Aniba rosaeodora (Var. amazonica Ducke) Essential Oil: Chemical Composition, Antibacterial, Antioxidant and Antitrypanosomal Activity

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
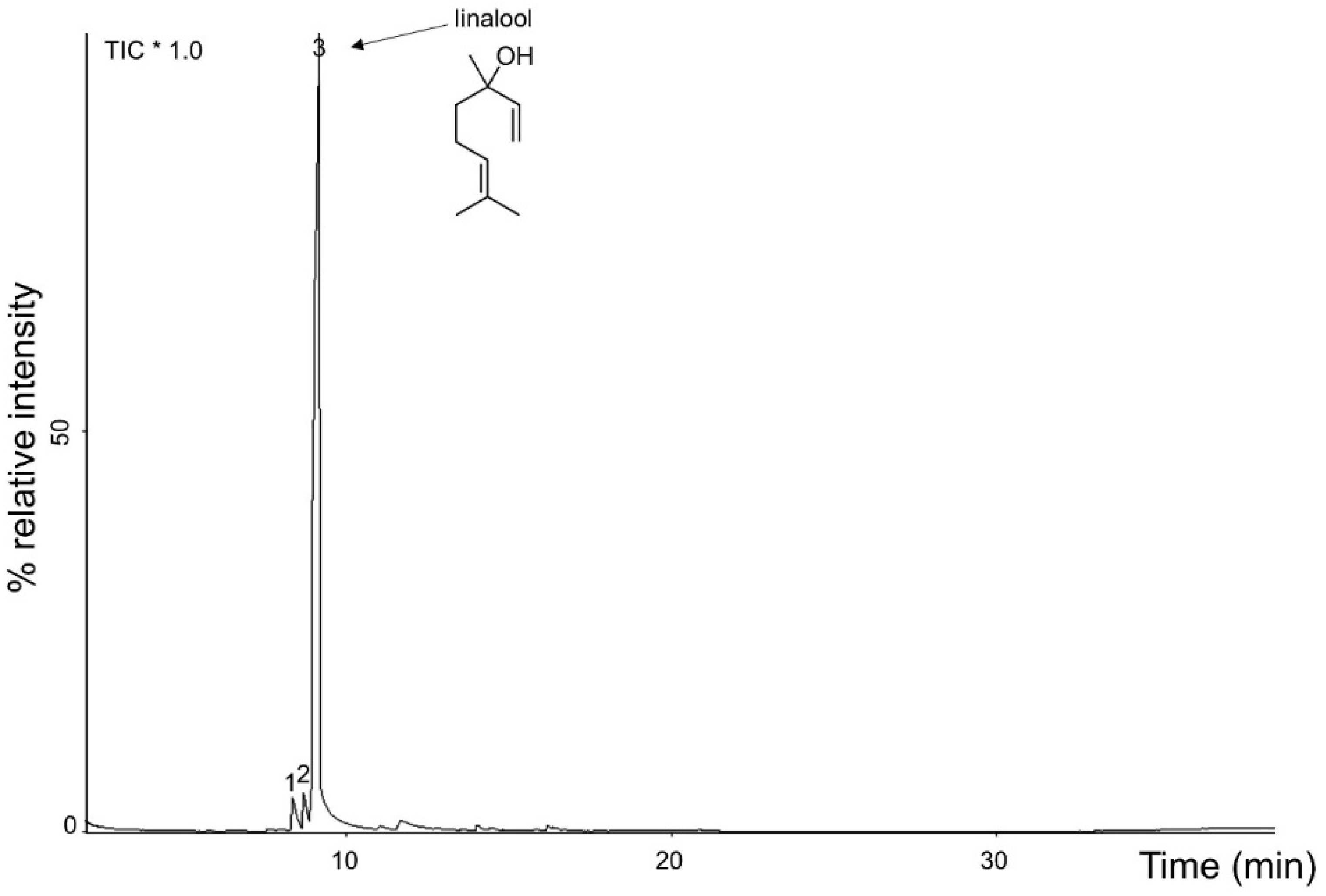
2.1. Physical and Chemical Characterization of A. rosaeodora Essential Oil
2.2. Antimicrobial Activity of A. rosaeodora Essential Oil and Linalool
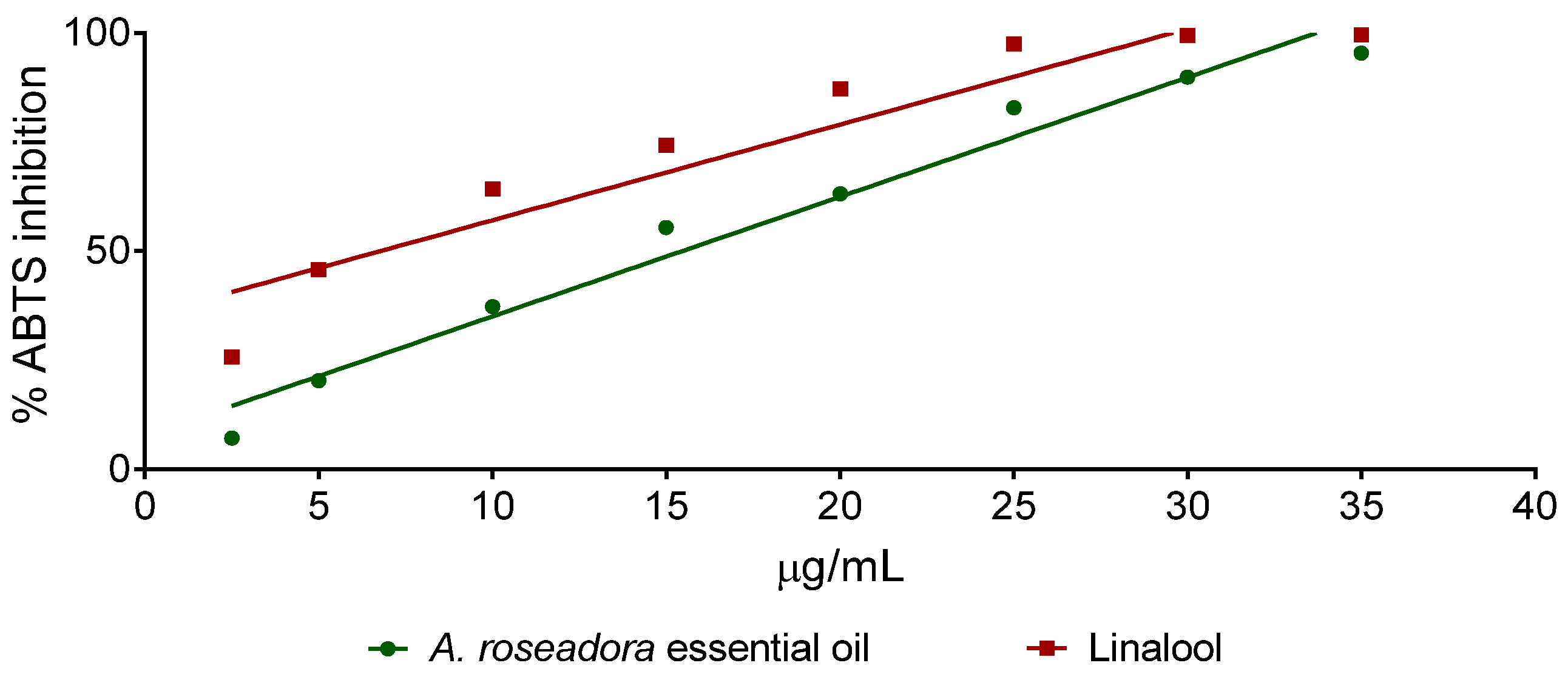
2.3. Antioxidant Activity of A. rosaeodora Essential Oil and Linalol
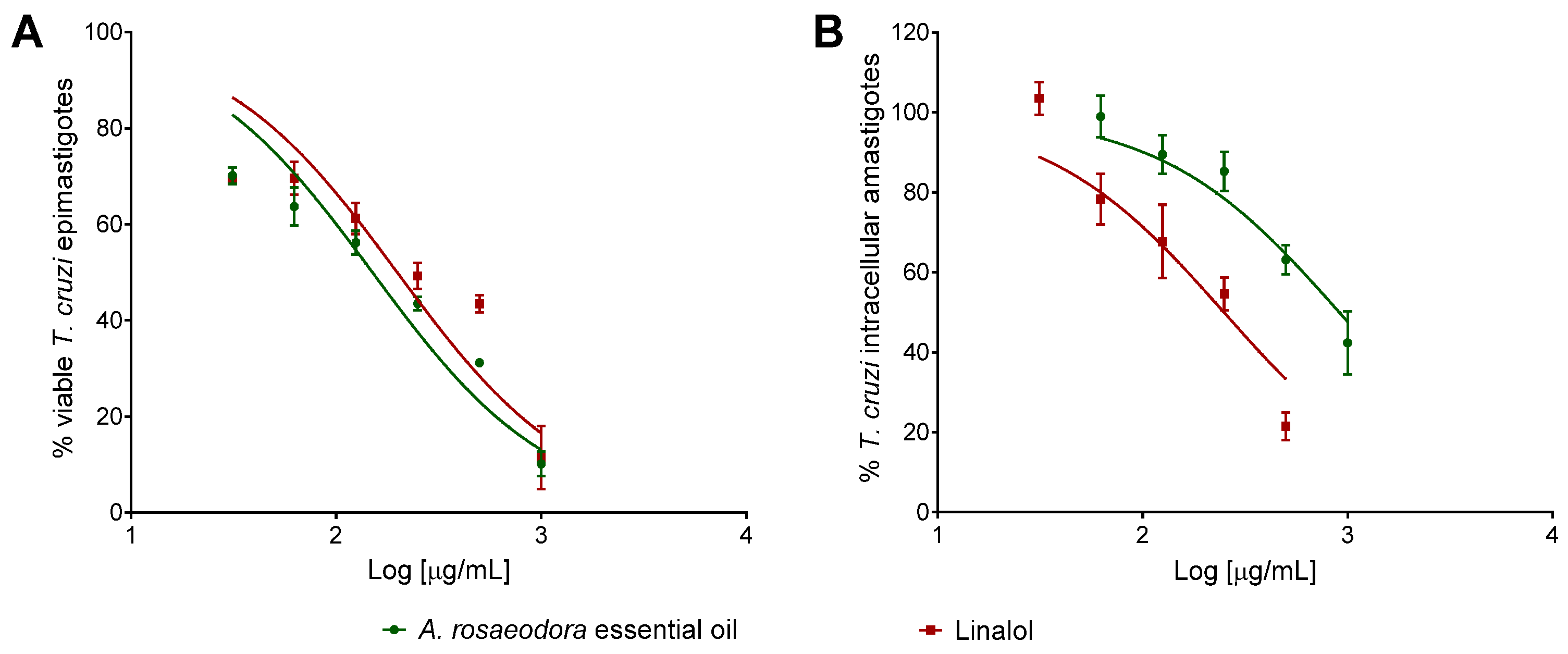
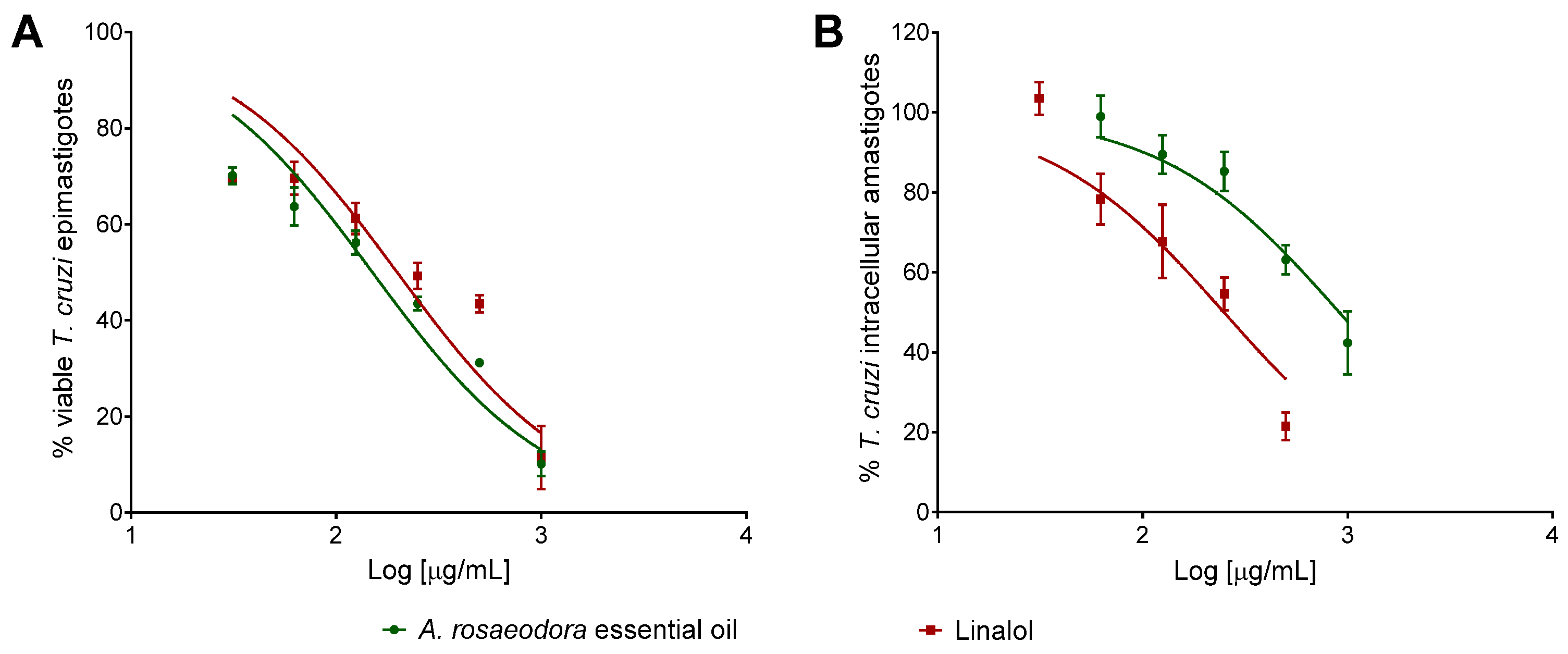
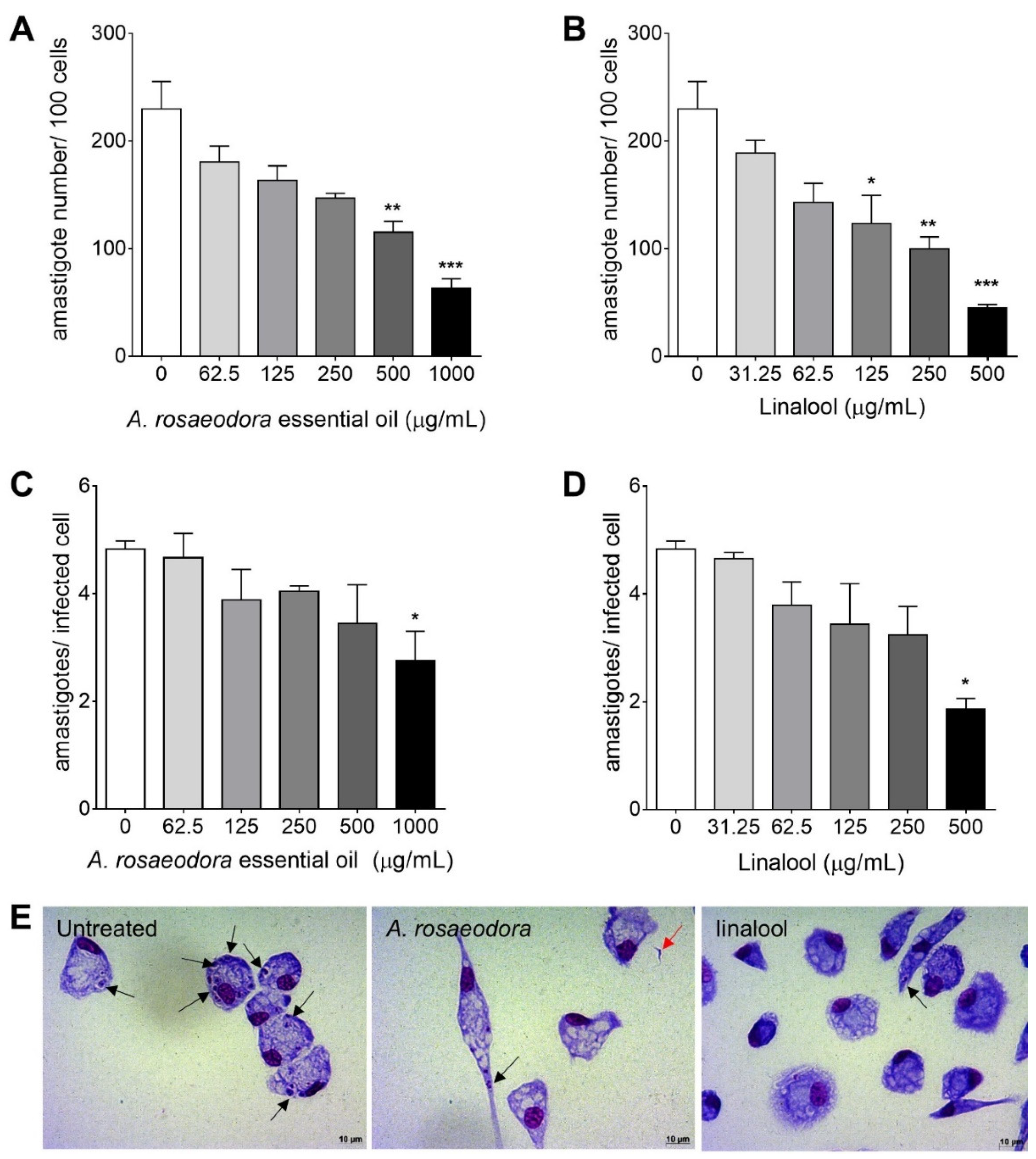
2.4. Cytotoxicity, Antitrypanosomal Activivy and Selectivity Index of A. rosaeodora Essential Oil and Linalool
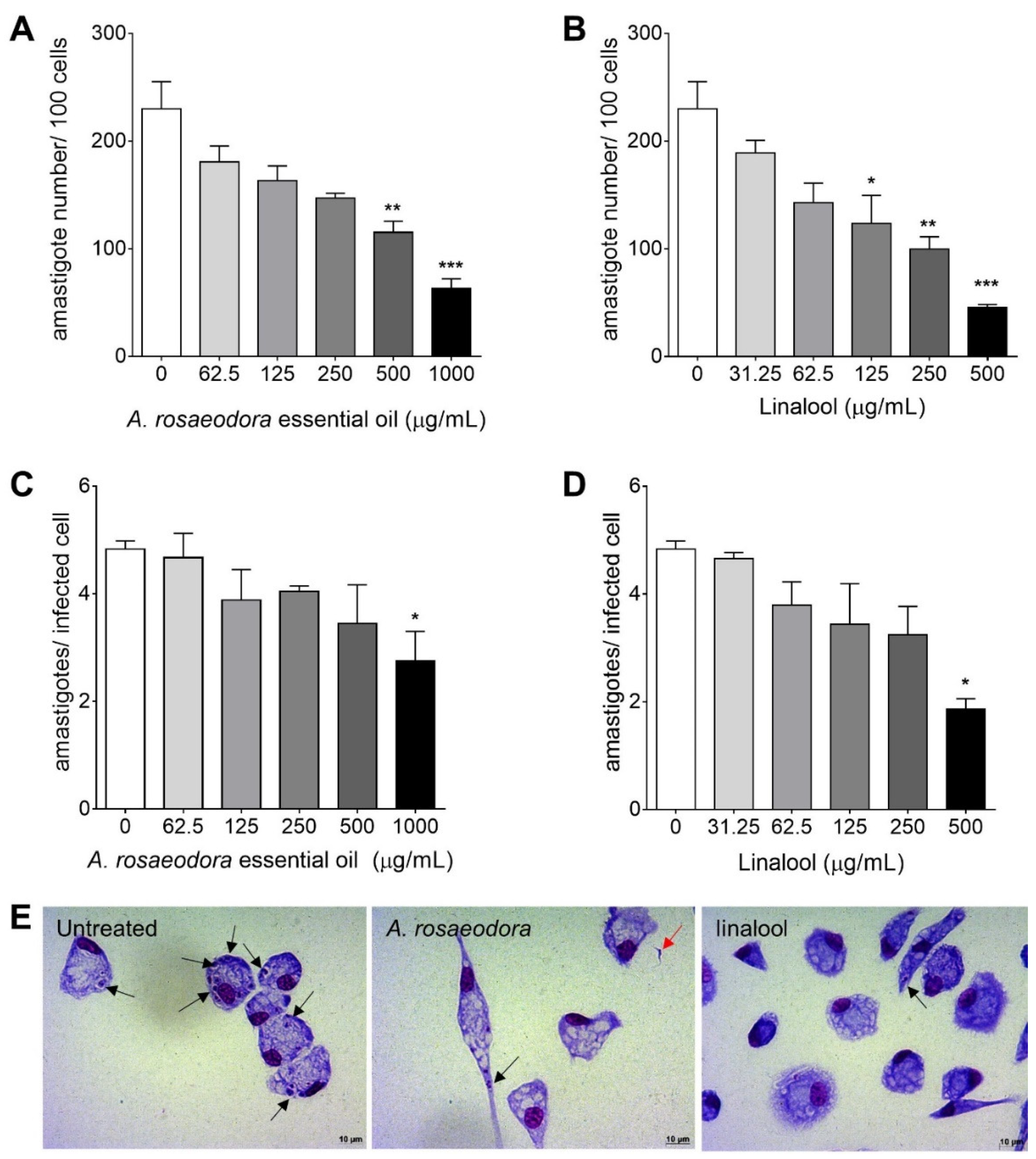
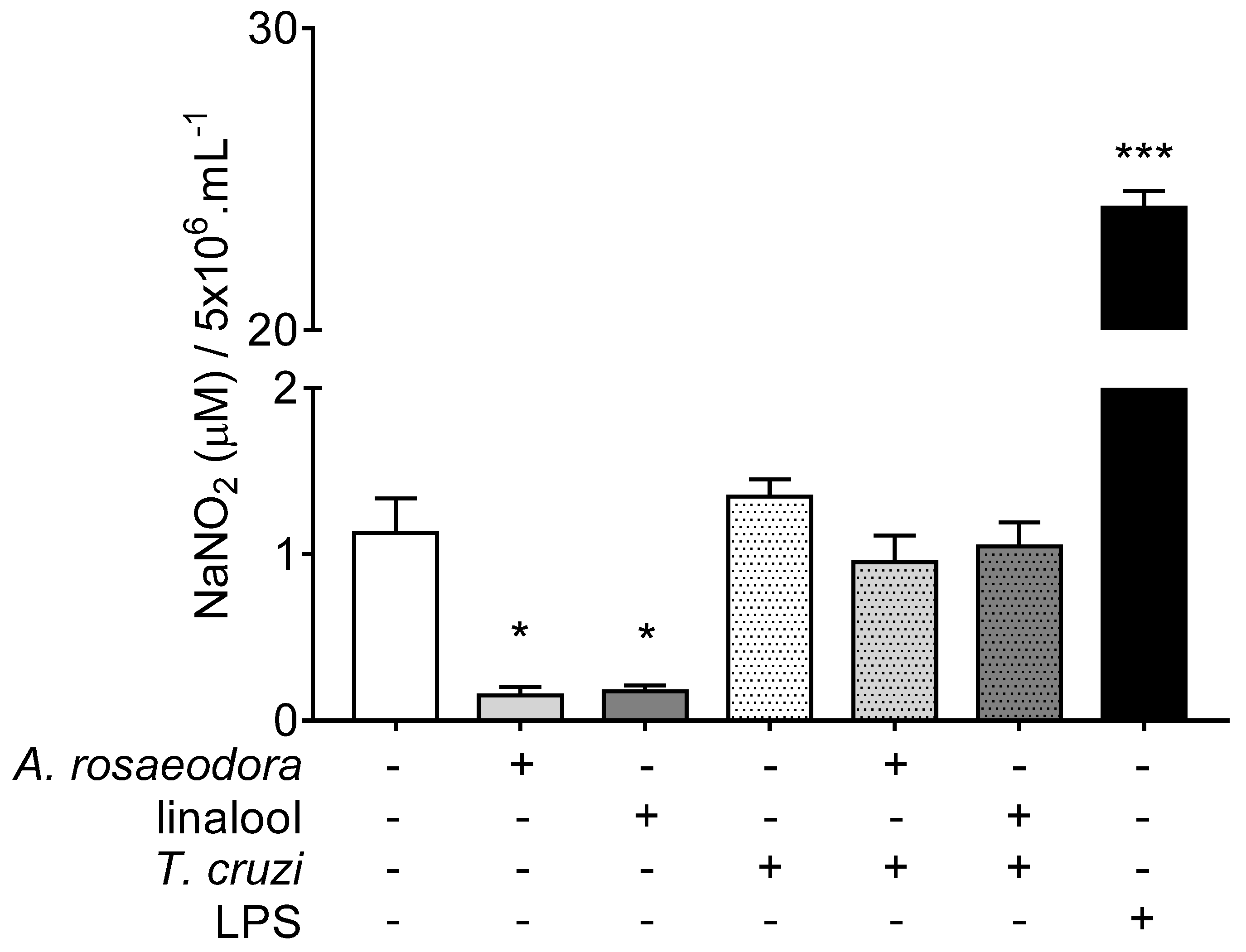
2.5. Nitrite Quantification in T. Cruzi-Infected Peritoneal Macrophages Treated with A. rosaeodora Essential Oil and Linalool
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Plant Material
4.3. Essential Oil Extraction
4.4. Physical-Chemical Analysis of Essential Oil
4.5. Gas Chromatography–Mass Spectrometry (GC–MS)
4.6. Bacteria from Marine Enviroment
4.7. Bacterial Strains and Culture Conditions
4.8. Antimicrobial Assays
4.9. Antioxidant Assay
4.10. Parasites
4.11. Anti-Epimastigote Assay
4.12. Animals and Ethical Statements
4.13. Peritoneal Macrophage Collection and Culture
4.14. Cytotoxicity Assay
4.15. Activitiy Against Intracellular Amastigotes and Selectivity Index (SI)
4.16. Nitrite Quantification
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial Resistance and the Alternative Resources with Special Emphasis on Plant-Based Antimicrobials-A Review. Plants 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.H.; Kota, B.P.; Razmovski, V.; Roufogalis, B.D. Herbal or natural medicines as modulators of peroxisome proliferator-activated receptors and related nuclear receptors for therapy of metabolic syndrome. Basic Clin. Pharmacol. Toxicol. 2005, 96. [Google Scholar] [CrossRef] [PubMed]
- Orchard, A.; van Vuuren, S. Commercial Essential Oils as Potential Antimicrobials to Treat Skin Diseases. Evid. Based Complementary Altern. Med. Ecam 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Morais, M.C.; Souza, J.V.; da Silva Maia Bezerra Filho, C.; Dolabella, S.S.; Sousa, D.P. Trypanocidal Essential Oils: A Review. Molecules 2020, 25, 4568. [Google Scholar] [CrossRef]
- Maia, J.G.S.; Andrade, E.H.A.; Couto, H.A.R.; Silva, A.C.M.; Marx, F.; Henke, C. Plant sources of amazon rosewood oil. Química Nova 2007, 30, 1906–1910. [Google Scholar] [CrossRef]
- May, P.H.; Barata, L.E.S. Rosewood exploitation in the Brazilian Amazon: Options for sustainable production. Econ. Bot. 2004, 58, 257–265. [Google Scholar] [CrossRef]
- Maia, J.G.S.; Zoghbi, M.G.B.; Andrade, E.H.A. Essential Oils of Aeollanthus suaveolens Matt. ex Spreng. J. Essent. Oil Res. 2003, 15, 86–87. [Google Scholar] [CrossRef]
- Maia, J.G.S.; Andrade, E.H.A. Database of the Amazon aromatic plants and their essential oils. Química Nova 2009, 32, 595–622. [Google Scholar] [CrossRef] [Green Version]
- Pattnaik, S.; Subramanyam, V.R.; Bapaji, M.; Kole, C.R. Antibacterial and antifungal activity of aromatic constituents of essential oils. Microbios 1997, 89, 39. [Google Scholar]
- Herman, A.; Tambor, K.; Herman, A. Linalool Affects the Antimicrobial Efficacy of Essential Oils. Curr. Microbiol. 2016, 72, 165–172. [Google Scholar] [CrossRef]
- Seol, G.H.; Kang, P.; Lee, H.S.; Seol, G.H. Antioxidant activity of linalool in patients with carpal tunnel syndrome. BMC Neurol. 2016, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bero, J.; Kpoviessi, S.; Quetin-Leclercq, J. Anti-Parasitic Activity of Essential Oils and their Active Constituents against Plasmodium, Trypanosoma and Leishmania. Nov. Plant Bioresour. 2014, 455–469. [Google Scholar] [CrossRef]
- Soeur, J.; Marrot, L.; Perez, P.; Iraqui, I.; Kienda, G.; Dardalhon, M.; Meunier, J.R.; Averbeck, D.; Huang, M.E. Selective cytotoxicity of Aniba rosaeodora essential oil towards epidermoid cancer cells through induction of apoptosis. Mutat. Res. 2011, 718. [Google Scholar] [CrossRef]
- Da Silva, Y.C.; Silva, E.M.S.; Fernandes, N.S.; Lopes, N.L.; Orlandi, P.P.; Nakamura, C.V.; Costa, E.V.; da Veiga Junior, V.F. Antimicrobial substances from Amazonian Aniba (Lauraceae) species. Nat. Prod. Res. 2019, 1–4. [Google Scholar] [CrossRef]
- Sampaio Lde, F.; Maia, J.G.; de Parijos, A.M.; de Souza, R.Z.; Barata, L.E. Linalool from rosewood (Aniba rosaeodora Ducke) oil inhibits adenylate cyclase in the retina, contributing to understanding its biological activity. Phytother. Res. 2012, 26, 73–77. [Google Scholar] [CrossRef]
- D’ACAMPORA Zellner, B.; Lo Presti, M.; Barata, L.E.; Dugo, P.; Dugo, G.; Mondello, L. Evaluation of leaf-derived extracts as an environmentally sustainable source of essential oils by using gas chromatography-mass spectrometry and enantioselective gas chromatography-olfactometry. Anal. Chem. 2006, 78. [Google Scholar] [CrossRef]
- Almeida, M.R.; Fidelis, C.H.; Barata, L.E.; Poppi, R.J. Classification of Amazonian rosewood essential oil by Raman spectroscopy and PLS-DA with reliability estimation. Talanta 2013, 117, 305–311. [Google Scholar] [CrossRef]
- Ye, M.; Sun, M.; Huang, D.; Zhang, Z.; Zhang, H.; Zhang, S.; Hu, F.; Jiang, X.; Jiao, W. A review of bacteriophage therapy for pathogenic bacteria inactivation in the soil environment. Environ. Int. 2019, 129. [Google Scholar] [CrossRef]
- Holetz, F.B.; Pessini, G.L.; Sanches, N.R.; Cortez, D.A.; Nakamura, C.V.; Filho, B.P. Screening of some plants used in the Brazilian folk medicine for the treatment of infectious diseases. Mem. Inst. Oswaldo Cruz. 2002, 97, 1027–1031. [Google Scholar] [CrossRef] [Green Version]
- Rosato, A.; Piarulli, M.; Corbo, F.; Muraglia, M.; Carone, A.; Vitali, M.E.; Vitali, C. In vitro synergistic antibacterial action of certain combinations of gentamicin and essential oils. Curr. Med. Chem. 2010, 17, 3289–3295. [Google Scholar] [CrossRef]
- Jabir, M.S.; Taha, A.A.; Sahib, U.I. Linalool loaded on glutathione-modified gold nanoparticles: A drug delivery system for a successful antimicrobial therapy. Artif. Cells Nanomed Biotechnol. 2018, 46, 345–355. [Google Scholar] [CrossRef]
- Vainstein, A.; Lewinsohn, E.; Pichersky, E.; Weiss, D. Floral fragrance. New inroads into an old commodity. Plant Physiol. 2001, 127, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Christensson, J.B.; Matura, M.; Gruvberger, B.; Bruze, M.; Karlberg, A.T. Linalool—A significant contact sensitizer after air exposure. Contact Dermat. 2010, 62. [Google Scholar] [CrossRef] [PubMed]
- Bråred Christensson, J.; Andersen, K.E.; Bruze, M.; Johansen, J.D.; Garcia-Bravo, B.; Gimenez Arnau, A.; Goh, C.L.; Nixon, R.; White, I.R. Air-oxidized linalool: A frequent cause of fragrance contact allergy. Contact Dermat. 2012, 67. [Google Scholar] [CrossRef] [PubMed]
- Audrain, H.; Kenward, C.; Lovell, C.R.; Green, C.; Ormerod, A.D.; Sansom, J.; Chowdhury, M.M.; Cooper, S.M.; Johnston, G.A.; Wilkinson, M.; et al. Allergy to oxidized limonene and linalool is frequent in the U.K. Br. J. Dermatol. 2014, 171. [Google Scholar] [CrossRef]
- Martins, F.J.; Caneschi, C.A.; Vieira, J.L.; Barbosa, W.; Raposo, N.R. Antioxidant activity and potential photoprotective from amazon native flora extracts. J. Photochem. Photobiol. B Biol. 2016, 161. [Google Scholar] [CrossRef]
- Oliveira de Souza, L.I.; Bezzera-Silva, P.C.; do Amaral Ferraz Navarro, D.M.; da Silva, A.G.; Dos Santos Correia, M.T.; da Silva, M.V.; de Figueiredo, R.C.B.Q. The chemical composition and trypanocidal activity of volatile oils from Brazilian Caatinga plants. Biomed. Pharmacother. 2017, 96. [Google Scholar] [CrossRef]
- Dos Santos Sales, V.; Monteiro, Á.B.; Delmondes, G.A.; do Nascimento, E.P.; Sobreira Dantas Nóbrega de Figuêiredo, F.R.; de Souza Rodrigues, C.K.; Evangelista de Lacerda, J.F.; Fernandes, C.N.; Barbosa, M.O.; Brasil, A.X.; et al. Antiparasitic activity and essential oil chemical analysis of the piper tuberculatum jacq fruit. Iran. J. Pharm. Res. IJPR 2018, 17, 268. [Google Scholar]
- Giongo, J.L.; Vaucher, R.A.; Da Silva, A.S.; Oliveira, C.B.; de Mattos, C.B.; Baldissera, M.D.; Sagrillo, M.R.; Monteiro, S.G.; Custódio, D.L.; Souza de Matos, M.; et al. Trypanocidal activity of the compounds present in Aniba canelilla oil against Trypanosoma evansi and its effects on viability of lymphocytes. Microb. Pathog. 2017, 103. [Google Scholar] [CrossRef]
- Jackson, Y.; Wyssa, B.; Chappuis, F. Tolerance to Nifurtimox and Benznidazole in Adult Patients With Chronic Chagas’ Disease. J. Antimicrob. Chemother. 2020, 75. [Google Scholar] [CrossRef] [Green Version]
- Izumi, E.; Ueda-Nakamura, T.; Veiga, V.F.; Pinto, A.C.; Nakamura, C.V. Terpenes from Copaifera demonstrated in vitro antiparasitic and synergic activity. J. Med. Chem. 2012, 55. [Google Scholar] [CrossRef]
- Santoro, G.F.; Cardoso, M.G.; Guimarães, L.G.; Mendonça, L.Z.; Soares, M.J. Trypanosoma cruzi: Activity of essential oils from Achillea millefolium L., Syzygium aromaticum L. and Ocimum basilicum L. on epimastigotes and trypomastigotes. Exp. Parasitol. 2007, 116, 283–290. [Google Scholar] [CrossRef]
- Villamizar, L.H.; Cardoso, M.G.; Andrade, J.; Teixeira, M.L.; Soares, M.J. Linalool, a Piper aduncum essential oil component, has selective activity against Trypanosoma cruzi trypomastigote forms at 4 °C. Memórias Do Inst. Oswaldo Cruz. 2017, 112, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Suman Gupta, N. Visceral leishmaniasis: Experimental models for drug discovery. Indian J. Med. Res. 2011, 133, 27–39. [Google Scholar]
- Do Socorro S Rosa, M.d.S.; Mendonça-Filho, R.R.; Bizzo, H.R.; de Almeida Rodrigues, I.; Soares, R.M.; Souto-Padrón, T.; Alviano, C.S.; Lopes, A.H. Antileishmanial activity of a linalool-rich essential oil from Croton cajucara. Antimicrob. Agents Chemother. 2003, 47. [Google Scholar] [CrossRef] [Green Version]
- Dutra, F.L.; Oliveira, M.M.; Santos, R.S.; Silva, W.S.; Alviano, D.S.; Vieira, D.P.; Lopes, A.H. Effects of linalool and eugenol on the survival of Leishmania (L.) infantum chagasi within macrophages. Acta Trop. 2016, 164, 69–76. [Google Scholar] [CrossRef]
- Vespa, G.N.; Cunha, F.Q.; Silva, J.S. Nitric oxide is involved in control of Trypanosoma cruzi-induced parasitemia and directly kills the parasite in vitro. Infect. Immun. 1994, 62. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.G.; Kim, S.M.; Min, J.H.; Kwon, O.K.; Park, M.H.; Park, J.W.; Ahn, H.I.; Hwang, J.Y.; Oh, S.R.; Lee, J.W.; et al. Anti-inflammatory effects of linalool on ovalbumin-induced pulmonary inflammation. Int. Immunopharmacol. 2019, 74. [Google Scholar] [CrossRef]
- Peana, A.T.; Marzocco, S.; Popolo, A.; Pinto, A. (-)-Linalool inhibits in vitro NO formation: Probable involvement in the antinociceptive activity of this monoterpene compound. Life Sci. 2006, 78. [Google Scholar] [CrossRef]
- Salfinger, Y.; Tortorello, M.L. Compendium of Methods for the Microbiological Examination of Foods; American Public Health Association (APHA): Washington, DC, USA, 1992. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI: Wayne, PA, USA, 2009; p. 99. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- El Babili, F.; Bouajila, J.; Souchard, J.P.; Bertrand, C.; Bellvert, F.; Fouraste, I.; Moulis, C.; Valentin, A. Oregano: Chemical analysis and evaluation of its antimalarial, antioxidant, and cytotoxic activities. J. Food Sci. 2011, 76. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.G.; Webby, R.F.; Markham, K.R.; Mitchell, K.A.; Da Cunha, A.P. Age-induced diminution of free radical scav-enging capacity in bee pollens and the contribution of constituent flavonoids. J. Agric. Food. Chem. 2003, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rottini, M.M.; Amaral, A.C.F.; Ferreira, J.L.P.; Oliveira, E.S.C.; Silva, J.R.A.; Taniwaki, N.N.; Dos Santos, A.R.; Almeida-Souza, F.; de Souza, C.S.F.; Calabrese, K.S. Endlicheria bracteolata (Meisn.) essential oil as a weapon against Leishmania amazonensis: In vitro assay. Molecules 2019, 24, 2525. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, V.D.; Almeida-Souza, F.; Teles, A.M.; Neto, P.A.; Mondego-Oliveira, R.; Mendes Filho, N.E.; Taniwaki, N.N.; Abreu-Silva, A.L.; Calabrese, K.d.S.; Mouchrek Filho, V.E. Chemical composition of Ocimum canum Sims. essential oil and the antimicrobial, antiprotozoal and ultrastructural alterations it induces in Leishmania amazonensis promastigotes. Ind. Crop. Prod. 2018, 119, 201–208. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65. [Google Scholar] [CrossRef]
- Oliveira, I.S.S.; Colares, A.V.; Cardoso, F.O.; Tellis, C.J.M.; Chagas, M.S.S.; Behrens, M.D.; Calabrese, K.S.; Almeida-Souza, F.; Abreu-Silva, A.L. Vernonia polysphaera baker: Anti-inflammatory activity in vivo and inhibitory effect in lps-stimulated RAW 264.7 cells. PLoS ONE 2019, 14, e0225275. [Google Scholar] [CrossRef] [Green Version]
- Teles, A.M.; Rosa, T.; Mouchrek, A.N.; Abreu-Silva, A.L.; Calabrese, K.D.S.; Almeida-Souza, F. Cinnamomum zeylanicum, Origanum vulgare, and Curcuma longa essential oils: Chemical composition, antimicrobial and antileishmanial activity. Evid Based Complement Alternat. Med. 2019, 2019, 2421695. [Google Scholar] [CrossRef] [Green Version]
- Almeida-Souza, F.; Silva, V.D.D.; Silva, G.X.; Taniwaki, N.N.; Hardoim, D.J.; Buarque, C.D.; Abreu-Silva, A.L.; Calabrese, K.D.S. 1,4-Disubstituted-1,2,3-Triazole compounds induce ultrastructural alterations in leishmania amazonensis promastigote: An in vitro antileishmanial and in silico pharmacokinetic study. Int. J. Mol. Sci. 2020, 21, 6839. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Compounds | Retention Time (min) | Peak Area (%) 1 |
|---|---|---|---|
| 1 | α-terpinolene | 8.361 | 3.37 |
| 2 | linalool cis-oxide | 8.702 | 3.03 |
| 3 | linalool | 9.177 | 93.60 |
| Compounds/Antibiotics | Bacteria Strain | ||||
|---|---|---|---|---|---|
| Aeromonas caviae | Aeromonas hydrophila | Enterococcus faecalis | Klebsiella pneumoniae | Providencia stuartii | |
| A. rosaeodora essential oil | 22.33 ± 0.577 | 11.33 ± 0.577 | 21.33 ± 0.577 | 10.33 ± 0.577 | 9.33 ± 0.577 |
| linalool | 7.33 ± 0.577 | 11.66 ± 0.577 | 13.33 ± 0.577 | 11.66 ± 0.577 | 11.33 ± 0.577 |
| pipemidic acid | 22.33 ± 0.577 | 21.66 ± 0.577 | 18.66 ± 0.577 | 22.33±0.577 | 22.33 ± 0.577 |
| ampicillin | 0 ± 0.00 | 0 ± 0.00 | 15.66 ± 0.577 | 0 ± 0.00 | 0 ± 0.00 |
| cefotaxime | 24.67 ± 0.577 | 23.66 ± 0.577 | 17.66 ± 0.577 | 17.33 ± 0.577 | 25.33 ± 0.577 |
| cefoxitin | 0 ± 0.00 | 13.66 ± 0.577 | 22.66 ± 0.577 | 8.33 ± 0.577 | 15.33 ± 0.577 |
| chloramphenicol | 23.33 ± 1.155 | 20.66 ± 0.577 | 24.66 ± 0.577 | 13.66 ± 0.577 | 13.33 ± 0.577 |
| erythromycin | 0 ± 0.00 | 0 ± 0.00 | 25.66 ± 1.527 | 0 ± 0.00 | 0 ± 0.00 |
| gentamycin | 17.66 ± 0.577 | 14.66 ± 0.577 | 22.33 ± 0.577 | 14.33 ± 0.577 | 14.33 ± 0.577 |
| lincomycin | 0 ± 0.00 | 0 ± 0.00 | 29.66 ± 0.577 | 0 ± 0.00 | 0 ± 0.00 |
| oxacillin | 0 ± 0.00 | 0 ± 0.00 | 18.66 ± 0.577 | 0 ± 0.00 | 0 ± 0.00 |
| sulfazotrin | 21.66 ± 0.577 | 19.66 ± 0.577 | 21 ± 1.00 | 20.33 ± 0.577 | 21.33 ± 0.577 |
| tetracycline | 10.33 ± 0.577 | 22.66 ± 0.577 | 19.66 ± 0.577 | 0 ± 0.00 | 18.33 ± 0.577 |
| vancomycin | 0 ± 0.00 | 0 ± 0.00 | 20.66 ± 0.577 | 0 ± 0.00 | 0 ± 0.00 |
| Antimicrobial assay | Compounds | Bacteria Strain | |||
|---|---|---|---|---|---|
| Escherichia coli | Staphylococcus aureus | Pseudomonas aeruginosa | Salmonella choleraesuis | ||
| Inhibition zones (mm) by disc-diffusion | A. rosaeodora | 15.0 ± 0.00 | 18.0 ± 0.00 | 13.0 ± 0.00 | 14.0 ± 0.00 |
| linalool | 13.0 ± 0.00 | 15.0 ± 0.00 | 12.0 ± 0.00 | 12.0 ± 0.00 | |
| gentamycin | 14.0 ± 0.00 | 20.5 ± 0.70 | 17.0 ± 0.00 | n.d. | |
| penicillin | n.d. | n.d. | n.d. | 18.5 ± 0.70 | |
| MIC (μg/mL) | A. rosaeodora | 350.0 ± 0.00 | 250.0 ± 0.00 | 450.0 ± 0.00 | 400.0 ± 0.00 |
| linalool | 650.0 ± 0.00 | 550.0 ± 0.00 | 650.0 ± 0.00 | 650.0 ± 0.00 | |
| amoxicillin | 16.0 ± 0.00 | 8.0 ± 0.00 | n.d. | n.d. | |
| gentamycin | n.d. | 2.0 ± 0.00 | n.d. | 8.0 ± 0.00 | |
| polymyxin B | n.d. | n.d. | 16.0 ± 0.00 | n.d. | |
| Essential oil/Compounds | Citotoxicity CC50 (µg/mL) | T. cruzi IC50 (µg/mL) | SI | |
|---|---|---|---|---|
| Peritoneal Macrophage | Epimastigote | Intracellular Amastigote | ||
| A. rosaeodora | >1000 | 150.5 ± 1.08 | 911.6 ± 1.15 | >1.0 |
| linalool | >1000 | 198.6 ± 1.12 | 249.6 ± 1.18 | >4.0 |
| benznidazole | 162.0 ± 1.11 | 1.805 ± 1.13 | 0.4820 ± 1.17 | 336.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teles, A.M.; Silva-Silva, J.V.; Fernandes, J.M.P.; Calabrese, K.d.S.; Abreu-Silva, A.L.; Marinho, S.C.; Mouchrek, A.N.; Filho, V.E.M.; Almeida-Souza, F. Aniba rosaeodora (Var. amazonica Ducke) Essential Oil: Chemical Composition, Antibacterial, Antioxidant and Antitrypanosomal Activity. Antibiotics 2021, 10, 24. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10010024
Teles AM, Silva-Silva JV, Fernandes JMP, Calabrese KdS, Abreu-Silva AL, Marinho SC, Mouchrek AN, Filho VEM, Almeida-Souza F. Aniba rosaeodora (Var. amazonica Ducke) Essential Oil: Chemical Composition, Antibacterial, Antioxidant and Antitrypanosomal Activity. Antibiotics. 2021; 10(1):24. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10010024
Chicago/Turabian StyleTeles, Amanda Mara, João Victor Silva-Silva, Juan Matheus Pereira Fernandes, Kátia da Silva Calabrese, Ana Lucia Abreu-Silva, Silvio Carvalho Marinho, Adenilde Nascimento Mouchrek, Victor Elias Mouchrek Filho, and Fernando Almeida-Souza. 2021. "Aniba rosaeodora (Var. amazonica Ducke) Essential Oil: Chemical Composition, Antibacterial, Antioxidant and Antitrypanosomal Activity" Antibiotics 10, no. 1: 24. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10010024