
Effect of Subinhibitory Concentrations of Antibiotics and Disinfectants on ISAba-Mediated Inactivation of Lipooligosaccharide Biosynthesis Genes in Acinetobacter baumannii


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Development of an Assay for Quantifying ISAba Insertion Frequency in LOS Biosynthesis Genes
2.2. Characterization of ISAba Insertion Frequencies in LOS Biosynthesis Genes in Different A. baumannii Strains
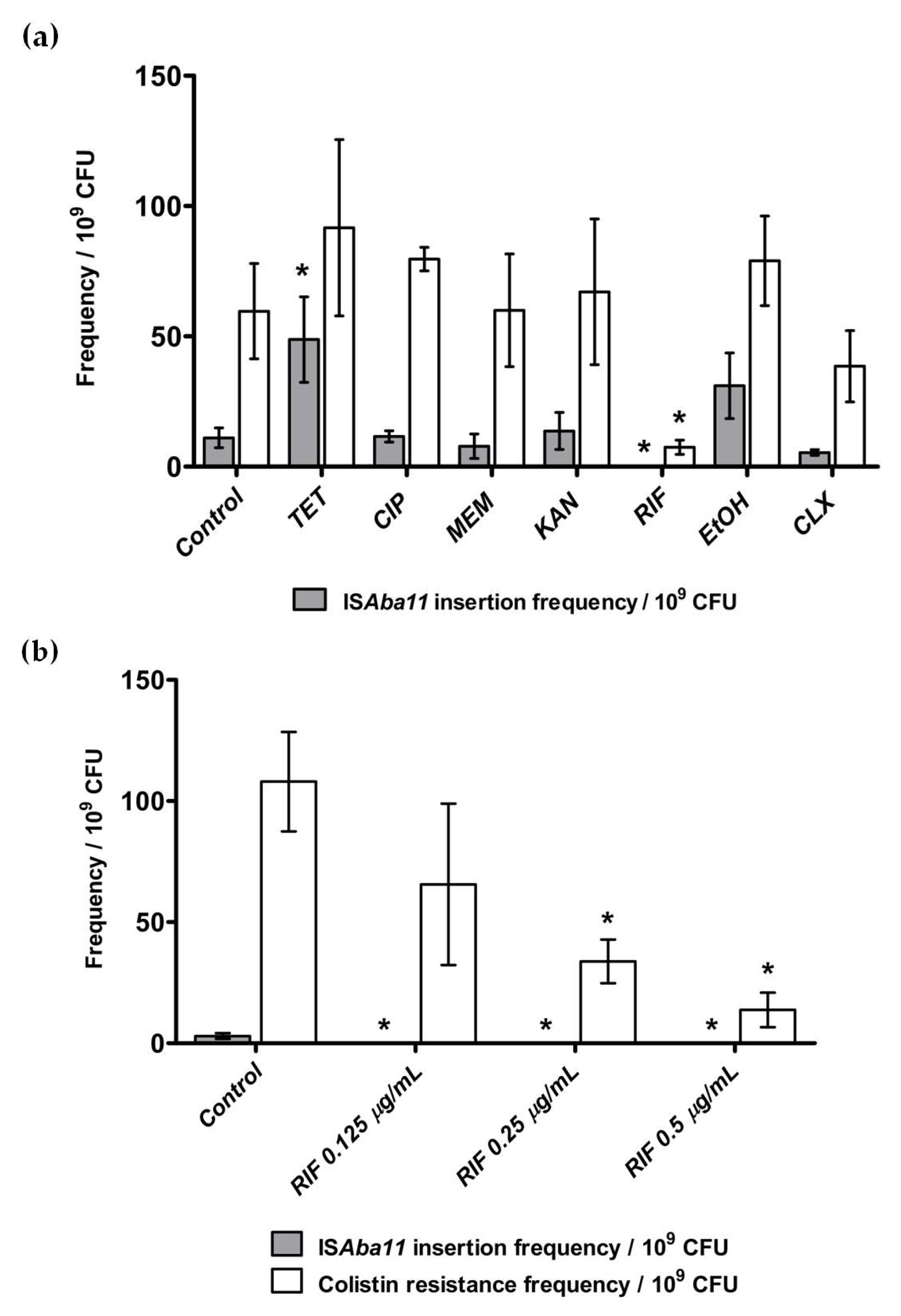
2.3. Effect of Antibiotics and Disinfectants on ISAba11 Insertion Frequencies in lpxC
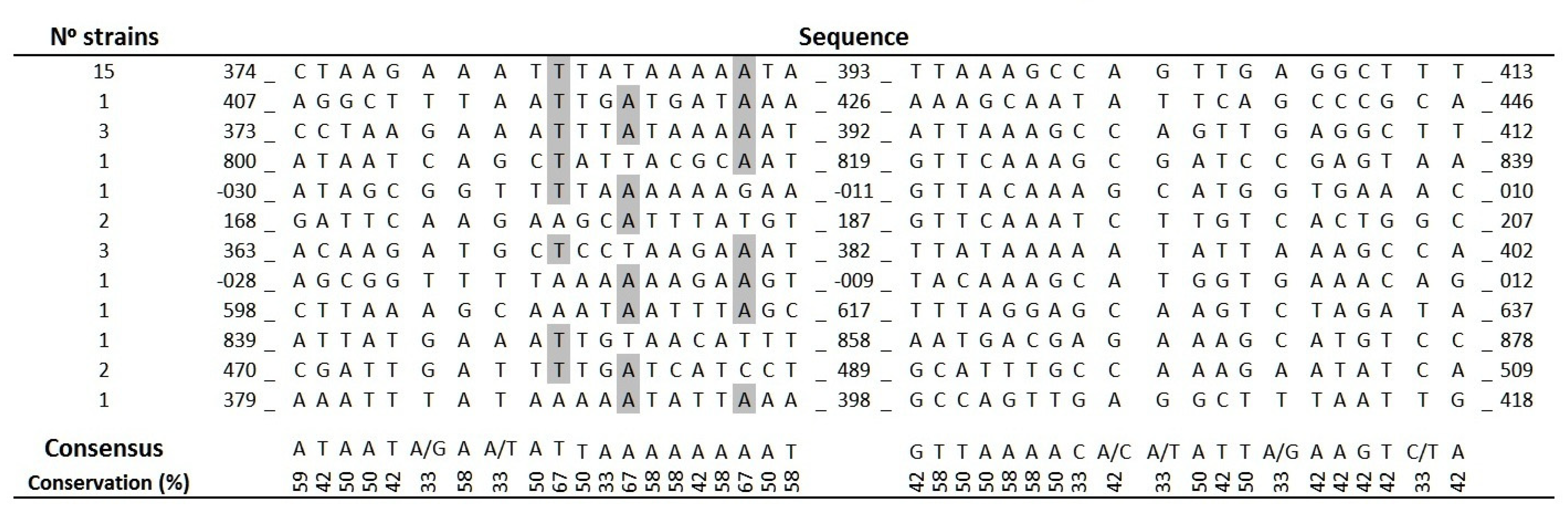
2.4. Analysis of ISAba11 Insertion Sites in lpxC
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Antibiotics, Disinfectants and Minimum Inhibitory Concentration Determination
4.3. Quantification of ISAba Insertion in LOS Biosynthesis Genes
4.4. Sequencing of ISAba11 Insertions in lpxC
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McConnell, M.J.; Actis, L.; Pachon, J. Acinetobacter baumannii: Human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol. Rev. 2013, 37, 130–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, J.; Zander, E.; Stefanik, D.; Higgins, P.G.; Roca, I.; Vila, J.; McConnell, M.; Cisneros, J.M.; Seifert, H. High incidence of pandrug-resistant Acinetobacter baumannii isolates collected from patients with ventilator-associated pneumonia in Greece, Italy and Spain as part of the MagicBullet clinical trial. J. Antimicrob. Chemother. 2017, 72, 3277–3282. [Google Scholar] [CrossRef]
- Moffatt, J.H.; Harper, M.; Boyce, J.D. Mechanisms of Polymyxin Resistance. Adv. Exp. Med. Biol. 2019, 1145, 55–71. [Google Scholar]
- Andrade, F.F.; Silva, D.; Rodrigues, A.; Pina-Vaz, C. Colistin Update on Its Mechanism of Action and Resistance, Present and Future Challenges. Microorganisms 2020, 8, 1716. [Google Scholar] [CrossRef]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.F.; Vinogradov, E.; Seemann, T.; Henry, R.; Crane, B.; Michael, F.S.; Cox, A.D.; et al. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrob. Agents Chemother. 2010, 54, 4971–4977. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, J.H.; Harper, M.; Adler, B.; Nation, R.; Li, J.; Boyce, J.D. Insertion sequence ISAba11 is involved in colistin resistance and loss of lipopolysaccharide in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 3022–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Quintanilla, M.; Pulido, M.R.; Moreno-Martínez, P.; Martin-Pena, R.; López-Rojas, R.; Pachón, J.; McConnell, M.J. Activity of host antimicrobials against multidrug-resistant Acinetobacter baumannii acquiring colistin resistance through loss of lipopolysaccharide. Antimicrob Agents Chemother. 2014, 58, 2972–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carretero-Ledesma, M.; García-Quintanilla, M.; Martín-Peña, R.; Pulido, M.R.; Pachón, J.; McConnell, M.J. Phenotypic changes associated with Colistin resistance due to Lipopolysaccharide loss in Acinetobacter baumannii. Virulence 2018, 9, 930–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaiarsa, S.; Bitar, I.; Comandatore, F.; Corbella, M.; Piazza, A.; Scaltriti, E.; Villa, L.; Postiglione, U.; Marone, P.; Nucleo, M.; et al. Can Insertion Sequences Proliferation Influence Genomic Plasticity? Comp. Anal. Of. Front. Microbiol. 2019, 10, 2080. [Google Scholar] [CrossRef] [PubMed]
- Toleman, M.A.; Walsh, T.R. Combinatorial events of insertion sequences and ICE in Gram-negative bacteria. FEMS Microbiol. Rev. 2011, 35, 912–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botelho, J.; Schulenburg, H. The Role of Integrative and Conjugative Elements in Antibiotic Resistance Evolution. Trends Microbiol. 2021, 29, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Guerin, E.; Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Erill, I.; Da Re, S.; Gonzalez-Zorn, B.; Barbé, J.; Ploy, M.; Mazel, D. The SOS response controls integron recombination. Science 2009, 324, 1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jara, L.M.; Cortés, P.; Bou, G.; Barbé, J.; Aranda, J. Differential roles of antimicrobials in the acquisition of drug resistance through activation of the SOS response in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 4318–4320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Quintanilla, M.; Carretero-Ledesma, M.; Moreno-Martínez, P.; Martin-Pena, R.; Pachón, J.; McConnell, M.J. Lipopolysaccharide loss produces partial colistin dependence and collateral sensitivity to azithromycin, rifampicin and vancomycin in Acinetobacter baumannii. Int. J. Antimicrob. Agents. 2015, 46, 696–702. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olmeda-López, H.; Corral-Lugo, A.; McConnell, M.J. Effect of Subinhibitory Concentrations of Antibiotics and Disinfectants on ISAba-Mediated Inactivation of Lipooligosaccharide Biosynthesis Genes in Acinetobacter baumannii. Antibiotics 2021, 10, 1259. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101259
Olmeda-López H, Corral-Lugo A, McConnell MJ. Effect of Subinhibitory Concentrations of Antibiotics and Disinfectants on ISAba-Mediated Inactivation of Lipooligosaccharide Biosynthesis Genes in Acinetobacter baumannii. Antibiotics. 2021; 10(10):1259. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101259
Chicago/Turabian StyleOlmeda-López, Héctor, Andrés Corral-Lugo, and Michael J. McConnell. 2021. "Effect of Subinhibitory Concentrations of Antibiotics and Disinfectants on ISAba-Mediated Inactivation of Lipooligosaccharide Biosynthesis Genes in Acinetobacter baumannii" Antibiotics 10, no. 10: 1259. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101259