Spectrophotometric Assays for Sensing Tyrosinase Activity and Their Applications

Abstract
:1. Introduction
2. Biochemical Characteristics of TYR
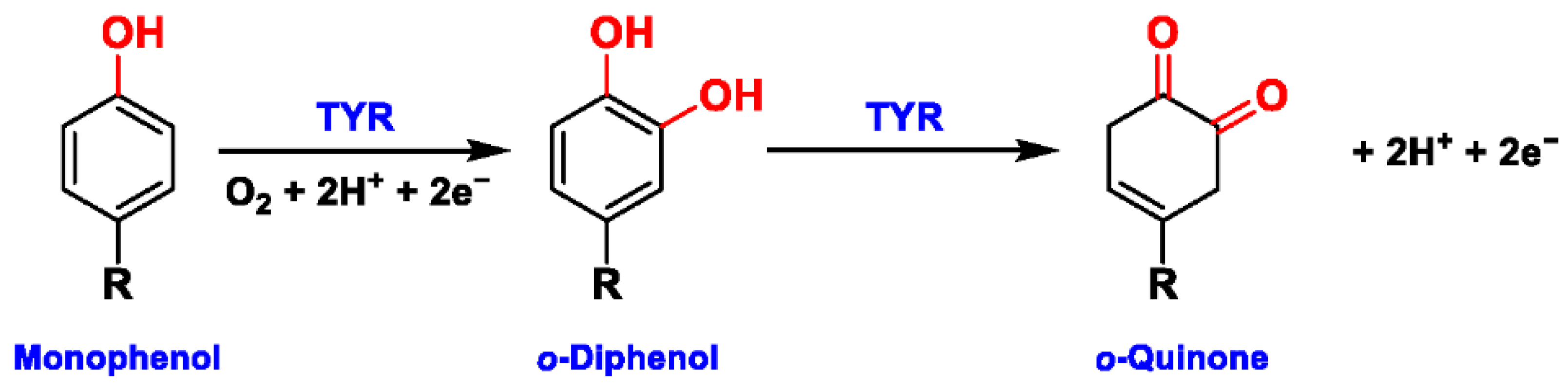
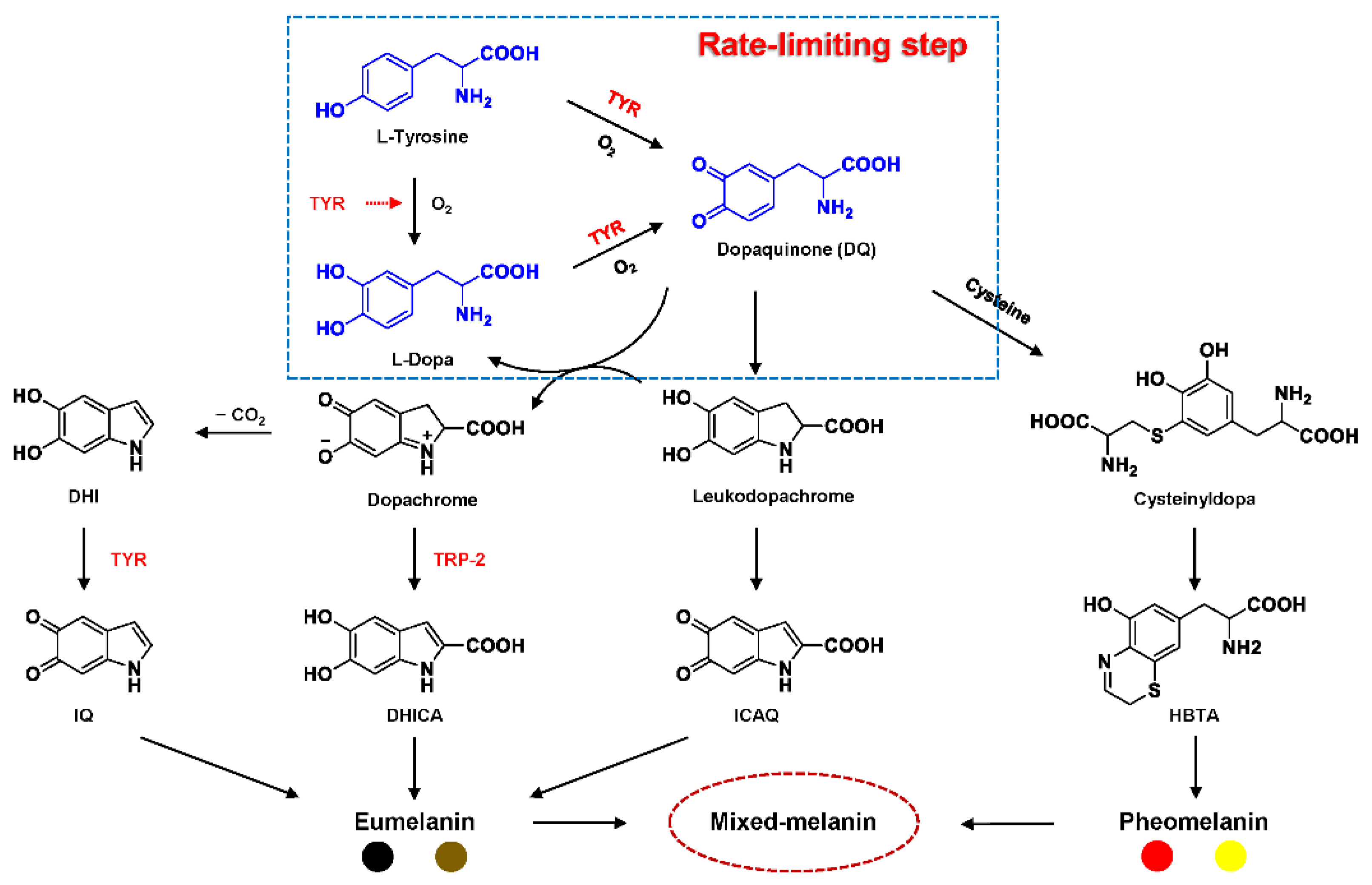
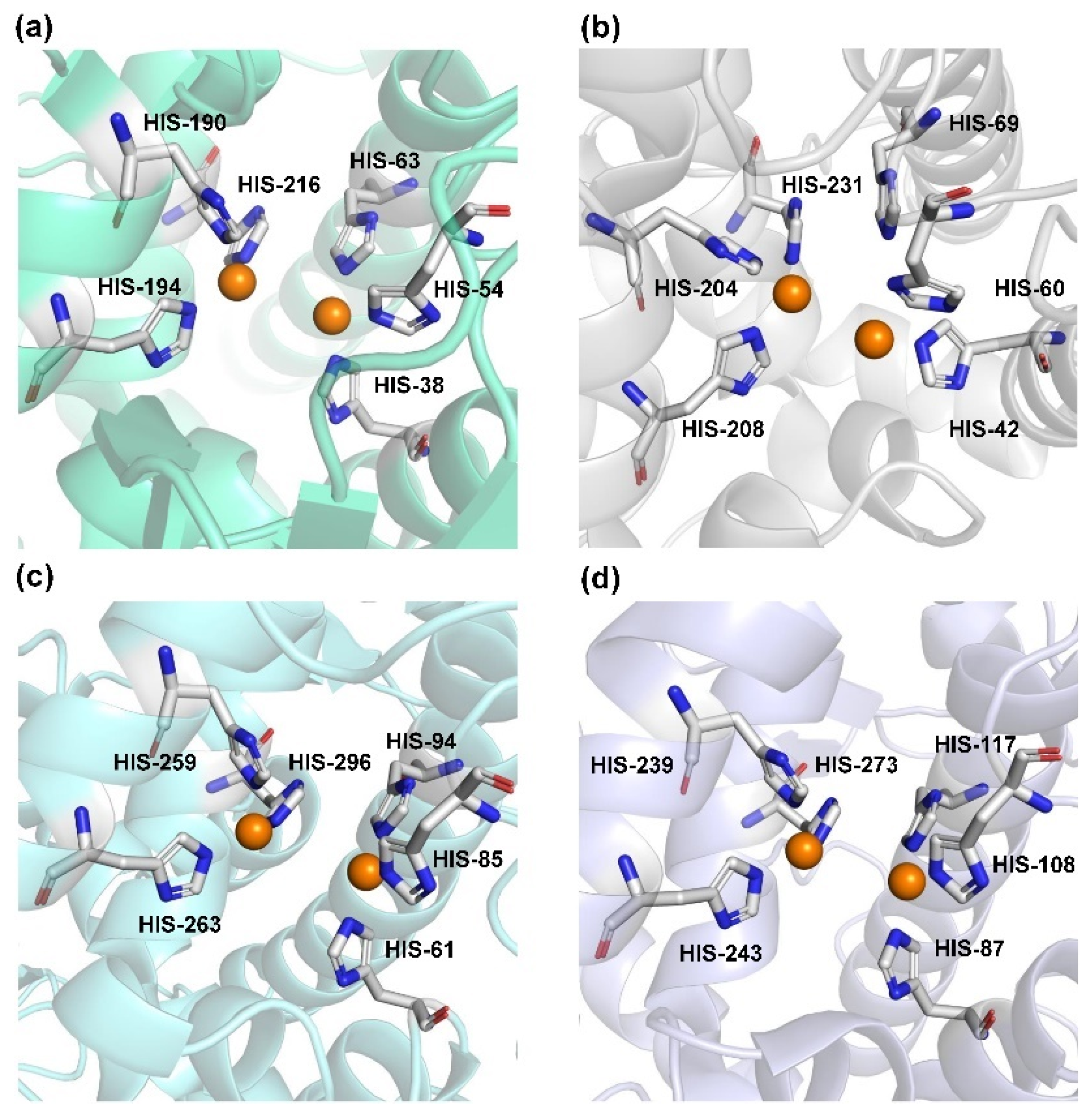
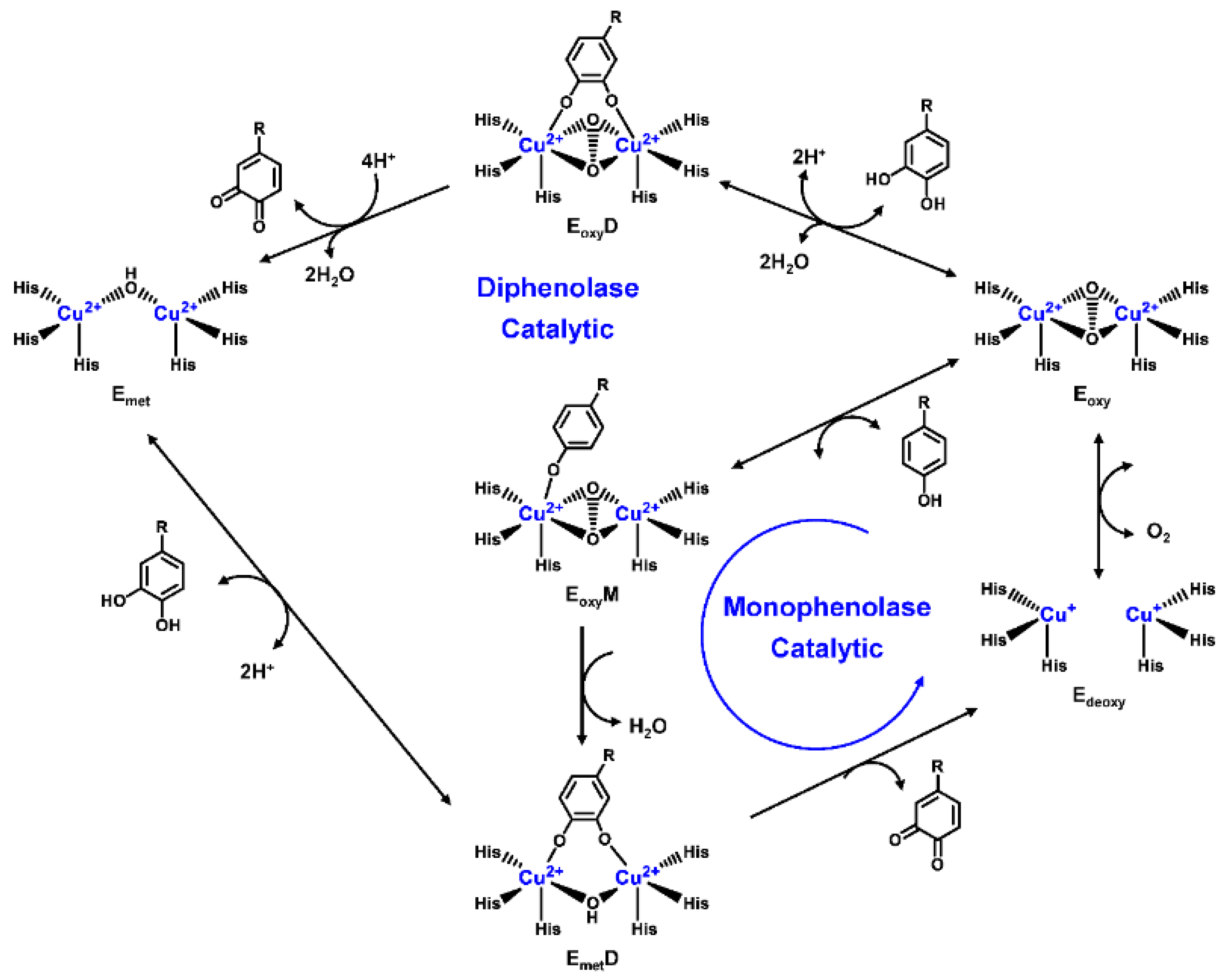
2.1. Structural Feature and Catalytic Mechanism of TYR
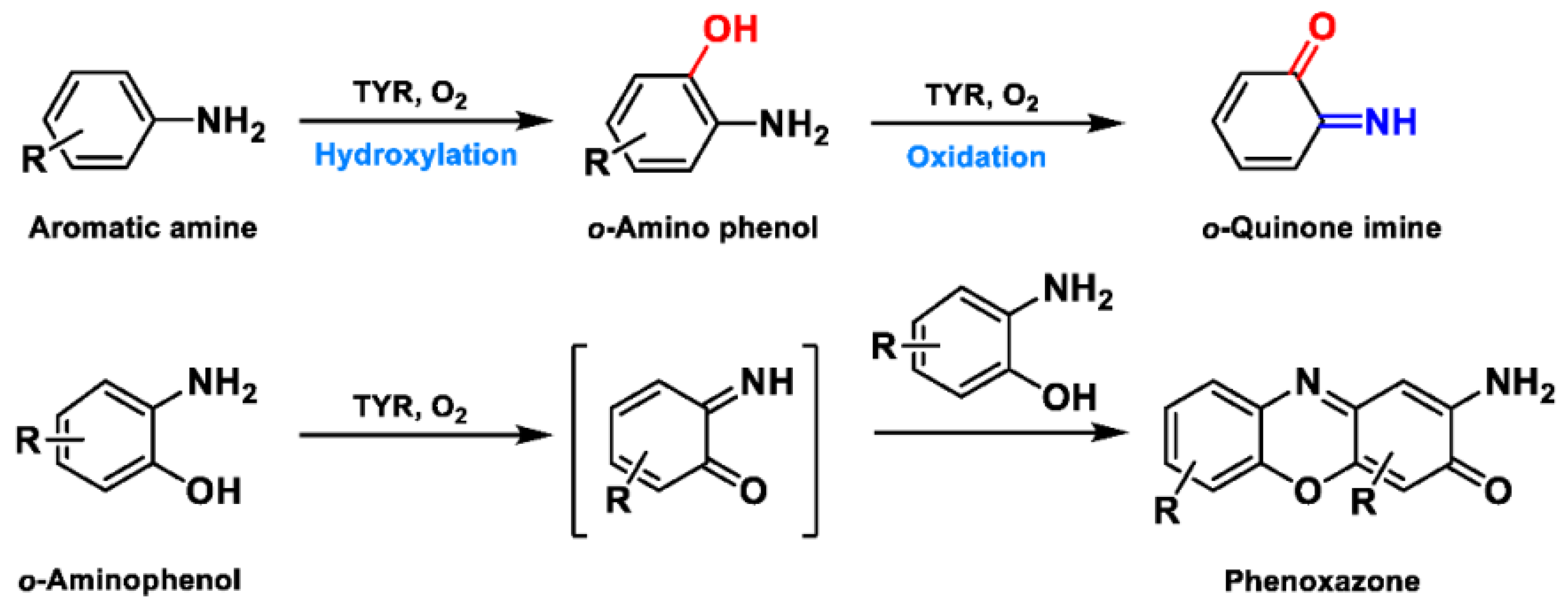
2.2. Substrate Specificity of TYR
3. Optical Assays for Sensing TYR Activity
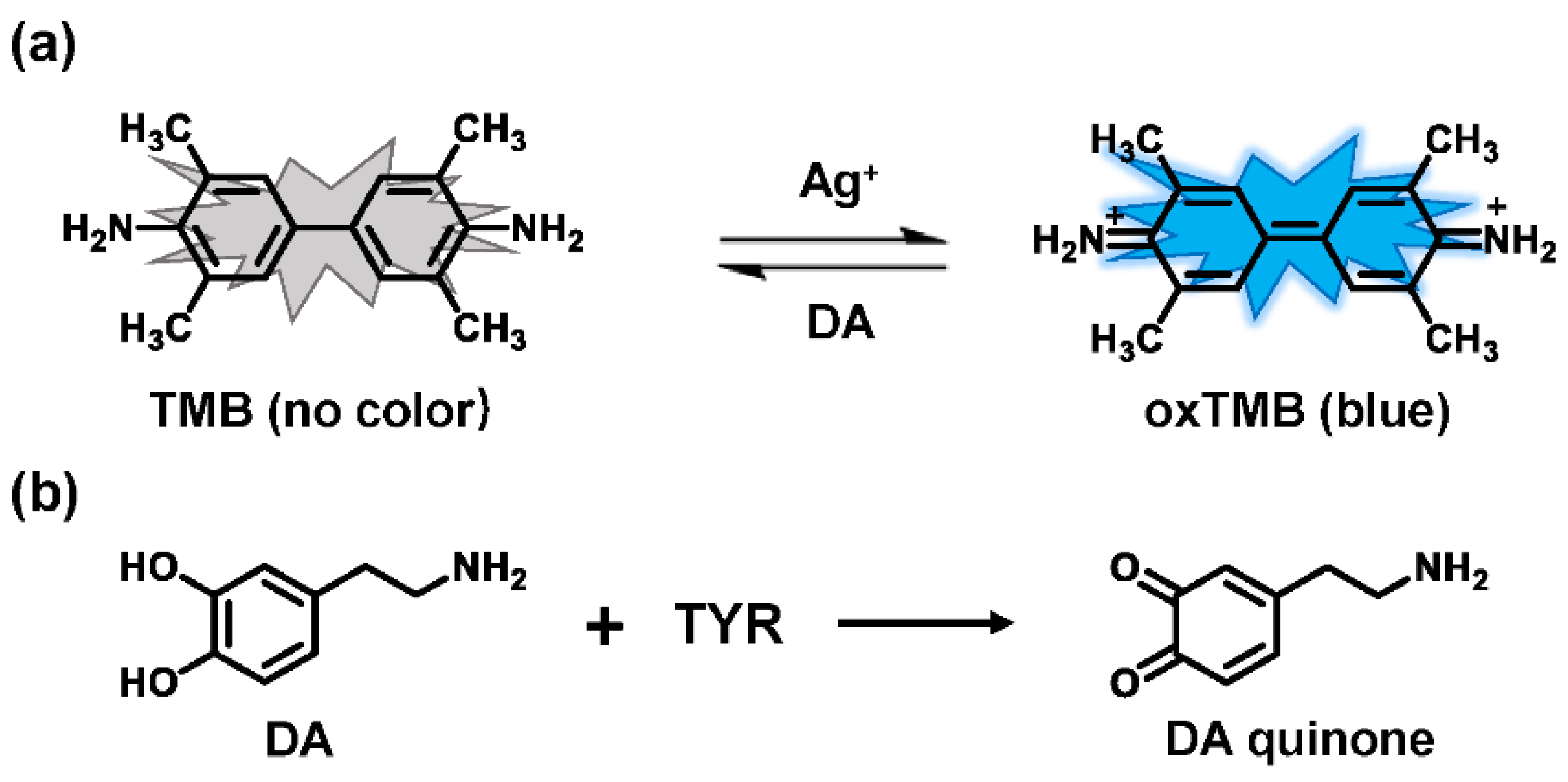
3.1. Spectrophotometric Assays
3.2. Fluorometric Assays
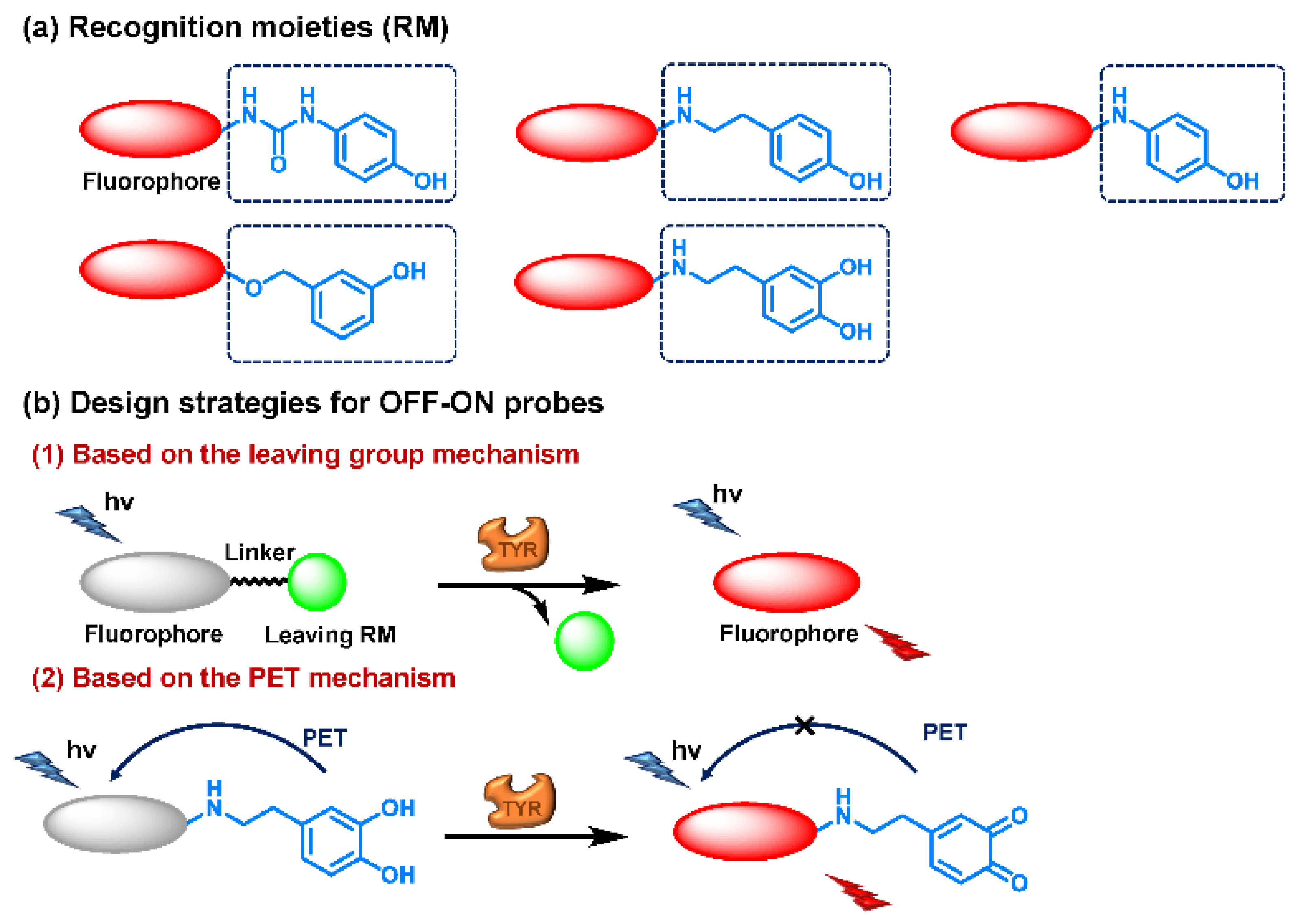
3.2.1. Small Molecule-Based Fluorescent Substrates
4-Hydroxyphenyl Recognition Units
3-Hydroxyphenyl Recognition Units
3.2.2. Nanoparticle-Based Fluorescent Probes
4. Biomedical Applications of TYR Activity Assays
4.1. Sensing and Imaging TYR Activities in Biological Systems
4.2. Screening and Characterization of TYR Inhibitors
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lai, X.; Wichers, H.J.; Soler-Lopez, M.; Dijkstra, B.W. Structure and Function of Human Tyrosinase and Tyrosinase-Related Proteins. Chemistry 2018, 24, 47–55. [Google Scholar] [CrossRef]
- Min, K.; Park, G.W.; Yoo, Y.J.; Lee, J.S. A perspective on the biotechnological applications of the versatile tyrosinase. Bioresour. Technol. 2019, 289, 121730. [Google Scholar] [CrossRef] [PubMed]
- Olivares, C.; Solano, F. New insights into the active site structure and catalytic mechanism of tyrosinase and its related proteins. Pigment. Cell Melanoma Res. 2009, 22, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Baek, K.; Lee, J.E.; Kim, B.G. Using tyrosinase as a monophenol monooxygenase: A combined strategy for effective inhibition of melanin formation. Biotechnol. Bioeng. 2016, 113, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Jung, S.H. Downregulation of melanogenesis: Drug discovery and therapeutic options. Drug Discov. Today 2017, 22, 282–298. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyar, T.; Namasivayam, V.; Manickam, M.; Jung, S.H. Inhibitors of Melanogenesis: An Updated Review. J. Med. Chem. 2018, 61, 7395–7418. [Google Scholar] [CrossRef]
- Lee, S.Y.; Baek, N.; Nam, T.G. Natural, semisynthetic and synthetic tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2016, 31, 1–13. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Hearing, V.J.; Tsukamoto, K. Enzymatic control of pigmentation in mammals. FASEB J. 1991, 5, 2902–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, H.; Ichihashi, M.; Hearing, V.J. Role of the ubiquitin proteasome system in regulating skin pigmentation. Int. J. Mol. Sci. 2009, 10, 4428–4434. [Google Scholar] [CrossRef] [Green Version]
- Costin, G.E.; Hearing, V.J. Human skin pigmentation: Melanocytes modulate skin color in response to stress. FASEB J. 2007, 21, 976–994. [Google Scholar] [CrossRef]
- Ray, K.; Chaki, M.; Sengupta, M. Tyrosinase and ocular diseases: Some novel thoughts on the molecular basis of oculocutaneous albinism type 1. Prog. Retin. Eye Res. 2007, 26, 323–358. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, V.I.F.; Lima, M.D.F.; Sofia, M. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar]
- Ghanem, G.; Fabrice, J. Tyrosinase related protein 1 (TYRP1/gp75) in human cutaneous melanoma. Mol. Oncol. 2011, 5, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Gradilone, A.; Cigna, E.; Agliano, A.M.; Frati, L. Tyrosinase Expression as a Molecular Marker for Investigating the Presence of Circulating Tumor Cells in Melanoma Patients. Curr. Cancer Drug Tar. 2010, 10, 529–538. [Google Scholar] [CrossRef]
- Jawaid, S.; Khan, T.H.; Osborn, H.M.; Williams, N.A. Tyrosinase activated melanoma prodrugs. Anticancer. Agents Med. Chem. 2009, 9, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Carballo-Carbajal, I.; Laguna, A.; Romero-Gimenez, J.; Cuadros, T.; Bove, J.; Martinez-Vicente, M.; Parent, A.; Gonzalez-Sepulveda, M.; Penuelas, N.; Torra, A.; et al. Brain tyrosinase overexpression implicates age-dependent neuromelanin production in Parkinson’s disease pathogenesis. Nat. Commun. 2019, 10, 973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, G.F.; Ming, L.J. Alzheimer’s disease related copper(II)- beta-amyloid peptide exhibits phenol monooxygenase and catechol oxidase activities. Angew. Chem. Int. Ed. Engl. 2005, 44, 5501–5504. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T. Tyrosinase-expressing neuronal cell line as in vitro model of Parkinson’s disease. Int. J. Mol. Sci. 2010, 11, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Y.; Wang, P.; Zhao, D.F.; Gonzalez, F.J.; Fan, Y.F.; Xia, Y.L.; Ge, G.B.; Yang, L. Analytical methodologies for sensing catechol-O-methyltransferase activity and their applications. J. Pharm. Anal. 2021, 11, 15–27. [Google Scholar] [CrossRef]
- Cordes, P.; Sun, W.; Wolber, R.; Kolbe, L.; Klebe, G.; Rohm, K.H. Expression in non-melanogenic systems and purification of soluble variants of human tyrosinase. Biol. Chem. 2013, 394, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Dolinska, M.B.; Wingfield, P.T.; Sergeev, Y.V. Purification of Recombinant Human Tyrosinase from Insect Larvae Infected with the Baculovirus Vector. Curr. Protoc. Protein. Sci. 2017, 89, 6.15.1–6.15.12. [Google Scholar]
- Matoba, Y.; Kumagai, T.; Yamamoto, A.; Yoshitsu, H.; Sugiyama, M. Crystallographic evidence that the dinuclear copper center of tyrosinase is flexible during catalysis. J. Biol. Chem. 2006, 281, 8981–8990. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Uyama, H. Tyrosinase inhibitors from natural and synthetic sources: Structure, inhibition mechanism and perspective for the future. Cell Mol. Life Sci. 2005, 62, 1707–1723. [Google Scholar] [CrossRef]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismaya, W.T.; Rozeboom, H.J.; Weijn, A.; Mes, J.J.; Fusetti, F.; Wichers, H.J.; Dijkstra, B.W. Crystal structure of Agaricus bisporus mushroom tyrosinase: Identity of the tetramer subunits and interaction with tropolone. Biochemistry 2011, 50, 5477–5486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ionita, E.; Aprodu, I.; Stanciuc, N.; Rapeanu, G.; Bahrim, G. Advances in structure-function relationships of tyrosinase from Agaricus bisporus-investigation on heat-induced conformational changes. Food Chem. 2014, 156, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Sendovski, M.; Kanteev, M.; Ben-Yosef, V.S.; Adir, N.; Fishman, A. First structures of an active bacterial tyrosinase reveal copper plasticity. J. Mol. Biol. 2011, 405, 227–237. [Google Scholar] [CrossRef]
- Bijelic, A.; Pretzler, M.; Molitor, C.; Zekiri, F.; Rompel, A. The Structure of a Plant Tyrosinase from Walnut Leaves Reveals the Importance of “Substrate-Guiding Residues” for Enzymatic Specificity. Angew. Chem. Int. Ed. Engl. 2015, 54, 14677–14680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenoll, L.G.; Peñalver, M.J.; Rodríguez-López, J.N.; Varón, R.; García-Cánovas, F.; Tudela, J. Tyrosinase kinetics: Discrimination between two models to explain the oxidation mechanism of monophenol and diphenol substrates. Int. J. Biochem. Cell Biol. 2004, 36, 235–246. [Google Scholar] [CrossRef]
- Fenoll, L.G.; Rodriguez-Lopez, J.N.; Garcia-Sevilla, F.; Garcia-Ruiz, P.A.; Varon, R.; Garcia-Canovas, F.; Tudela, J. Analysis and interpretation of the action mechanism of mushroom tyrosinase on monophenols and diphenols generating highly unstable o-quinones. Biochim. Biophys. Acta 2001, 1548, 1–22. [Google Scholar] [CrossRef]
- Sanchez-Ferrer, A.; Rodriguez-Lopez, J.N.; Garcia-Canovas, F.; Garcia-Carmona, F. Tyrosinase: A comprehensive review of its mechanism. Biochim. Biophys. Acta 1995, 1247, 1–11. [Google Scholar] [CrossRef]
- Qu, Y.; Zhan, Q.; Du, S.; Ding, Y.; Fang, B.; Du, W.; Wu, Q.; Yu, H.; Li, L.; Huang, W. Catalysis-based specific detection and inhibition of tyrosinase and their application. J. Pharm. Anal. 2020, 10, 414–425. [Google Scholar] [CrossRef]
- Rescigno, A.; Sollai, F.; Pisu, B.; Rinaldi, A.; Sanjust, E. Tyrosinase inhibition: General and applied aspects. J. Enzyme Inhib. Med. Chem. 2002, 17, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjust, E.; Cecchini, G.; Sollai, F.; Curreli, N.; Rescigno, A. 3-Hydroxykynurenine as a substrate/activator for mushroom tyrosinase. Arch. Biochem. Biophys 2003, 412, 272–278. [Google Scholar] [CrossRef]
- McLarin, M.A.; Leung, I.K.H. Substrate specificity of polyphenol oxidase. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 274–308. [Google Scholar] [CrossRef]
- Gasowska, B.; Kafarski, P.; Wojtasek, H. Interaction of mushroom tyrosinase with aromatic amines, o-diamines and o-aminophenols. Biochim. Biophys. Acta 2004, 1673, 170–177. [Google Scholar] [CrossRef]
- Rescigno, A.; Bruyneel, F.; Padiglia, A.; Sollai, F.; Salis, A.; Marchand-Brynaert, J.; Sanjust, E. Structure-activity relationships of various amino-hydroxy-benzenesulfonic acids and sulfonamides as tyrosinase substrates. Biochim. Biophys. Acta 2011, 1810, 799–807. [Google Scholar] [CrossRef]
- Espin, J.C.; Garcia-Ruiz, P.A.; Tudela, J.; Garcia-Canovas, F. Study of stereospecificity in mushroom tyrosinase. Biochem. J. 1998, 331, 547–551. [Google Scholar] [CrossRef] [Green Version]
- Espin, J.C.; Varon, R.; Fenoll, L.G.; Gilabert, M.A.; Garcia-Ruiz, P.A.; Tudela, J.; Garcia-Canovas, F. Kinetic characterization of the substrate specificity and mechanism of mushroom tyrosinase. Eur. J. Biochem. 2000, 267, 1270–1279. [Google Scholar] [CrossRef] [Green Version]
- Toussaint, O.; Lerch, K.J.B. Catalytic oxidation of 2-aminophenols and ortho hydroxylation of aromatic amines by tyrosinase. Biochemistry 1987, 26, 8567–8571. [Google Scholar] [CrossRef]
- Garcia-Molina, F.; Munoz, J.L.; Varon, R.; Rodriguez-Lopez, J.N.; Garcia-Canovas, F.; Tudela, J. A review on spectrophotometric methods for measuring the monophenolase and diphenolase activities of tyrosinase. J. Agric. Food Chem. 2007, 55, 9739–9749. [Google Scholar] [CrossRef]
- Rodríguez-López, J.N.; Serna-Rodríguez, P.; Tudela, J.; Varón, R.; Garcia-Cánovas, F. A continuous spectrophotometric method for the determination of diphenolase activity of tyrosinase using 3,4-dihydroxymandelic acid. Anal. Biochem. 1991, 195, 369–374. [Google Scholar] [CrossRef]
- Jimenez, M.; Garcia-Carmona, F.; Garcia-Canovas, F.; Iborra, J.L.; Lozano, J.A.; Martinez, F. Chemical intermediates in dopamine oxidation by tyrosinase, and kinetic studies of the process. Arch. Biochem. Biophys. 1984, 235, 438–448. [Google Scholar] [CrossRef]
- Jiménez, M.; García-Cánovas, F.; García-Carmona, F.; Iborra, J.L.; Lozano, J.A. Isoproterenol oxidation by tyrosinase: Intermediates characterization and kinetic study. Biochem. Int. 1985, 11, 51–59. [Google Scholar]
- Rzepecki, L.M.; Waite, J.H. A chromogenic assay for catecholoxidases based on the addition of L-proline to quinones. Anal. Biochem. 1989, 179, 375–381. [Google Scholar] [CrossRef]
- Gauillard, F.; Richard-Forget, F.; Nicolas, J. New spectrophotometric assay for polyphenol oxidase activity. Anal. Biochem. 1993, 215, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.B.; Feng, L.; Jin, Q.; Wang, Y.R.; Liu, Z.M.; Zhu, X.Y.; Wang, P.; Hou, J.; Cui, J.N.; Yang, L. A novel substrate-inspired fluorescent probe to monitor native albumin in human plasma and living cells. Anal. Chim. Acta 2017, 989, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-R.; Feng, L.; Xu, L.; Hou, J.; Jin, Q.; Zhou, N.; Lin, Y.; Cui, J.-N.; Ge, G.-B. An ultrasensitive and conformation sensitive fluorescent probe for sensing human albumin in complex biological samples. Sens. Actuators B Chem. 2017, 245, 923–931. [Google Scholar] [CrossRef]
- Kamel, S.; Khattab, T.A.J.B. Recent Advances in Cellulose-Based Biosensors for Medical Diagnosis. Biosensors 2020, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.D.; Pang, H.L.; Zhou, Q.H.; Qin, Z.F.; Jin, Q.; Finel, M.; Wang, Y.N.; Qin, W.W.; Lu, Y.; Wang, D.D.; et al. An ultra-sensitive and easy-to-use assay for sensing human UGT1A1 activities in biological systems. J. Pharm. Anal. 2020, 10, 263–270. [Google Scholar] [CrossRef]
- Haghbeen, K.; Tan, E.W. Direct spectrophotometric assay of monooxygenase and oxidase activities of mushroom tyrosinase in the presence of synthetic and natural substrates. Anal. Biochem. 2003, 312, 23–32. [Google Scholar] [CrossRef]
- Munoz, J.L.; Garcia-Molina, F.; Varon, R.; Rodriguez-Lopez, J.N.; Garcia-Canovas, F.; Tudela, J. Calculating molar absorptivities for quinones: Application to the measurement of tyrosinase activity. Anal. Biochem. 2006, 351, 128–138. [Google Scholar] [CrossRef]
- Lin, Y.F.; Hu, Y.H.; Lin, H.T.; Liu, X.; Chen, Y.H.; Zhang, S.; Chen, Q.X. Inhibitory effects of propyl gallate on tyrosinase and its application in controlling pericarp browning of harvested longan fruits. J. Agric. Food Chem. 2013, 61, 2889–2895. [Google Scholar] [CrossRef] [PubMed]
- Winder, A.J.; Harris, H. New assays for the tyrosine hydroxylase and dopa oxidase activities of tyrosinase. Eur. J. Biochem. 1991, 198, 317–326. [Google Scholar] [CrossRef]
- Moonrungsee, N.; Shimamura, T.; Kashiwagi, T.; Jakmunee, J.; Higuchi, K.; Ukeda, H. Sequential injection spectrophotometric system for evaluation of mushroom tyrosinase-inhibitory activity. Talanta 2012, 101, 233–239. [Google Scholar] [CrossRef]
- Nolan, L.C.; O’Connor, K.E. A spectrophotometric method for the quantification of an enzyme activity producing 4-substituted phenols: Determination of toluene-4-monooxygenase activity. Anal. Biochem. 2005, 344, 224–231. [Google Scholar] [CrossRef]
- Mann, T.; Scherner, C.; Röhm, K.H.; Kolbe, L. Structure-Activity Relationships of Thiazolyl Resorcinols, Potent and Selective Inhibitors of Human Tyrosinase. Int. J. Mol. Sci. 2018, 19, 690. [Google Scholar] [CrossRef] [Green Version]
- Rescigno, A.; Sanjust, E.; Soddu, G.; Rinaldi, A.C.; Sollai, F.; Curreli, N.; Rinaldi, A. Effect of 3-hydroxyanthranilic acid on mushroom tyrosinase activity. Biochim. Biophys. Acta 1998, 1384, 268–276. [Google Scholar] [CrossRef]
- Teng, Y.; Jia, X.; Li, J.; Wang, E. Ratiometric fluorescence detection of tyrosinase activity and dopamine using thiolate-protected gold nanoclusters. Anal. Chem. 2015, 87, 4897–4902. [Google Scholar] [CrossRef]
- Zhang, W.H.; Ma, W.; Long, Y.T. Redox-Mediated Indirect Fluorescence Immunoassay for the Detection of Disease Biomarkers Using Dopamine-Functionalized Quantum Dots. Anal. Chem. 2016, 88, 5131–5136. [Google Scholar] [CrossRef]
- Liu, S.; Tian, J.; Wang, L.; Sun, X. Highly sensitive and selective colorimetric detection of Ag(I) ion using 3,3′,5,5′,-tetramethylbenzidine (TMB) as an indicator. Sens. Actuators B Chem. 2012, 165, 44–47. [Google Scholar] [CrossRef]
- Ni, P.; Sun, Y.; Dai, H.; Hu, J.; Jiang, S.; Wang, Y.; Li, Z. Highly sensitive and selective colorimetric detection of glutathione based on Ag [I] ion-3,3′,5,5′-tetramethylbenzidine (TMB). Biosens. Bioelectron. 2015, 63, 47–52. [Google Scholar] [CrossRef]
- Lei, C.; Zhao, X.E.; Sun, J.; Yan, X.; Gao, Y.; Gao, H.; Zhu, S.; Wang, H. A simple and novel colorimetric assay for tyrosinase and inhibitor screening using 3,3′,5,5′-tetramethylbenzidine as a chromogenic probe. Talanta 2017, 175, 457–462. [Google Scholar] [CrossRef]
- Deng, H.H.; Lin, X.L.; He, S.B.; Wu, G.W.; Wu, W.H.; Yang, Y.; Lin, Z.; Peng, H.P.; Xia, X.H.; Chen, W. Colorimetric tyrosinase assay based on catechol inhibition of the oxidase-mimicking activity of chitosan-stabilized platinum nanoparticles. Mikrochim Acta 2019, 186, 301. [Google Scholar] [CrossRef] [PubMed]
- Acuna, A.U.; Alvarez-Perez, M.; Liras, M.; Coto, P.B.; Amat-Guerri, F. Synthesis and photophysics of novel biocompatible fluorescent oxocines and azocines in aqueous solution. Phys. Chem. Chem. Phys. 2013, 15, 16704–16712. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Bao, X.; Wang, S.; Lu, S.; Sun, J.; Yang, X. In Situ Fluorogenic and Chromogenic Reactions for the Sensitive Dual-Readout Assay of Tyrosinase Activity. Anal. Chem. 2017, 89, 10529–10536. [Google Scholar] [CrossRef]
- Sun, W.; Li, M.; Fan, J.; Peng, X.J.A. Activity-Based Sensing and Theranostic Probes Based on Photoinduced Electron Transfer. ACC Chem Res. 2019, 52, 2818–2831. [Google Scholar] [CrossRef]
- Zou, L.; Wang, P.; Qian, X.; Feng, L.; Yu, Y.; Wang, D.; Jin, Q.; Hou, J.; Liu, Z.; Ge, G.; et al. Bioelectronics, a highly specific ratiometric two-photon fluorescent probe to detect dipeptidyl peptidase IV in plasma and living systems. Biosens. Bioelectron. 2017, 90, 283–289. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, H.; Zhong, M.; Wang, S.; Xu, Q.; Cho, D.-H.; Qiu, H. A novel off-on fluorescent probe for specific detection and imaging of cysteine in live cells and in vivo. Chin. Chem. Lett. 2020, 31, 153–155. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Zhang, Y.; Cao, Q.; Chen, Y.J.C.C.L. A GSH-responsive PET-based fluorescent probe for cancer cells imaging. Chin. Chem. Lett. 2020, 32, 1541–1544. [Google Scholar] [CrossRef]
- Yang, S.; Jiang, J.; Zhou, A.; Zhou, Y.; Ye, W.; Cao, D.S.; Yang, R. Substrate-Photocaged Enzymatic Fluorogenic Probe Enabling Sequential Activation for Light-Controllable Monitoring of Intracellular Tyrosinase Activity. Anal. Chem. 2020, 92, 7194–7199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, H.; Yao, Q.; Ghazali, S.; Fan, J.; Wang, J.; Gao, F.; Li, M.; Wang, H.; Dong, C.; et al. A unique two-photon fluorescent probe based on ICT mechanism for imaging palladium in living cells and mice. Chin. Chem. Lett. 2020, 31, 2913–2916. [Google Scholar] [CrossRef]
- Jin, Q.; Feng, L.; Zhang, S.J.; Wang, D.D.; Wang, F.; Zhang, Y.; Cui, J.N.; Guo, W.Z.; Ge, G.B.; Yang, L.J.A.C. Real-Time Tracking the Synthesis and Degradation of Albumin in Complex Biological Systems with a near-Infrared Fluorescent Probe. Anal. Chem. 2017, 89, 9884–9891. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Tiwari, K.; Tiwari, R.; Pramanik, S.K.; Das, A. Small Molecule as Fluorescent Probes for Monitoring Intracellular Enzymatic Transformations. Chem. Rev. 2019, 119, 11718–11760. [Google Scholar] [CrossRef]
- Zhao, C.; An, J.; Zhou, L.; Fei, Q.; Wang, F.; Tan, J.; Shi, B.; Wang, R.; Guo, Z.; Zhu, W.H. Transforming the recognition site of 4-hydroxyaniline into 4-methoxyaniline grafted onto a BODIPY core switches the selective detection of peroxynitrite to hypochlorous acid. Chem. Commun. 2016, 52, 2075–2078. [Google Scholar] [CrossRef]
- Lu, P.; Zhang, X.; Ren, T.; Yuan, L.J.C.C.L. Molecular engineering of ultra-sensitive fluorescent probe with large Stokes shift for imaging of basal HOCl in tumor cells and tissues. Chin. Chem. Lett. 2020, 31, 2980–2984. [Google Scholar] [CrossRef]
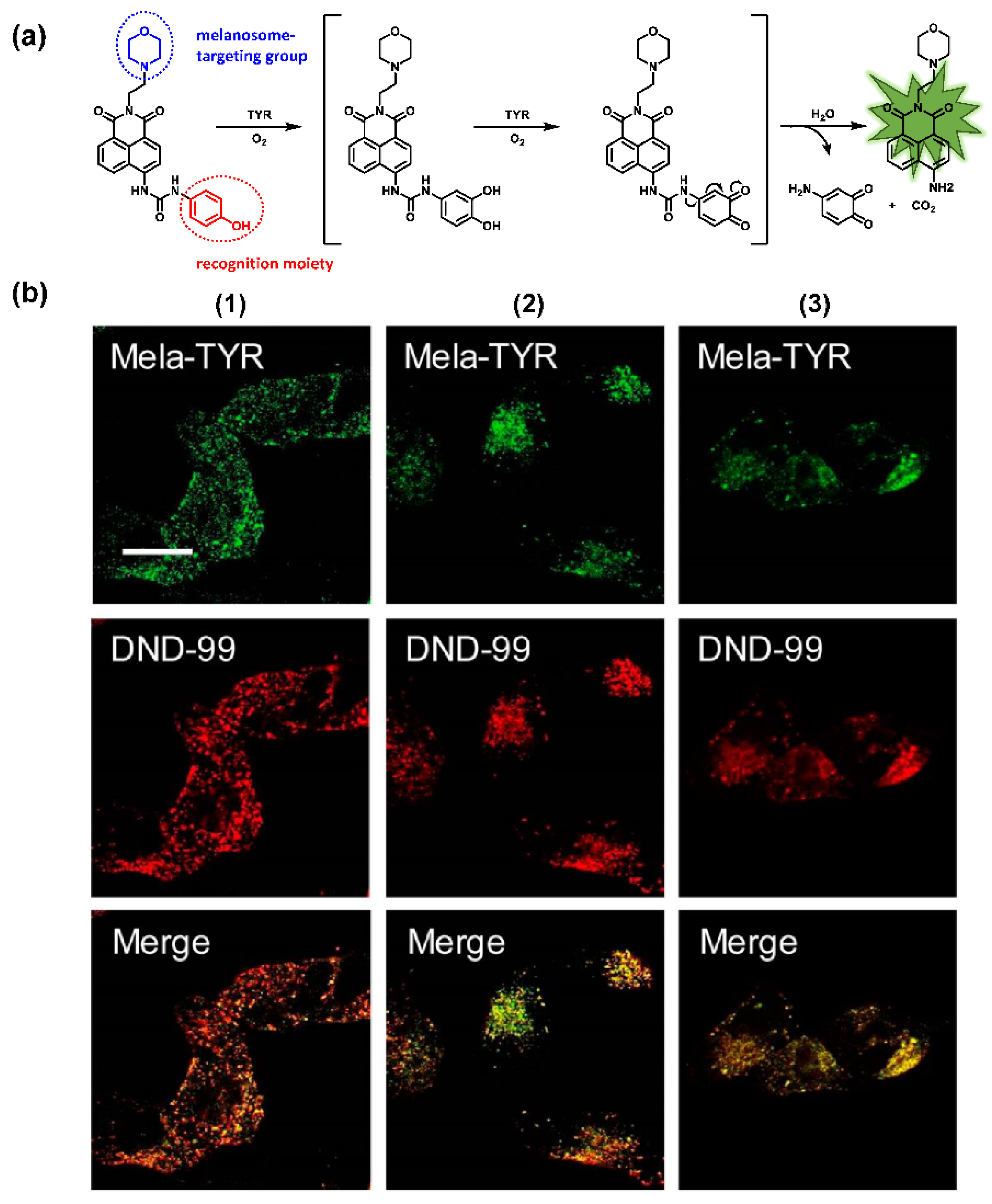
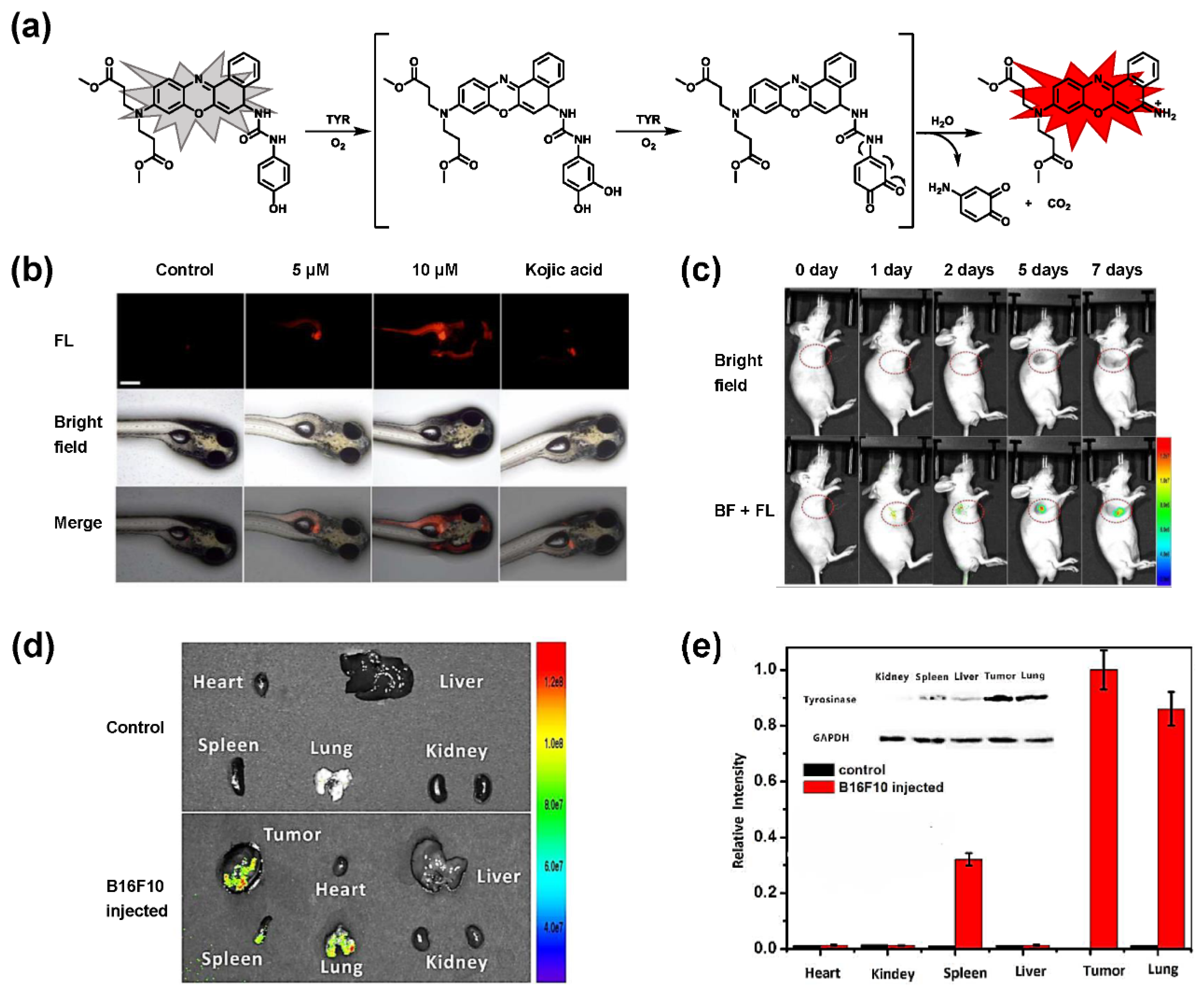
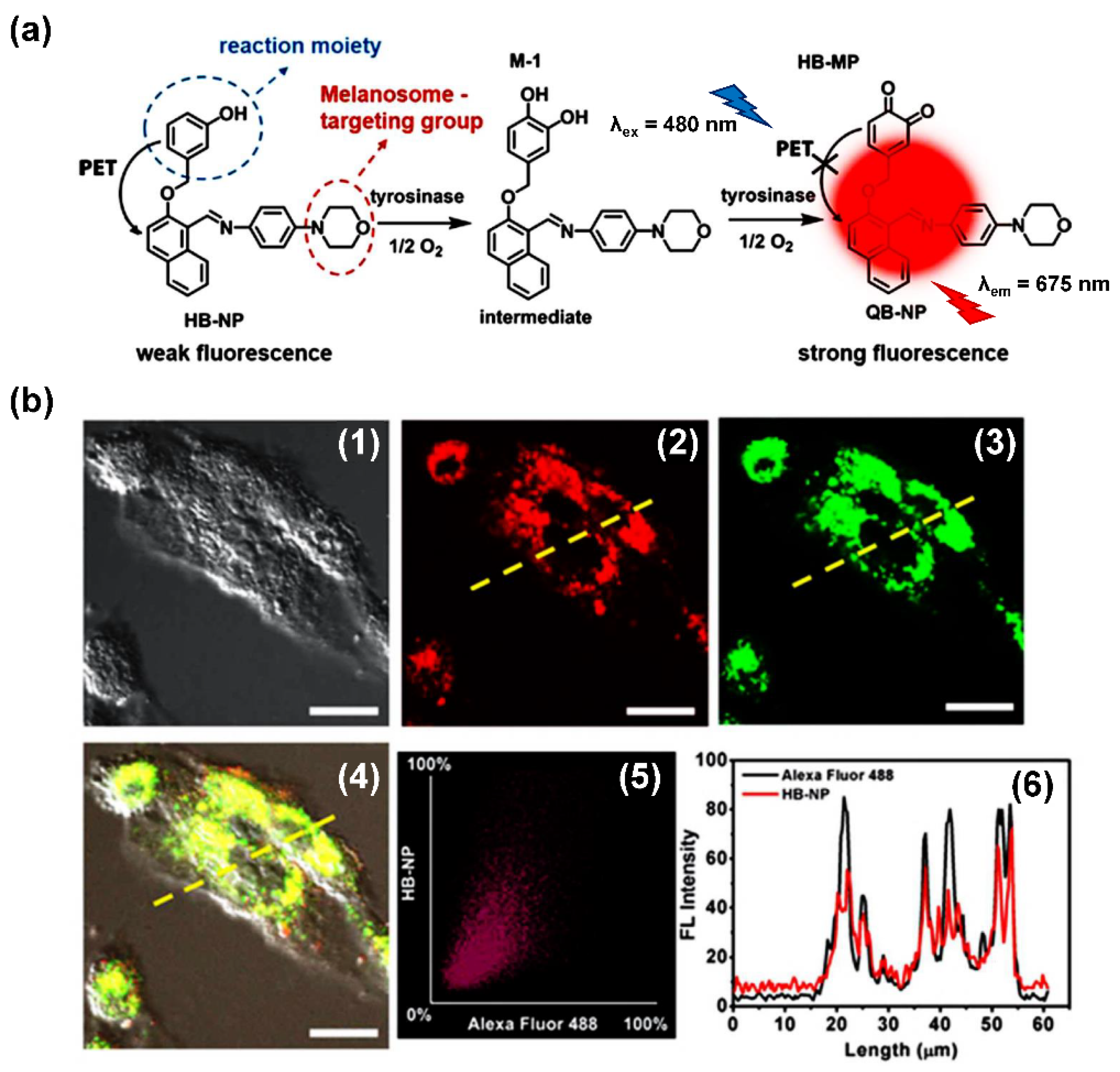
- Peng, M.; Wang, Y.; Fu, Q.; Sun, F.; Na, N.; Ouyang, J. Melanosome-Targeting Near-Infrared Fluorescent Probe with Large Stokes Shift for in Situ Quantification of Tyrosinase Activity and Assessing Drug Effects on Differently Invasive Melanoma Cells. Anal. Chem. 2018, 90, 6206–6213. [Google Scholar] [CrossRef]
- Sidhu, J.S.; Singh, A.; Garg, N.; Kaur, N.; Singh, N. A highly selective naphthalimide-based ratiometric fluorescent probe for the recognition of tyrosinase and cellular imaging. Analyst 2018, 143, 4476–4483. [Google Scholar] [CrossRef]
- Wu, X.; Li, L.; Shi, W.; Gong, Q.; Ma, H. Near-Infrared Fluorescent Probe with New Recognition Moiety for Specific Detection of Tyrosinase Activity: Design, Synthesis, and Application in Living Cells and Zebrafish. Angew. Chem. Int. Ed. Engl. 2016, 55, 14728–14732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, Z.; Tian, X.; Ding, N. A novel hydrosoluble near-infrared fluorescent probe for specifically monitoring tyrosinase and application in a mouse model. Chem. Commun. 2019, 55, 9463–9466. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, X.; Li, H.; Shi, W.; Ma, H. A highly sensitive and selective fluorescence off-on probe for the detection of intracellular endogenous tyrosinase activity. Chem. Commun. 2017, 53, 2443–2446. [Google Scholar] [CrossRef]
- Li, H.; Liu, W.; Zhang, F.; Zhu, X.; Huang, L.; Zhang, H. Highly Selective Fluorescent Probe Based on Hydroxylation of Phenylboronic Acid Pinacol Ester for Detection of Tyrosinase in Cells. Anal. Chem. 2018, 90, 855–858. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Huang, R.; Wang, C.; Zhou, Y.; Wang, J.; Fu, B.; Weng, X.; Zhou, X. A two-photon fluorescent probe for intracellular detection of tyrosinase activity. Chem. Asian. J. 2012, 7, 2782–2785. [Google Scholar] [CrossRef]
- Wang, C.; Yan, S.; Huang, R.; Feng, S.; Fu, B.; Weng, X.; Zhou, X. A turn-on fluorescent probe for detection of tyrosinase activity. Analyst 2013, 138, 2825–2828. [Google Scholar] [CrossRef]
- Zhan, C.; Cheng, J.; Li, B.; Huang, S.; Zeng, F.; Wu, S. A Fluorescent Probe for Early Detection of Melanoma and Its Metastasis by Specifically Imaging Tyrosinase Activity in a Mouse Model. Anal. Chem. 2018, 90, 8807–8815. [Google Scholar] [CrossRef]
- Zhou, J.; Shi, W.; Li, L.; Gong, Q.; Wu, X.; Li, X.; Ma, H. Detection of Misdistribution of Tyrosinase from Melanosomes to Lysosomes and Its Upregulation under Psoralen/Ultraviolet A with a Melanosome-Targeting Tyrosinase Fluorescent Probe. Anal. Chem. 2016, 88, 4557–4564. [Google Scholar] [CrossRef]
- Zhang, P.; Li, S.; Fu, C.; Zhang, Q.; Xiao, Y.; Ding, C. A colorimetric and near -infrared ratiometric fluorescent probe for the determination of endogenous tyrosinase activity based on cyanine aggregation. Analyst 2019, 144, 5472–5478. [Google Scholar] [CrossRef]
- Li, X.; Shi, W.; Chen, S.; Jia, J.; Ma, H.; Wolfbeis, O.S. A near-infrared fluorescent probe for monitoring tyrosinase activity. Chem. Commun. 2010, 46, 2560–2562. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.F.; Zeng, C.; Hu, L.; Liang, X.J. Ultrasensitive Tyrosinase-Activated Turn-On Near-Infrared Fluorescent Probe with a Rationally Designed Urea Bond for Selective Imaging and Photodamage to Melanoma Cells. Anal. Chem. 2018, 90, 3666–3669. [Google Scholar] [CrossRef] [Green Version]
- Naidu Bobba, K.; Won, M.; Shim, I.; Velusamy, N.; Yang, Z.; Qu, J.; Kim, J.S.; Bhuniya, S. A BODIPY-based two-photon fluorescent probe validates tyrosinase activity in live cells. Chem. Commun. 2017, 53, 11213–11216. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.I.; Park, J.; Park, S.; Choi, Y.; Kim, Y. Visualization of tyrosinase activity in melanoma cells by a BODIPY-based fluorescent probe. Chem. Commun. 2011, 47, 12640–12642. [Google Scholar] [CrossRef]
- Li, Z.; Xia, X.; You, Y.; Lu, C.; Yang, G.; Ma, C.; Nie, J.; Sun, Q.; Wu, S.; Ren, J.; et al. Fast-response fluorescent probe with favorable water solubility for highly sensitive imaging of endogenous tyrosinase in living cells and zebrafish model. Chin. Chem. Lett. 2021, 32, 1785–1789. [Google Scholar] [CrossRef]
- Zhou, Q.H.; Qin, W.W.; Finel, M.; He, Q.Q.; Ge, G.B. A broad-spectrum substrate for the human UDP-glucuronosyltransferases and its use for investigating glucuronidation inhibitors. Int. J. Biol. Macromol. 2021, 180, 252–261. [Google Scholar] [CrossRef]
- Qiang, J.A.; Hm, A.; Lei, F.B.; Ping, W.A.; Rh, A.; Jing, N.B.; Ling, Y.A.; Gg, A.J.C.C.L. Sensing cytochrome P450 1A1 activity by a resorufin-based isoform-specific fluorescent probe. Chin. Chem. Lett. 2020, 31, 2945–2949. [Google Scholar]
- Dl, A.; Wc, A.; Sheng, H.; Xc, B.; Jya, B.J.C.C.L. The regulation of biothiol-responsive performance and bioimaging application of benzo[c][1,2,5]oxadiazole dyes. Chin. Chem. Lett. 2020, 31, 2891–2896. [Google Scholar]
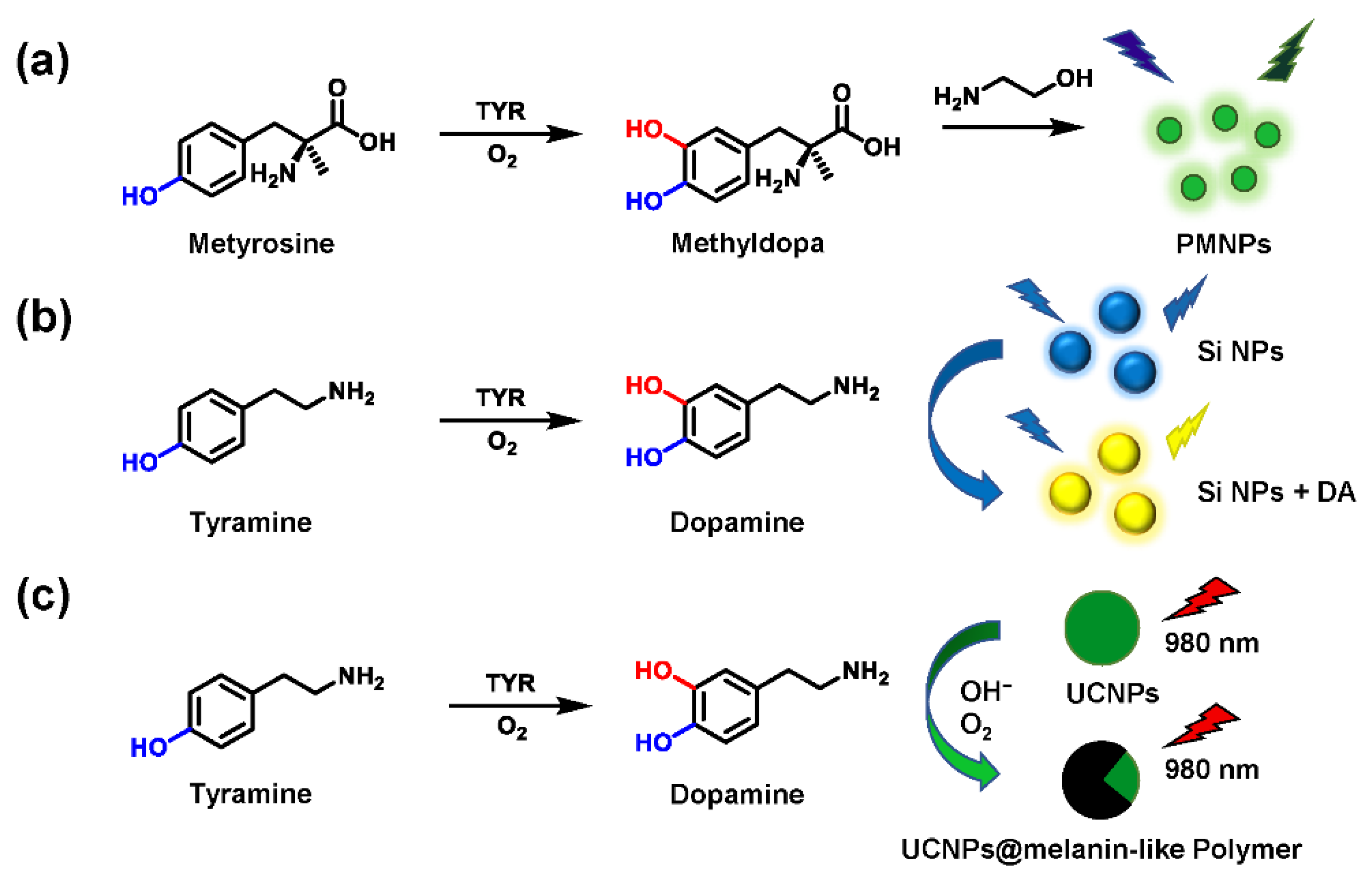
- Liu, G.; Zhao, J.; Lu, S.; Wang, S.; Sun, J.; Yang, X. Polymethyldopa Nanoparticles-Based Fluorescent Sensor for Detection of Tyrosinase Activity. ACS Sens. 2018, 3, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.Z.; Wang, W.F.; Chai, T.; Qiang, Y.; Shi, Y.P.; Yang, J.L. Ratiometric target-triggered fluorescent silicon nanoparticles probe for quantitative visualization of tyrosinase activity. Talanta 2019, 197, 113–121. [Google Scholar] [CrossRef]
- Wang, H.; Lu, Y.; Wang, L.; Chen, H. Detection of tyramine and tyrosinase activity using red region emission NaGdF4:Yb, Er@NaYF4 upconversion nanoparticles. Talanta 2019, 197, 558–566. [Google Scholar] [CrossRef]
- Pinho, J.O.; Matias, M.; Gaspar, M.M. Emergent Nanotechnological Strategies for Systemic Chemotherapy against Melanoma. Nanomaterials 2019, 9, 1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kounalakis, N.; Goydos, J.S. Tumor cell and circulating markers in melanoma: Diagnosis, prognosis, and management. Curr. Oncol. Rep. 2005, 7, 377–382. [Google Scholar] [CrossRef]
- Reintgen, D.; Cruse, C.; Wells, K.; Berman, C.; Fenske, N.; Glass, F.; Schroer, K.; Heller, R.; Ross, M.; Lyman, G.; et al. The orderly progression of melanoma nodal metastases. Ann. Surg. 1994, 220, 759–767. [Google Scholar] [CrossRef]
- Parkin, D.M.; Pisani, P.; Ferlay, J. Estimates of the worldwide incidence of 25 major cancers in 1990. Int. J. Cancer 1999, 80, 827–841. [Google Scholar] [CrossRef]
- Heo, J.R.; Kim, N.H.; Cho, J.; Choi, K.C. Current treatments for advanced melanoma and introduction of a promising novel gene therapy for melanoma (Review). Oncol. Rep. 2016, 36, 1779–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, R.A.; Pietsch, J.; Fryer, J.P.; Savage, S.; Brott, M.J.; Russell-Eggitt, I.; Summers, C.G.; Oetting, W.S.J.H.G. Tyrosinase gene mutations in oculocutaneous albinism1 (OCA1): Definition of the phenotype. Hum Genet. 2003, 113, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.; Selby, P.; Southgate, J.; Pittman, K.; Bradley, C.; Blair, G.E. Detection of melanoma cells in peripheral blood by means of reverse transcriptase and polymerase chain reaction. Lancet 1991, 338, 1227–1229. [Google Scholar] [CrossRef]
- Palmieri, G.; Ascierto, P.A.; Perrone, F.; Satriano, S.M.; Ottaiano, A.; Daponte, A.; Napolitano, M.; Caraco, C.; Mozzillo, N.; Melucci, M.T.; et al. Prognostic value of circulating melanoma cells detected by reverse transcriptase-polymerase chain reaction. J. Clin. Oncol. 2003, 21, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Quaglino, P.; Savoia, P.; Osella-Abate, S.; Bernengo, M.G. RT-PCR tyrosinase expression in the peripheral blood of melanoma patients. Expert Rev. Mol. Diagn. 2004, 4, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Visús, C.; Andres, R.; Mayordomo, J.I.; Martinez-Lorenzo, M.J.; Murillo, L.; Sáez-Gutiérrez, B.; Diestre, C.; Marcos, I.; Astier, P.; Godino, J.J.M.R. Prognostic role of circulating melanoma cells detected by reverse transcriptase-polymerase chain reaction for tyrosinase mRNA in patients with melanoma. Melanoma Res. 2007, 17, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Mendes, E.; Perry, M.D.J.; Francisco, A.P. Design and discovery of mushroom tyrosinase inhibitors and their therapeutic applications. Expert Opin. Drug Dis. 2014, 9, 533–554. [Google Scholar] [CrossRef]
- Ullah, S.; Son, S.; Yun, H.Y.; Kim, D.H.; Chun, P.; Moon, H.R. Tyrosinase inhibitors: A patent review (2011–2015). Expert Opin. Ther. Pat. 2016, 26, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Leyden, J.J.; Shergill, B.; Micali, G.; Downie, J.; Wallo, W. Natural options for the management of hyperpigmentation. J. Eur. Acad Derm. Venereol 2011, 25, 1140–1145. [Google Scholar] [CrossRef]
- Zhu, T.H.; Cao, S.W.; Yu, Y.Y. Synthesis, characterization and biological evaluation of paeonol thiosemicarbazone analogues as mushroom tyrosinase inhibitors. Int. J. Biol. Macromol. 2013, 62, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Kinst-Hori, I.; Chaudhuri, S.K.; Kubo, Y.; Sanchez, Y.; Ogura, T. Flavonols from Heterotheca inuloides: Tyrosinase inhibitory activity and structural criteria. Bioorg. Med. Chem 2000, 8, 1749–1755. [Google Scholar] [CrossRef]
- Jow, T.; Hantash, B.M. Hydroquinone-induced depigmentation: Case report and review of the literature. Dermatitis 2014, 25, e1–e5. [Google Scholar] [CrossRef]
- Draelos, Z.D. Skin lightening preparations and the hydroquinone controversy. Dermatol. Ther. 2007, 20, 308–313. [Google Scholar] [CrossRef]
- Searle, T.; Al-Niaimi, F.; Ali, F.R. The top 10 cosmeceuticals for facial hyperpigmentation. Dermatol. Ther. 2020, 33, e14095. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, J.; Li, A.; Reetz, M.T. Chemical and Biocatalytic Routes to Arbutin (dagger). Molecules 2019, 24, 3303. [Google Scholar] [CrossRef] [Green Version]
- Burnett, C.L.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G., Jr.; Shank, R.C.; Slaga, T.J.; Snyder, P.W.; et al. Final report of the safety assessment of Kojic acid as used in cosmetics. Int. J. Toxicol. 2010, 29, 244S–273S. [Google Scholar] [CrossRef]
- Saeedi, M.; Eslamifar, M.; Khezri, K. Kojic acid applications in cosmetic and pharmaceutical preparations. Biomed. Pharm. 2019, 110, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Ashooriha, M.; Khoshneviszadeh, M.; Khoshneviszadeh, M.; Rafiei, A.; Kardan, M.; Yazdian-Robati, R.; Emami, S. Kojic acid-natural product conjugates as mushroom tyrosinase inhibitors. Eur. J. Med. Chem. 2020, 201, 112480. [Google Scholar] [CrossRef]
- Tajima, R.; Oozeki, H.; Muraoka, S.; Tanaka, S.; Motegi, Y.; Nihei, H.; Yamada, Y.; Masuoka, N.; Nihei, K. Synthesis and evaluation of bibenzyl glycosides as potent tyrosinase inhibitors. Eur. J. Med. Chem. 2011, 46, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Ishioka, W.; Oonuki, S.; Iwadate, T.; Nihei, K.I. Resorcinol alkyl glucosides as potent tyrosinase inhibitors. Bioorg. Med. Chem. Lett. 2019, 29, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.J.; Noh, S.G.; Park, Y.; Kang, D.; Chun, P.; Chung, H.Y.; Moon, H.R. In vitro and in silico insights into tyrosinase inhibitors with (E)-benzylidene-1-indanone derivatives. Comput. Struct Biotechnol. J. 2019, 17, 1255–1264. [Google Scholar] [CrossRef]
- Durai, P.; Ko, Y.J.; Kim, J.C.; Pan, C.H.; Park, K. Identification of Tyrosinase Inhibitors and Their Structure-Activity Relationships via Evolutionary Chemical Binding Similarity and Structure-Based Methods. Molecules 2021, 26, 566. [Google Scholar] [CrossRef]
- Wang, D.; Zou, L.; Jin, Q.; Hou, J.; Ge, G.; Yang, L. Human carboxylesterases: A comprehensive review. Acta Pharm. Sinica. B 2018, 8, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Kondoh, H.; Ichihashi, M.; Hearing, V.J. Approaches to identify inhibitors of melanin biosynthesis via the quality control of tyrosinase. J. Investig. Derm. 2007, 127, 751–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Kim, Y.J.; Kang, J.S.; Kim, I.Y.; Choi, K.S.; Kim, H.M. Carboxylesterase-2-Selective Two-Photon Ratiometric Probe Reveals Decreased Carboxylesterase-2 Activity in Breast Cancer Cells. Anal. Chem. 2018, 90, 9465–9471. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Shi, W.; Li, X.; Ma, H. Recognition Moieties of Small Molecular Fluorescent Probes for Bioimaging of Enzymes. Acc. Chem. Res. 2019, 52, 1892–1904. [Google Scholar] [CrossRef]
- Li, S.; Hu, R.; Wang, S.; Guo, X.; Zeng, Y.; Li, Y.; Yang, G. Specific Imaging of Tyrosinase in Vivo with 3-Hydroxybenzyl Caged D-Luciferins. Anal. Chem. 2018, 90, 9296–9300. [Google Scholar] [CrossRef]
- Yi, X.; Wang, F.; Qin, W.; Yang, X.; Yuan, J. Near-infrared fluorescent probes in cancer imaging and therapy: An emerging field. Int. J. Nanomed. 2014, 9, 1347–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vendrell, M.; Zhai, D.; Er, J.C.; Chang, Y.T. Combinatorial strategies in fluorescent probe development. Chem. Rev. 2012, 112, 4391–4420. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Ogawa, M.; Alford, R.; Choyke, P.L.; Urano, Y. New strategies for fluorescent probe design in medical diagnostic imaging. Chem. Rev. 2010, 110, 2620–2640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pode, Z.; Peri-Naor, R.; Georgeson, J.M.; Ilani, T.; Kiss, V.; Unger, T.; Markus, B.; Barr, H.M.; Motiei, L.; Margulies, D. Protein recognition by a pattern-generating fluorescent molecular probe. Nat. Nanotechnol. 2017, 12, 1161–1168. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrates | Chemical Structures | Metabolite Stability | Coupled Reagent | Detectable Species | λ (nm) | ε (M−1·cm−1) | Ref. |
|---|---|---|---|---|---|---|---|
| 4-Tert-butylcatechol (TBC) |  | Stable | - | o-Quinone | 400 | 1200 | [44] |
| L-Tyrosine |  | Unstable | - | Dopachrome | 475 | 3600 | [45] |
| L-DOPA |  | Unstable | - | Dopachrome | 475 | 3600 | [45] |
| MBTH | MBTH-adduct | 484 | 22,300 | [42] | |||
| Dopamine (DA) |  | Unstable | MBTH | MBTH-adduct | 503 | 42,500 | [42] |
| Isoproterenol (ISO) |  | Unstable | MBTH | MBTH-adduct | 497 | 31,500 | [42] |
| Substrates | Chemical Structures | Enzyme Sources | Metabolites | Enzyme Activity | Km (μM) | Vmax (nmol/min/mg) | Ref. |
|---|---|---|---|---|---|---|---|
| L-Tyrosine |  | Mushroom | L-DOPA | Monophenolase activity | 270 | - | [41] |
| L-DOPA |  | Mushroom | Dopachrome | Diphenolase activity | 800 | - | [41] |
| 3-Hydroxyanthranilic acid |  | Mushroom | Cinnabarinic acid | Diphenolase activity | 780 | 12 | [61] |
| 4-Tertbutyl-catechol |  | Mushroom | 4-Tert-butyl-1,2-benzoquinone | Diphenolase activity | 990 | - | [37] |
| Names | Chemical Structures | λex/em (nm) | Folds | Km (μM) | Vmax (μM·min−1) | LOD (U·mL−1) | Biological Applications | Ref. |
|---|---|---|---|---|---|---|---|---|
| L3 |  | 425/467 425/535 | - | 43.5 | 1.87 | 0.2 | A375 cells | [81] |
| Probe 1 |  | 670/708 | - | 156 | 4.58 | 2.76 | B16 and HeLa cells, zebrafish | [82] |
| HB-NP |  | 480/675 | 48 | 87.35 | 1.07 | 0.5 | B16, HepG2, A549, HeLa, CCC-HPF-1 and CCC-HSF-1 cells; inhibitor screening. | [80] |
| Probe 1 |  | 670/708 | - | - | - | 0.11 | Imaging of TYR in B16 cells and zebrafish, melanoma diagnosis in a mouse. | [83] |
| Probe 1 |  | 550/586 | - | 30 | 0.72 | 0.04 | B16, HepG2 and MCF-7 cells | [84] |
| Probe 1 |  | 550/583 | - | 6.5 | 0.0009 | 0.5 | B16 and HepG2 cells | [85] |
| NHU |  | 382/503 | 12 | - | - | - | B16-F1 and HeLa cells | [86] |
| Probe 1 |  | 460/540 | 12 | - | - | - | Screening inhibitors | [87] |
| NBR-AP |  | 580/660 | 7 | 229.5 | 5.75 | - | Imaging of TYR in B16F10 and HeLa cells, in vivo imaging of zebrafish and mice | [88] |
| Mela-TYR |  | 425/547 | 50 | - | - | 0.07 | B16 and HeLa cells, the subcellular localization | [89] |
| Cy-tyr |  | 630/760 516/556 | - | - | - | 0.02 | B16, HeLa, MCF-7 and HUVEC cells | [90] |
| Probe 1 |  | 650/720 | - | - | - | 0.01 | Screening inhibitors | [91] |
| MB1 |  | 640/706 | >100 | 4.6 | 0.45 | - | B16F10 and HeLa cells, photodynamic therapy | [92] |
| Tyro-1 |  | 400/452 | 12.5 | - | - | 0.025 | B16F10 and HeLa cells | [93] |
| Probe 1 |  | 460/515 | - | 5.3 | 6.36 | - | B16F10 cells | [94] |
| CHMC-DOPA |  | 538/629 | - | - | - | 0.003 | HepG2 cells, zebrafish | [95] |
| Inhibitors | Chemical Structures | Sources | Adverse Effects | Dosage | Group | Ref. |
|---|---|---|---|---|---|---|
| Hydroquinone (HQ) |  | Plant |
| <4% | Forbidden | [117,118] |
| β-Arbutin |  | Plant | Facial tingling | <3% | Approved | [119,120] |
| Kojic acid (KA) |  | Fungus |
| <1% | Approved | [121,122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.-F.; Zhu, S.-X.; Hou, F.-B.; Zhao, D.-F.; Pan, Q.-S.; Xiang, Y.-W.; Qian, X.-K.; Ge, G.-B.; Wang, P. Spectrophotometric Assays for Sensing Tyrosinase Activity and Their Applications. Biosensors 2021, 11, 290. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11080290
Fan Y-F, Zhu S-X, Hou F-B, Zhao D-F, Pan Q-S, Xiang Y-W, Qian X-K, Ge G-B, Wang P. Spectrophotometric Assays for Sensing Tyrosinase Activity and Their Applications. Biosensors. 2021; 11(8):290. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11080290
Chicago/Turabian StyleFan, Yu-Fan, Si-Xing Zhu, Fan-Bin Hou, Dong-Fang Zhao, Qiu-Sha Pan, Yan-Wei Xiang, Xing-Kai Qian, Guang-Bo Ge, and Ping Wang. 2021. "Spectrophotometric Assays for Sensing Tyrosinase Activity and Their Applications" Biosensors 11, no. 8: 290. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11080290