Cells as Recognition Elements of Microbial Sensors: Processes Causing the Biosensor Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Microorganism and Culture Conditions
2.2. Preparation of A Set of Standard Solutions of Juglone and Reagents Used
2.3. Formation of Recognition Elements on The Basis of Intact or Immobilized Bacterial Cells
2.4. Determination of The Response to Juglone for Freshly Harvested Intact Cells (Amodel of The Microbial Reactor Sensor) and Immobilized Resting Cells (A Model of The Microbial Membrane Sensor)
2.5. Statistics
3. Results and Discussion
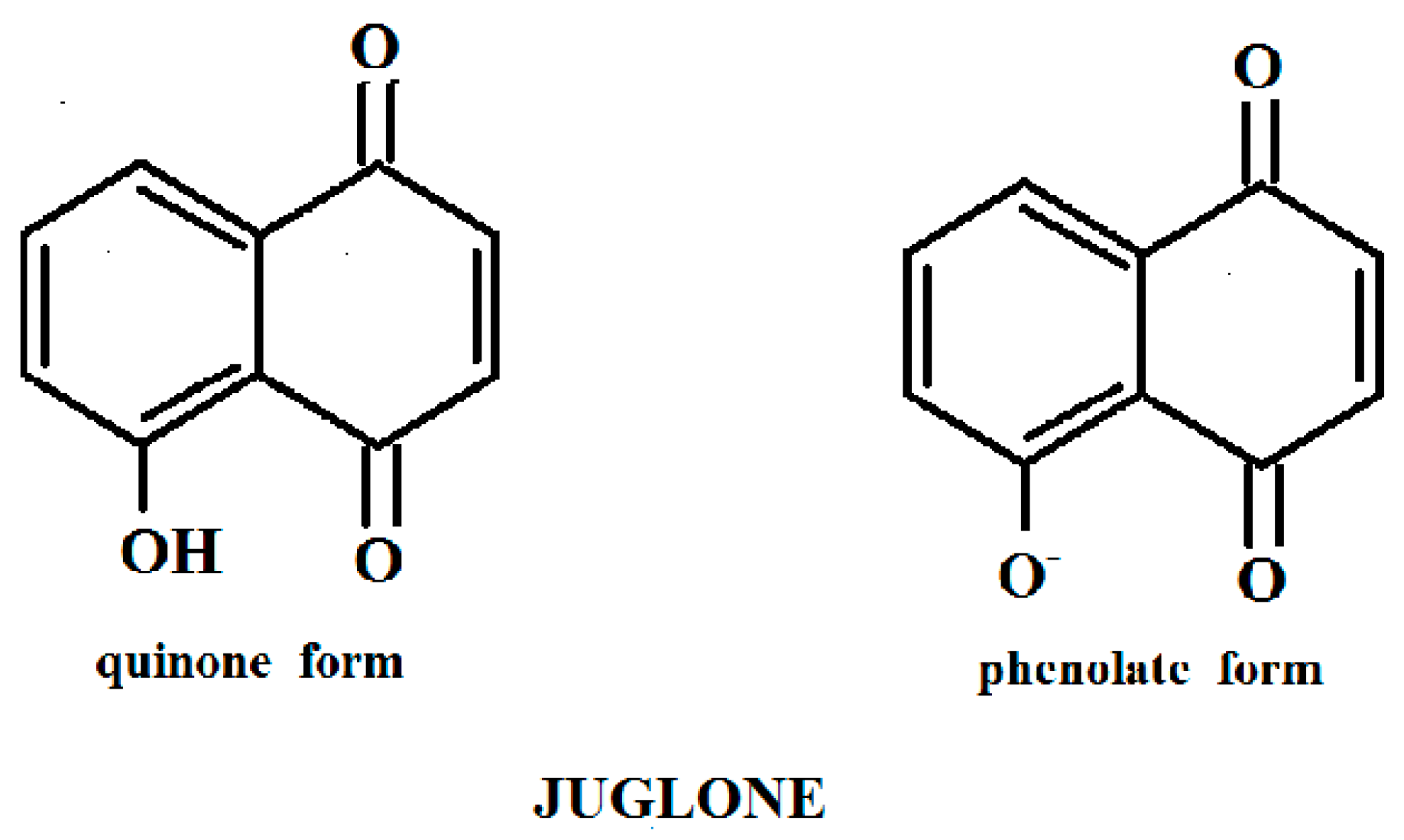
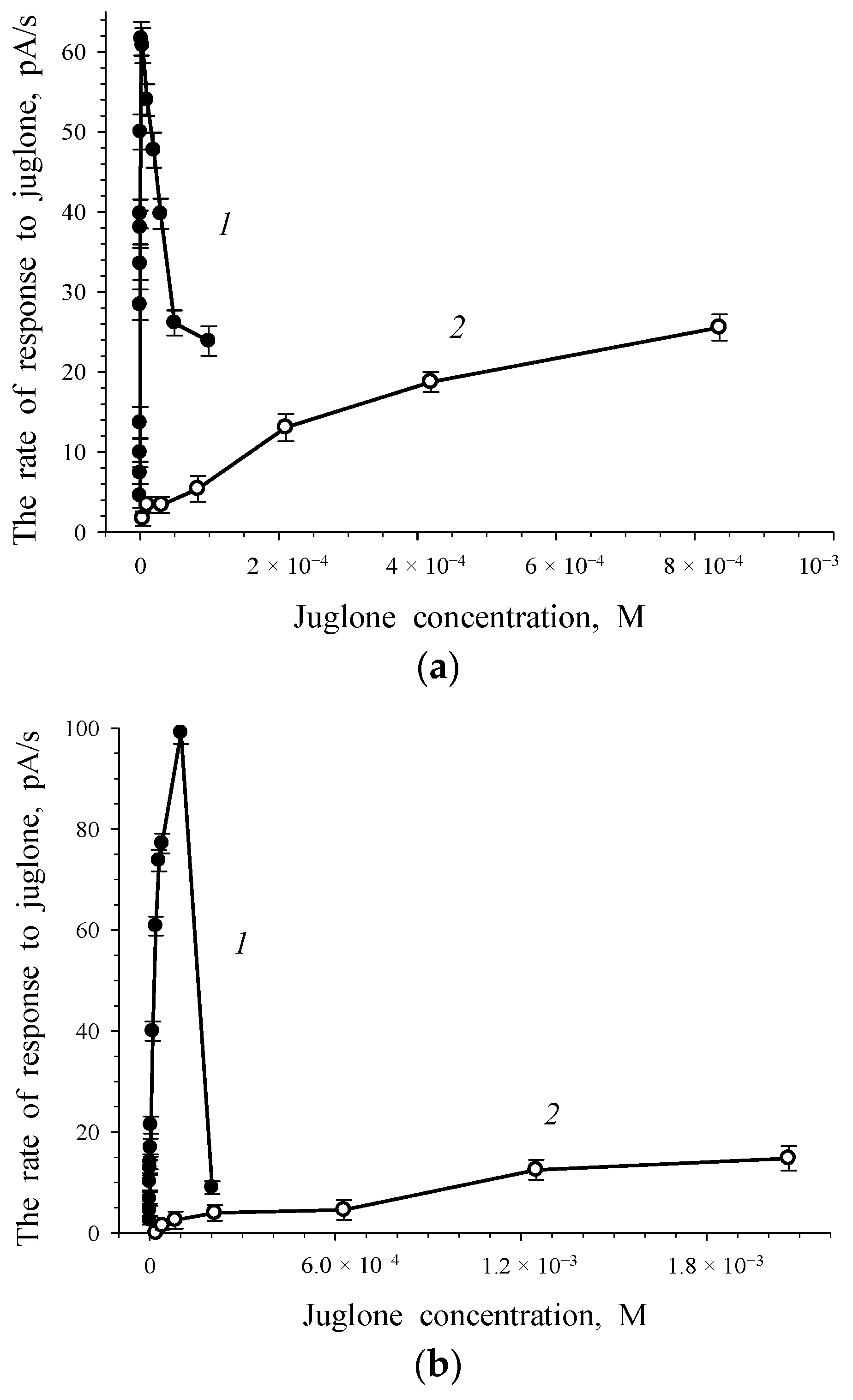
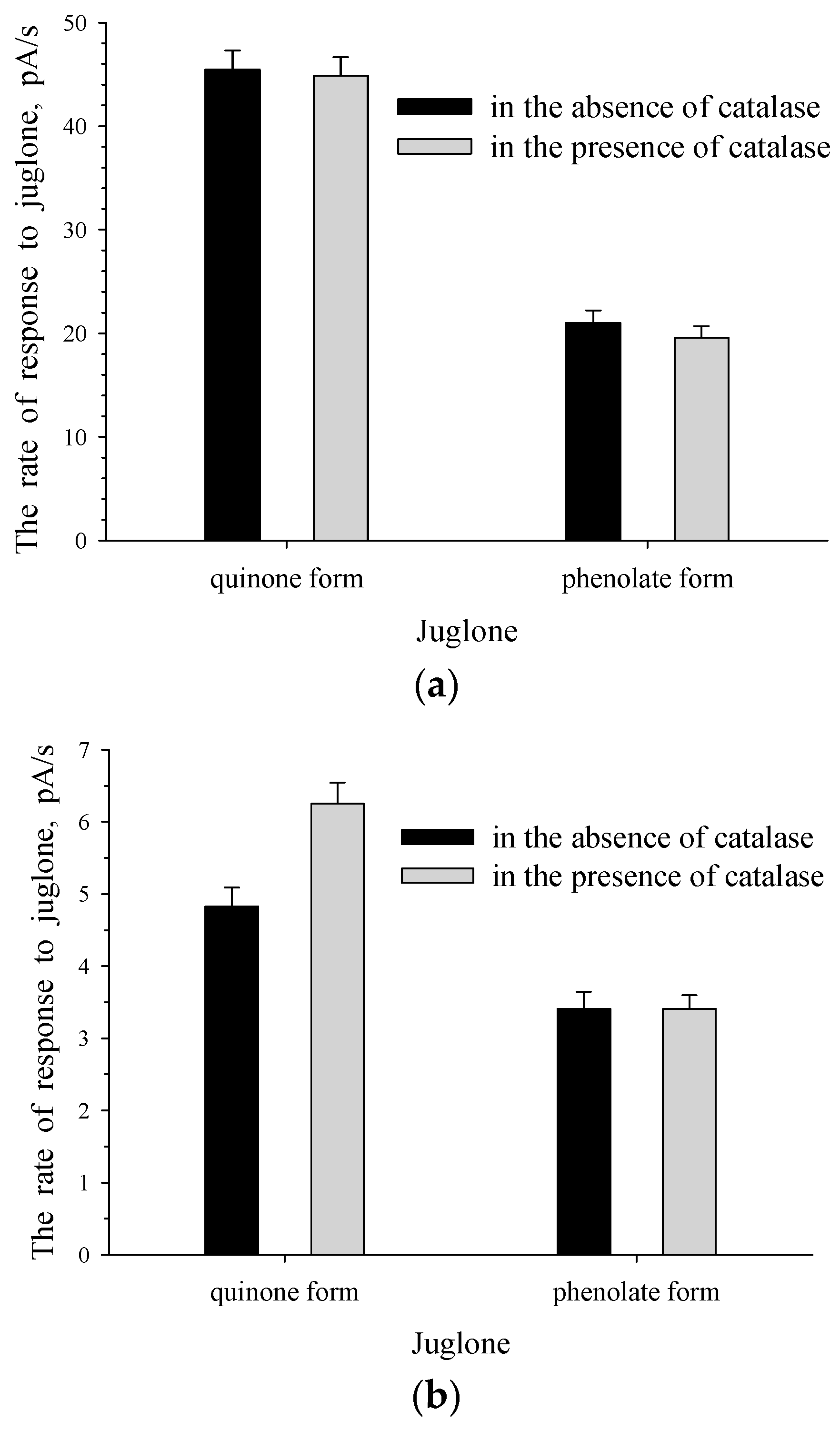
3.1. Response of Intact and Immobilized Cells of Rh. sp. 3 to The Phenolate Form and Quinone Form of Juglone
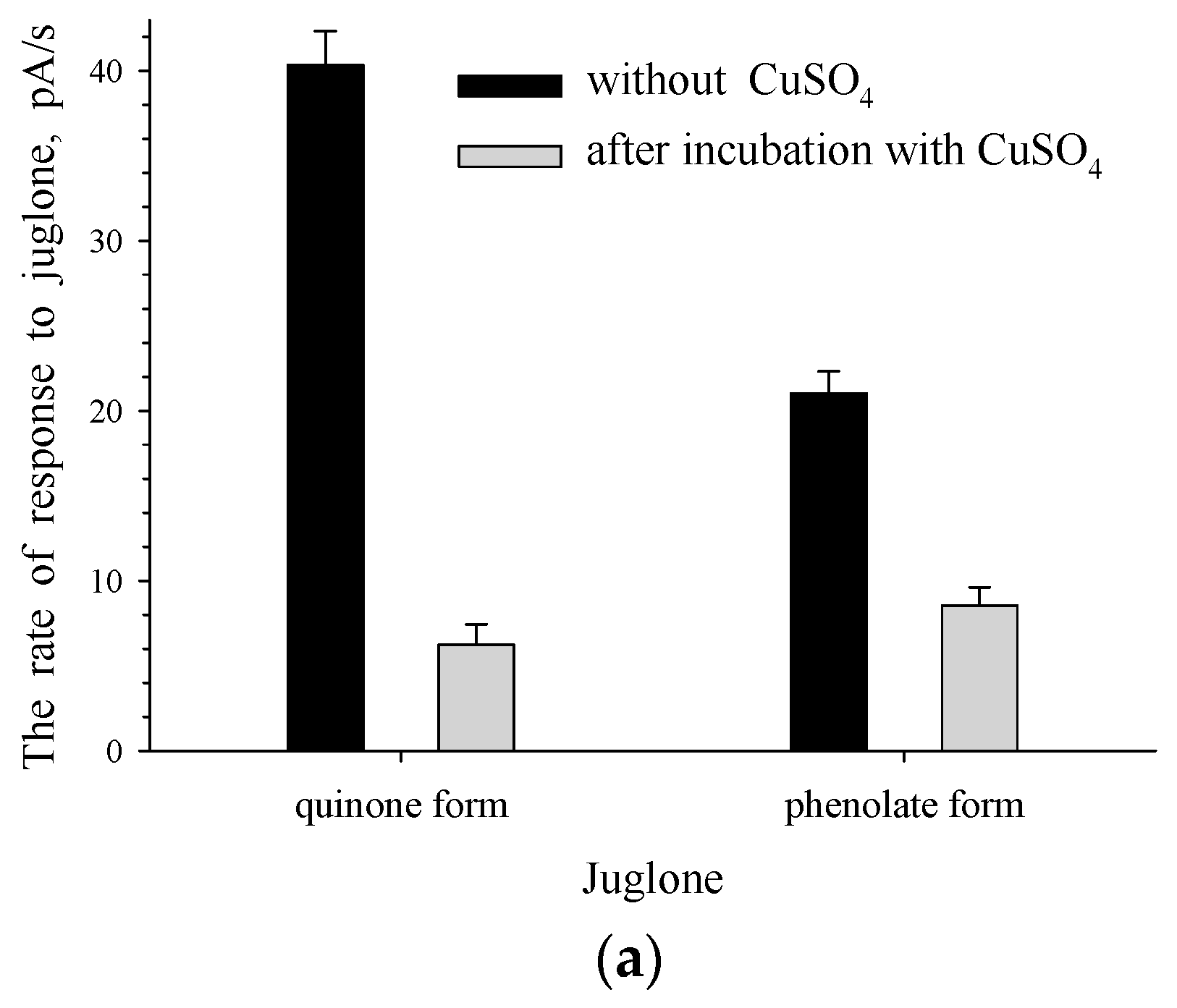
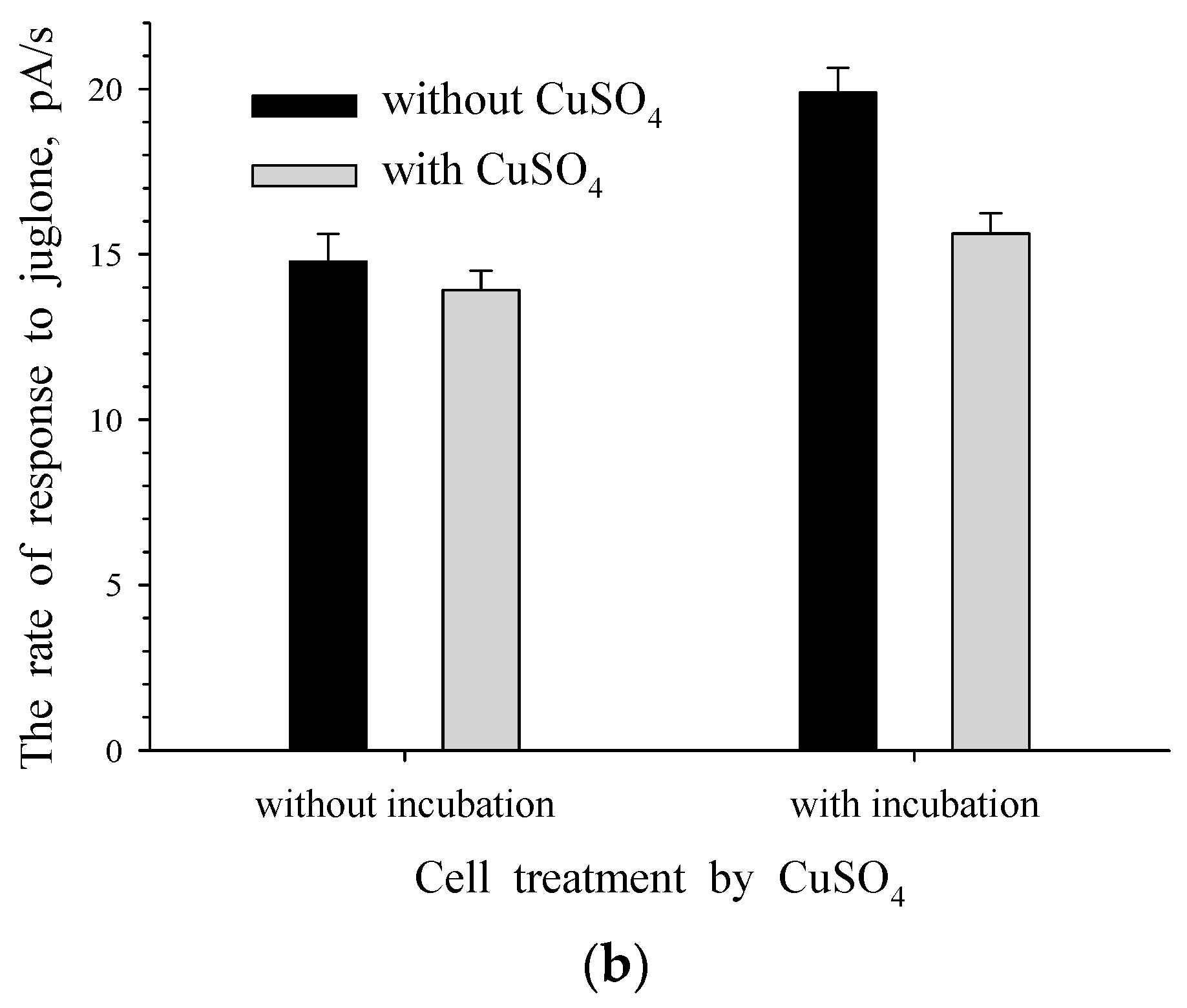
3.2. The Effect of Enzyme-Substrate Interaction on The Formation of Responses to Juglone for Intact and Immobilized Rhodococcus sp. 3 Cells
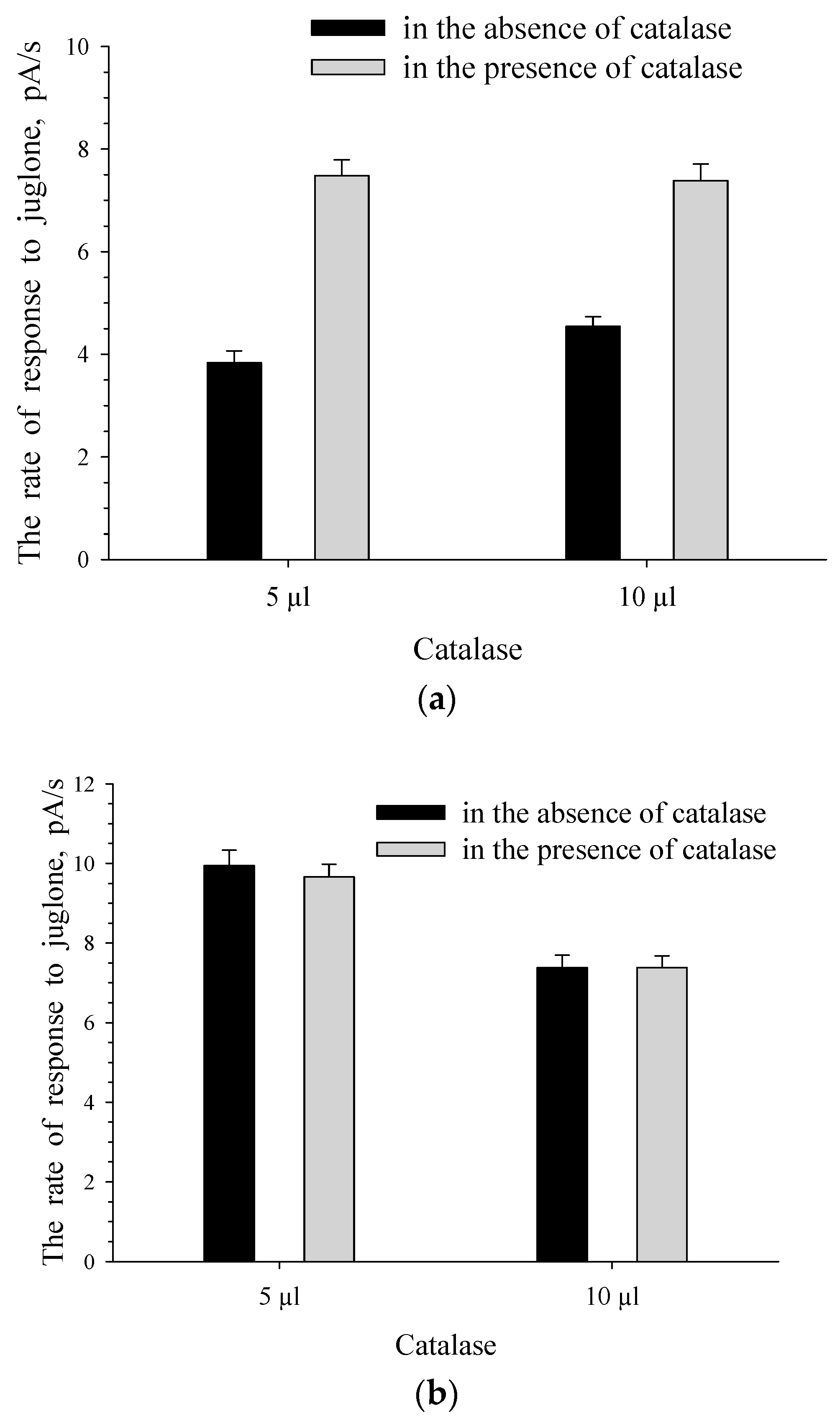
3.3. The Effect of Reactive Oxygen Species (ROS) on The Formation of The Response to Juglone for Intact Cells
3.4. The Effect of ROS on The Formation of The Response to Juglone for Immobilized Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thomson, R.H. Naturally Occurring Quinones, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1971; pp. 198–366. ISBN 978-0-12-689650-3. [Google Scholar]
- Thomson, R.H. Naturally Occurring Quinones IV: Recent Advances, 4th ed.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 141–143. ISBN 978-94-009-1551-0. [Google Scholar]
- Dayronas, J.V.; Zilfikarov, I.N. Natural Naphthoquinones: Perspectives of Medicinal Application; Markhotin, P.Yu.: Schyolkovo, Russia, 2011; p. 29. ISBN 978-5-904456-90-0. (In Russian) [Google Scholar]
- Mayer, F. Chemie der Organischen Farbstoffe: Zweiter Band Natűrliche Organische Farbstoffe; Springer: Berlin/Heidelberg, Germany, 1935; p. 86. ISBN 978-3-642-92508-5. [Google Scholar]
- Rietveld, W.J. Allelopathic effects of juglone on germination and growth of several herbaceous and wood species. J. Chem. Ecol. 1983, 9, 295–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ercisli, S.; Esitken, A.; Turkkal, C.; Orhan, E. The allelopathic effects of juglone and walnut leaf extracts on yield, growth, chemical and PNE compositions of strawberry cv. Fern. Plant Soil Environ. 2005, 51, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Topal, S.; Kocaçalişkan, I.; Arslan, O.; Tel, A.Z. Herbicidal effects of juglone as an allelochemical. Phyton (Horn, Austria) 2007, 46, 259–269. [Google Scholar]
- Krajci, W.M.; Lynch, D.L. The inhibition of various micro-organisms by crude walnut hull extracts and juglone. Microbios. Lett. 1977, 4, 175–181. [Google Scholar]
- Dawson, J.O.; Seymour, P.E. Effect of juglone concentration on growth in vitro of Frankia Ar13 and Rhizobium japonicum strain 71. J. Chem. Ecol. 1983, 9, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, Y.; Wu, R.; Jiang, D.; Bai, B.; Tan, D.; Yan, T.; Sun, X.; Zhang, Q.; Wu, Z. Antibacterial activity of juglone against Staphylococcus aureus: From apparent to proteomic. Int. J. Mol. Sci. 2016, 17, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, A. Juglone: A therapeutic phytochemical from Juglans regia L. J. Med. Plants Res. 2011, 5, 5324–5330. [Google Scholar]
- Ilicheva, E.S.; Miniakhmetova, E.R.; Safina, R.I.; Krainov, A.S. Main methods of the production of 5-hydroxy-1,4-naphthoquinone (juglone)-an antibacterial preparation with the wide range of activities. Bulletin Kazan Tech. Univ. 2015, 18, 147–150. (In Russian) [Google Scholar]
- Fang, F.; Qin, Y.; Qi, L.; Fang, Q.; Zhao, L.; Chen, S.; Li, Q.; Zhang, D.; Wang, L. Juglone exerts antitumor effect in ovarian cancer cells. Iran. J. Basic Med. Sci. 2015, 18, 544–548. [Google Scholar] [PubMed]
- Wu, J.; Zhang, H.; Xu, Y.; Zhang, J.; Zhu, W.; Zhang, Y.; Chen, L.; Hua, W.; Mao, Y. Juglone induced apoptosis of tumor stem-like cells through ROS-p38 pathway in glioblastoma. BMC Neurol. 2017, 17, 70. [Google Scholar] [CrossRef] [Green Version]
- Kurtyka, R.; Pokora, W.; Tukaj, Z.; Karcz, W. Effect of juglone and lawsone on oxidative stress in maize coleoptile cells treated with IAA. AoB Plants 2016, 8, plw073. [Google Scholar] [CrossRef] [Green Version]
- Medentsev, A.G.; Arinbasarova, A.Yu.; Akimenko, V.K. Adaptation of the Phytopathogenic Fungus Fusarium decemcellulare to Oxidative Stress. Microbiology 2001, 70, 26–30. [Google Scholar] [CrossRef]
- Biryukova, E.N.; Medentsev, A.G.; Arinbasarova, A.Yu.; Akimenko, V.K. Tolerance of the Yeast Yarrowia lipolytica to Oxidative Stress. Microbiology 2006, 75, 243–247. [Google Scholar] [CrossRef]
- Solyanikova, I.P.; Suzina, N.E.; Emelyanova, E.V.; Polivtseva, V.N.; Pshenichnikova, A.B.; Lobanok, A.G.; Golovleva, L.A. Morphological, physiological, and biochemical characteristics of a benzoate-degrading strain Rhodococcus opacus 1CP under stress conditions. Microbiology 2017, 86, 202–212. [Google Scholar] [CrossRef]
- Ahmad, T.; Suzuki, Y.J. Juglone in Oxidative Stress and Cell Signaling. Antioxidants 2019, 8, 91. [Google Scholar] [CrossRef] [Green Version]
- Rettenmaier, H.; Lingens, F. Purification and some properties of two isofunctional juglone hydroxylases from Pseudomonas putida J1. Biol. Chem. Hoppe-Seyler 1985, 366, 637–646. [Google Scholar] [CrossRef]
- Wessendorf, J.; Rettenmaier, H.; Lingens, F. Degradation of lawsone by Pseudomonas putida L2. Biol. Chem. Hoppe-Seyler 1985, 366, 945–951. [Google Scholar] [CrossRef]
- Muller, U.; Lingens, F. Degradation of 1,4-naphthoquinones by Pseudomonas putida. Biol. Chem. Hoppe-Seyler 1988, 369, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, D.; Schomburg, I.; Chang, A. Handbook of Enzyme. Vol. 27. Class, I. Oxidoreductases XII. EC 1.14.15-1.97; Springer: Berlin/Heidelberg, Germany, 2006; pp. 364–366. ISBN 3-540-30439-8. [Google Scholar]
- Emelyanova, E.V.; Reshetilov, A.N.; Solyanikova, I.P. Amperometric Rhodococcus biosensor for studies concerning juglone metabolism. Proceedings. Book II. In Proceedings of the PHYSICS AND RADIOELECTRONICS IN MEDICINE AND ECOLOGY-ΦPЭMЭ’2018, Vladimir-Suzdal, Russia, 3–5 July 2018; pp. 304–308, ISBN 978-5-905527-27-2. (Abstract in English). [Google Scholar]
- Emelyanova, E.V.; Solyanikova, I.P. Study of the effect of juglone on intact and immobilized microbial cells. In Proceedings of the MECHANISMS FACILITATING PLANT AND MICROORGANISM TOLERANCE IN HOSTILE ENVIRONMENTS, Irkutsk, Russia, 10–15 July; Publishing office of V.B. Sochava Institute of Geography, Siberian Branch of the Russian Academy of Sciences: Irkutsk, Russia, 2018. Part I. pp. 297–301, (Abstract in English). [Google Scholar] [CrossRef] [Green Version]
- Turner, A.P.F.; Karube, I.; Wilson, G.S. Biosensors: Fundamentals and Applications; Oxford University Press: New York, NY, USA, 1987; ISBN 0198547242. [Google Scholar]
- Emelyanova, E.V.; Solyanikova, I.P. Evaluation of phenol-degradation activity of Rhodococcus opacus 1CP using immobilized and intact cells. Int. J. Environ. Sci. Technol. 2020, 17, 2279–2294. [Google Scholar] [CrossRef]
- Emelyanova, E.V.; Solyanikova, I.P. Evaluation of 3-chlorobenzoate 1,2-dioxygenase inhibition by 2- and 4-chlorobenzoate with a cell-based technique. Biosensors 2019, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Qu, Z.; Zou, X.; Ji, C. Effect of Juglone on ROS Production and Microchondrial Transmembrane Potential in SGC-7901 Cells. In Information Technology and Agricultural Engineering. Advances in Intelligent and Soft Computing; Zhu, E., Sambath, S., Eds.; Springer: Berlin, Germany, 2012; Volume 134, pp. 7–14. ISBN 978-3-642-27536-4. [Google Scholar]
- Jha, B.K.; Jung, H.J.; Seo, I.; Suh, M.H.; Baek, W.K. Juglone induces cell death of Acanthamoeba through increased production of reactive oxygen species. Exp. Parasitol. 2015, 159, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Hirsch, O.; Mohsen, M.; Samuni, A. Effect of nitroxide stable radicals on juglone cytotoxicity. Arch. Biochem. Biophys. 1994, 312, 385–391. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emelyanova, E.V.; Solyanikova, I.P. Understanding the Mechanism of Formation of a Response to Juglone for Intact and Immobilized Bacterial Cells as Recognition Elements of Microbial Sensors: Processes Causing the Biosensor Response. Biosensors 2021, 11, 56. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11020056
Emelyanova EV, Solyanikova IP. Understanding the Mechanism of Formation of a Response to Juglone for Intact and Immobilized Bacterial Cells as Recognition Elements of Microbial Sensors: Processes Causing the Biosensor Response. Biosensors. 2021; 11(2):56. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11020056
Chicago/Turabian StyleEmelyanova, Elena V., and Inna P. Solyanikova. 2021. "Understanding the Mechanism of Formation of a Response to Juglone for Intact and Immobilized Bacterial Cells as Recognition Elements of Microbial Sensors: Processes Causing the Biosensor Response" Biosensors 11, no. 2: 56. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11020056