
Investigation of the Genes Involved in the Outbreaks of Escherichia coli and Salmonella spp. in the United States


Abstract
:1. Introduction
2. Results
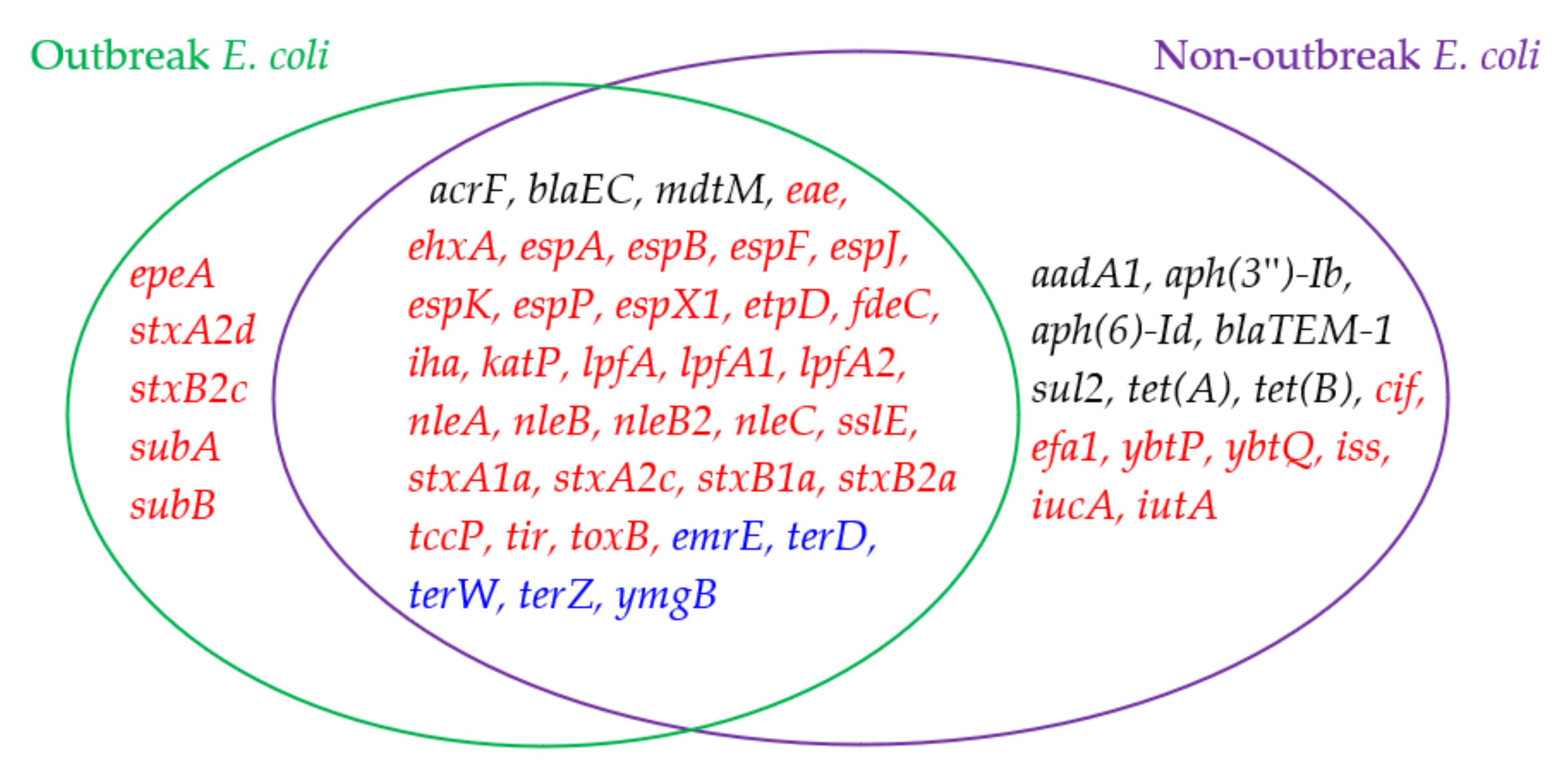
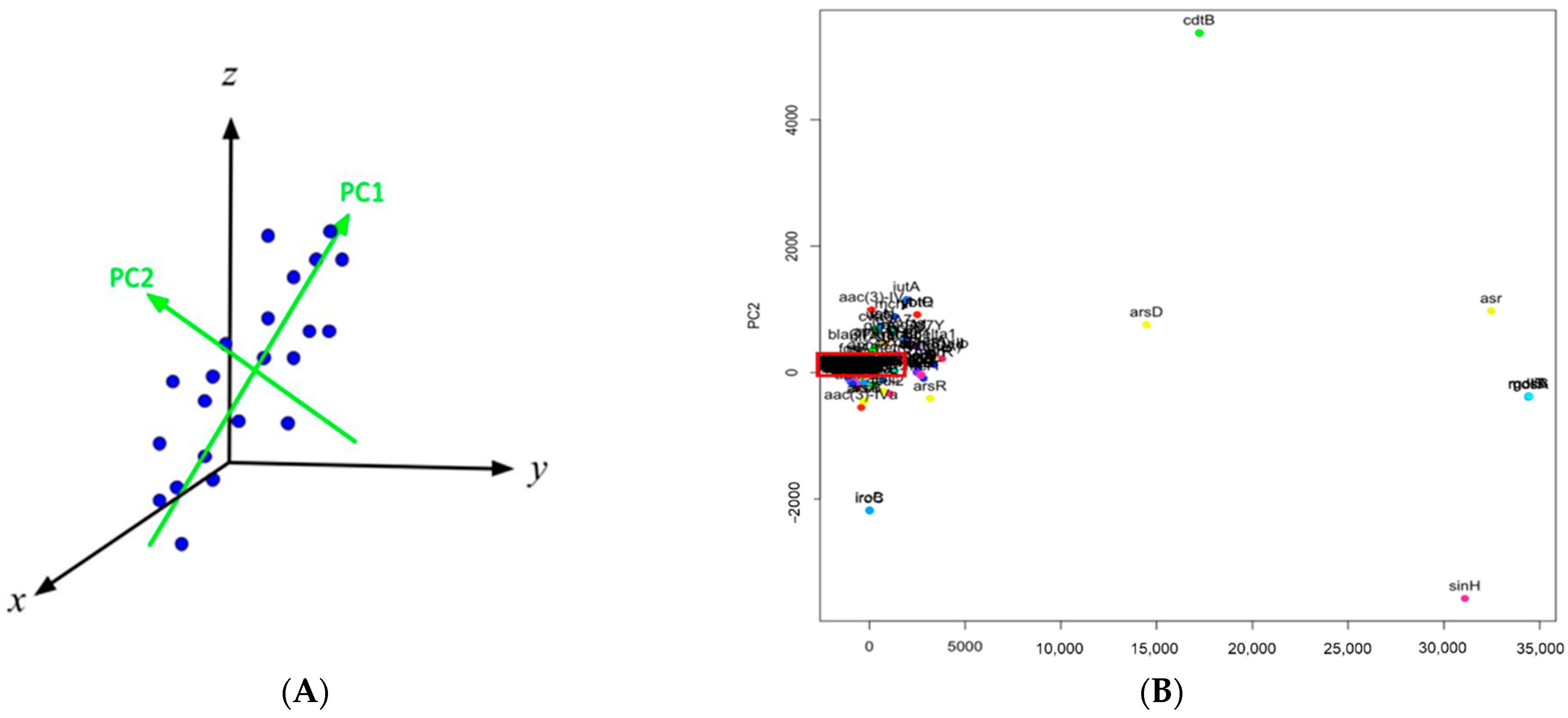
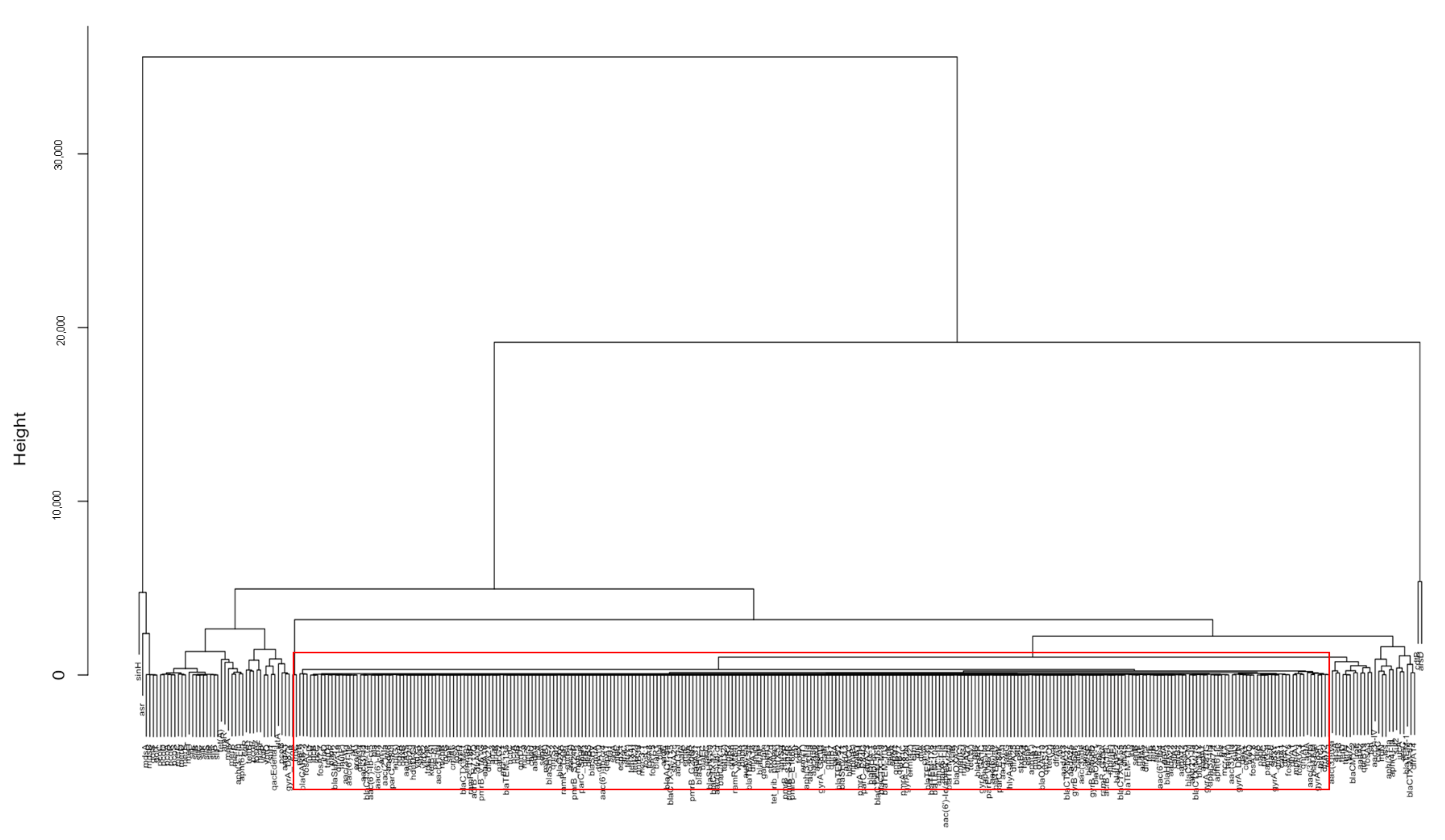
2.1. Analysis and Comparison of Genes Detected in the Outbreak and Non-Outbreak E. coli
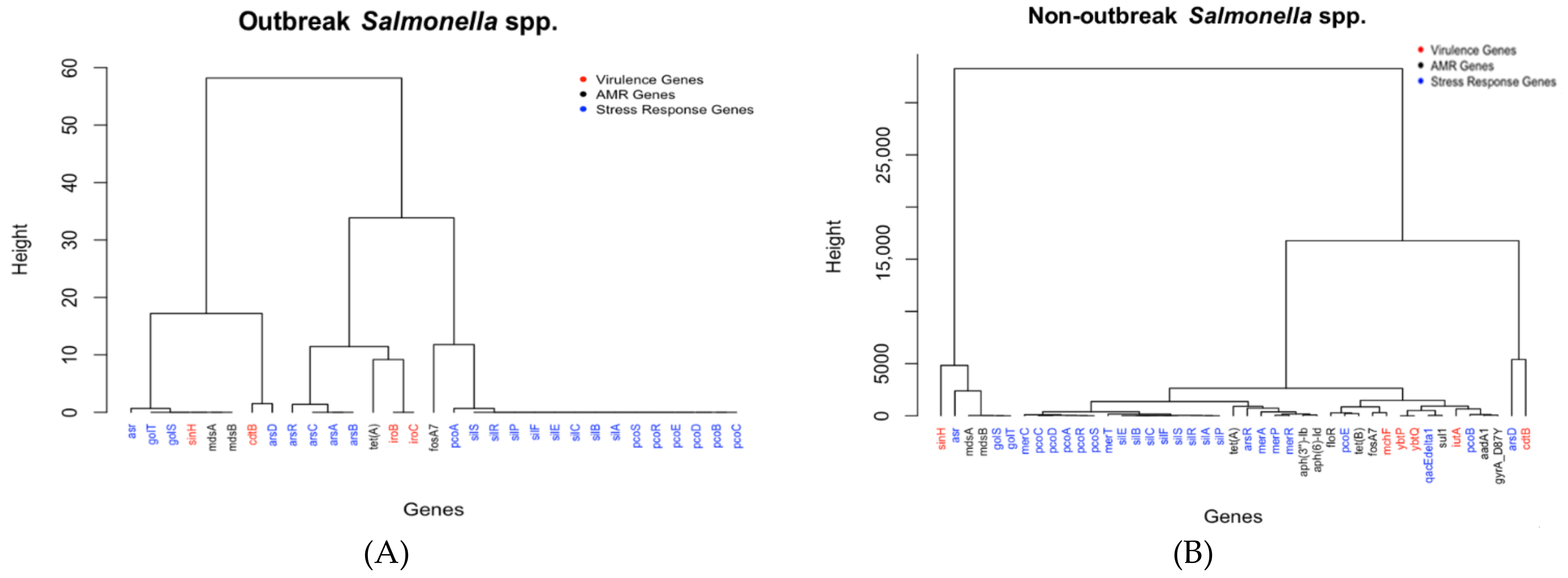
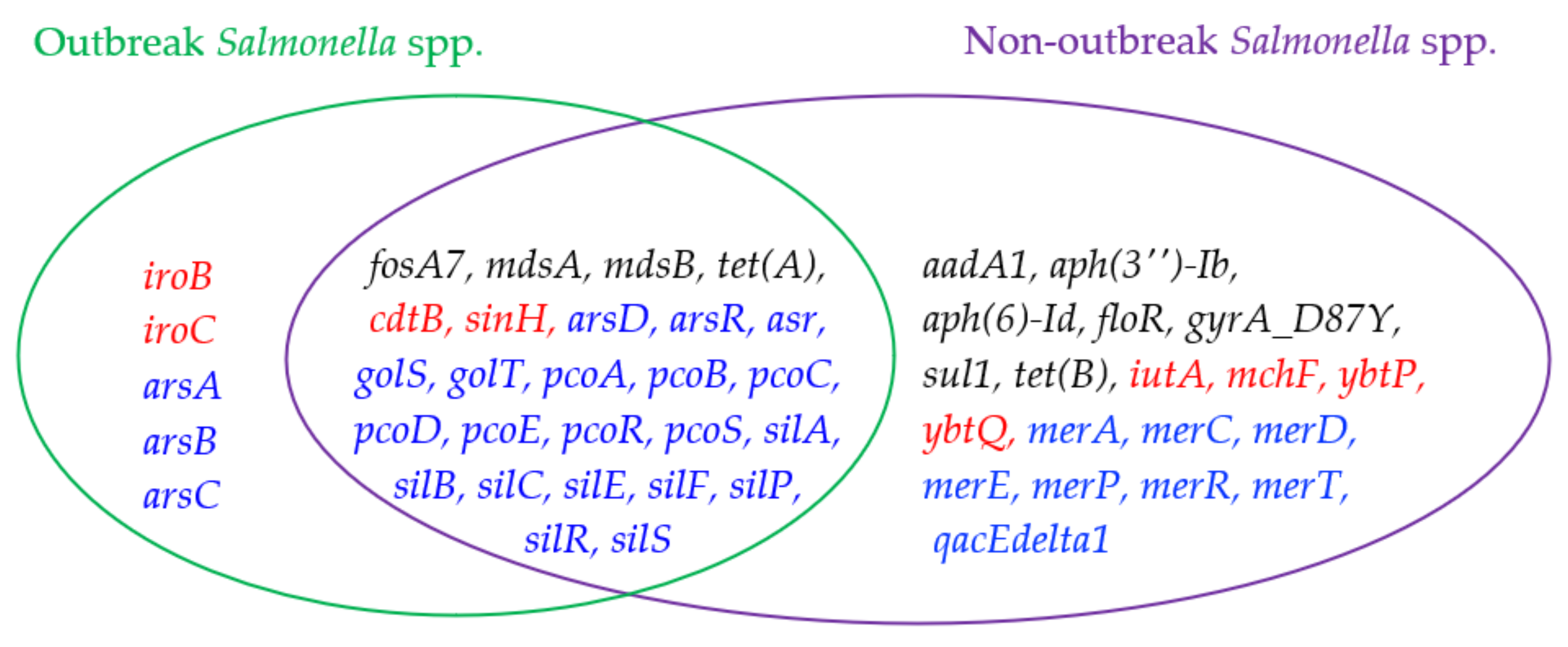
2.2. Analysis and Comparison of Genes Detected in the Outbreak and Non-Outbreak Salmonella spp.
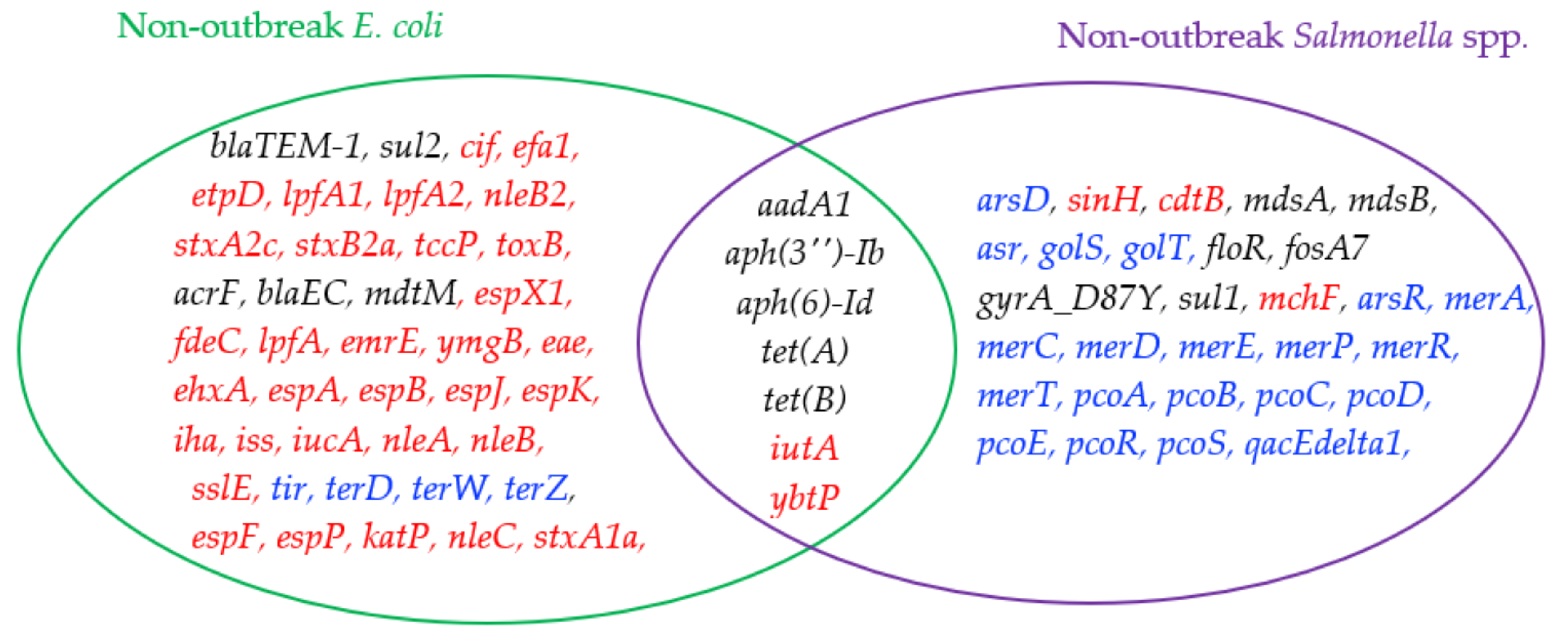
2.3. Comparison of Genes Detected in E. coli and Salmonella spp.
3. Discussion
3.1. Outlier Gene Difference between Outbreak and Non-Outbreak Pathogens
3.2. Outlier Gene Difference between Salmonella spp. and E. coli
3.3. Interaction between Antimicrobial Resistance Genes, Stress Response Genes, and Virulence Genes
4. Materials and Methods
4.1. Gene Data of E. coli and Salmonella spp. from NPIB Database
4.2. Identification of Important Genes via PCA and Hierarchical Clustering
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, J.L.; Fratamico, P.M. Emerging and Re-Emerging Foodborne Pathogens. Foodborne Pathog. Dis. 2018, 15, 12. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Most Common Foodborne Illnesses. Available online: https://www.fda.gov/files/food/published/Most-Common-Foodborne-Illnesses-%28PDF%29.pdf (accessed on 23 September 2021).
- Flanders, A. Economic Impact of Georgia Tomato Production Value Losses Due to the US Salmonella Outbreak. University of Georgia 2008. Center Report, CRO-08-17. Available online: http://hdl.handle.net/10724/18683 (accessed on 20 August 2021).
- Plumb, I.; Schwensohn, C.; Gieraltowski, L.; Tecle, S.; Schneider, Z.D.; Freiman, J.; Cote, A.; Noveroske, D.; Kolsin, J.; Brandenburg, J.; et al. Outbreak of Salmonella Newport Infections with Decreased Susceptibility to Azithromycin Linked to Beef Obtained in the United States and Soft Cheese Obtained in Mexico—United States, 2018–2019. Morb. Mortal. Wkly. Rep. 2019, 68, 713–717. [Google Scholar] [CrossRef] [Green Version]
- Vicente, K.J.; Christoffersen, K. The Walkerton Escherichia coli outbreak: A test of Rasmussen’s framework for risk management in a dynamic society. Theor. Issues Ergon. Sci. 2006, 7, 93–112. [Google Scholar] [CrossRef]
- Soborg, B.; Lassen, S.G.; Müller, L.; Jensen, T.; Ethelberg, S.; Mølbak, K.; Scheutz, F. A verocytotoxin-producing Escherichia coli outbreak with a surprisingly high risk of haemolytic uraemic syndrome, Denmark, September–October 2012. Eurosurveillance 2013, 18, 20350. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial resistance and virulence: A successful or deleterious association in the bacterial world? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [Green Version]
- Wuyts, V.; Denayer, S.; Roosens, N.H.; Mattheus, W.; Bertrand, S.; Marchal, K.; Dierick, K.; De Keersmaecker, S.C. Whole Genome Sequence Analysis of Salmonella Enteritidis PT4 Outbreaks from a National Reference Laboratory’s Viewpoint. PLoS Curr. 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, W.; Al-Khaldi, S.F.; Branham, W.S.; Han, T.; Fuscoe, J.C.; Han, J.; Foley, S.L.; Xu, J.; Fang, H.; Cerniglia, C.E.; et al. Microarray analysis of virulence gene profiles in Salmonella serovars from food/food animal environment. J. Infect. Dev. Ctries. 2011, 5, 94–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzuszkiewicz, E.; Thürmer, A.; Schuldes, J.; Leimbach, A.; Liesegang, H.; Meyer, F.D.; Boelter, J.; Petersen, H.; Gottschalk, G.; Daniel, R. Genome sequence analyses of two isolates from the recent Escherichia coli outbreak in Germany reveal the emergence of a new pathotype: Entero-Aggregative-Haemorrhagic Escherichia coli (EAHEC). Arch. Microbiol. 2011, 193, 883–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, M.K.; Li, L.; Nong, W.; Kwan, H.S. 2011 German Escherichia coli outbreak: Alignment-free whole-genome phylogeny by feature frequency profiles. Nat. Preced. 2011. [Google Scholar] [CrossRef] [Green Version]
- Decraene, V.; Phan, H.; George, R.; Wyllie, D.H.; Akinremi, O.; Aiken, Z.; Cleary, P.; Dodgson, A.; Pankhurst, L.; Crook, D.W.; et al. TRACE Investigators’ Group. A Large, Refractory Nosocomial Outbreak of Klebsiella pneumoniae Carbapenemase-Producing Escherichia coli Demonstrates Carbapenemase Gene Outbreaks Involving Sink Sites Require Novel Approaches to Infection Control. Antimicrob. Agents Chemother. 2018, 62, e01689-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.K.; Luo, H.; Zhang, Y.; Wang, B.; Gao, F. Pan-genomic analysis provides novel insights into the association of E. coli with human host and its minimal genome. Bioinformatics 2019, 35, 1987–1991. [Google Scholar] [CrossRef]
- Abram, K.; Udaondo, Z.; Bleker, C.; Wanchai, V.; Wassenaar, T.M.; Robeson, M.S., II; Ussery, D.W. Mash-based analyses of Escherichia coli genomes reveal 14 distinct phylogroups. Commun. Biol. 2021, 4, 117. [Google Scholar] [CrossRef]
- Vaughn, E.L.; Vo, Q.T.; Vostok, J.; Stiles, T.; Lang, A.; Brown, C.M.; Madoff, L. Linking Epidemiology and Whole-Genome Sequencing to Investigate Salmonella Outbreak, Massachusetts, USA, 2018. Emerg. Infect. Dis. 2020, 26, 1538–1541. [Google Scholar] [CrossRef]
- Zou, W.; Tang, H.; Zhao, W.; Meehan, J.; Foley, S.L.; Lin, W.J.; Chen, H.C.; Fang, H.; Nayak, R.; Chen, J.J. Data mining tools for Salmonella characterization: Application to gel-based fingerprinting analysis. BMC Bioinform. 2013, 14 (Suppl. S14), S15. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Deng, W.; Liu, S.; Yu, X.; Mustafa, G.R.; Chen, S.; He, L.; Ao, X.; Yang, Y.; Zhou, K.; et al. Presence of heavy metal resistance genes in Escherichia coli and Salmonella isolates and analysis of resistance gene structure in E. coli E308. J. Glob. Antimicrob. Resist. 2020, 21, 420–426. [Google Scholar] [CrossRef]
- Ibrahim, R.A.; Cryer, T.L.; Lafi, S.Q.; Basha, E.A.; Good, L.; Tarazi, Y.H. Identification of Escherichia coli from broiler chickens in Jordan, their antimicrobial resistance, gene characterization and the associated risk factors. BMC Vet. Res. 2019, 15, 159. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Meng, J.; McDermott, P.F.; Wang, F.; Yang, Q.; Cao, G.; Hoffmann, M.; Zhao, S. Presence of disinfectant resistance genes in Escherichia coli isolated from retail meats in the USA. J. Antimicrob. Chemother. 2014, 69, 2644–2649. [Google Scholar] [CrossRef] [Green Version]
- Maynard, C.; Bekal, S.; Sanschagrin, F.; Levesque, R.C.; Brousseau, R.; Masson, L.; Larivière, S.; Harel, J. Heterogeneity among virulence and antimicrobial resistance gene profiles of extraintestinal Escherichia coli isolates of animal and human origin. J. Clin. Microbiol. 2004, 42, 5444–5452. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, L.; Garneau, P.; Kurbasic, A.; Bruant, G.; Stegger, M.; Harel, J.; Jensen, K.S.; Brousseau, R.; Hammerum, A.M.; Frimodt-Møller, N. Microarray-based detection of extended virulence and antimicrobial resistance gene profiles in phylogroup B2 Escherichia coli of human, meat and animal origin. J. Med. Microbiol. 2011, 60 Pt 10, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.H.; Lee, J.H.; Park, S.H.; Song, M.O.; Park, S.H.; Jung, H.W.; Park, G.Y.; Choi, S.M.; Kim, M.S.; Chae, Y.Z.; et al. Antimicrobial resistance profiles among Escherichia coli strains isolated from commercial and cooked foods. Int. J. Food Microbiol. 2012, 159, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.; Fairbrother, J.M.; Bekal, S.; Sanschagrin, F.; Levesque, R.C.; Brousseau, R.; Masson, L.; Larivière, S.; Harel, J. Antimicrobial resistance genes in enterotoxigenic Escherichia coli O149:K91 isolates obtained over a 23-year period from pigs. Antimicrob. Agents Chemother. 2003, 47, 3214–3221. [Google Scholar] [CrossRef] [Green Version]
- Stubberfield, E.; AbuOun, M.; Sayers, E.; O’Connor, H.M.; Card, R.M.; Anjum, M.F. Use of whole genome sequencing of commensal Escherichia coli in pigs for antimicrobial resistance surveillance, United Kingdom, 2018. Eurosurveillance 2019, 50, 1900136. [Google Scholar] [CrossRef] [Green Version]
- Rosengren, L.B.; Waldner, C.L.; Reid-Smith, R.J. Associations between antimicrobial resistance phenotypes, antimicrobial resistance genes, and virulence genes of fecal Escherichia coli isolates from healthy grow-finish pigs. Appl. Environ. Microbiol. 2009, 75, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, E.L.; Mykytczuk, O.L.; Asensi, M.D.; Reis, E.M.; Ferraz, L.R.; Paula, F.L.; Ng, L.K.; Rodrigues, D.P. Clonality and antimicrobial resistance gene profiles of multidrug- resistant Salmonella enterica serovar infantis isolates from four public hospitals in Rio de Janeiro, Brazil. J. Clin. Microbiol. 2006, 44, 2767–2772. [Google Scholar] [CrossRef] [Green Version]
- Huehn, S.; La Ragione, R.M.; Anjum, M.; Saunders, M.; Woodward, M.J.; Bunge, C.; Helmuth, R.; Hauser, E.; Guerra, B.; Beutlich, J.; et al. Virulotyping and antimicrobial resistance typing of Salmonella enterica serovars relevant to human health in Europe. Foodborne Pathog. Dis. 2010, 7, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Mellou, K.; Gkova, M.; Panagiotidou, E.; Tzani, M.; Sideroglou, T.; Mandilara, G. Diversity and Resistance Profiles of Human Non-typhoidal Salmonella spp. in Greece, 2003–2020. Antibiotics 2021, 10, 983. [Google Scholar] [CrossRef]
- Matereke, L.T.; Okoh, A.I. Listeria monocytogenes Virulence, Antimicrobial Resistance and Environmental Persistence: A Review. Pathogens 2020, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, F.; van Schaik, W.; Sanguinetti, M.; Posteraro, B.; Torelli, R.; Le Bras, F.; Verneuil, N.; Zhang, X.; Giard, J.C.; Dhallui, A.; et al. AsrR is an oxidative stress sensing regulator modulating Enterococcus faecium opportunistic traits, antimicrobial resistance, and pathogenicity. PLoS Pathog. 2012, 8, e1002834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrzejewska, M.; Szczepańska, B.; Śpica, D.; Klawe, J.J. Prevalence, Virulence, and Antimicrobial Resistance of Campylobacter spp. in Raw Milk, Beef, and Pork Meat in Northern Poland. Foods 2019, 8, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Zheng, J.; Deng, T.; Peng, J.; Daniel, D.; Jia, Q.; Huang, Z. An Analysis of Antimicrobial Resistance of Clinical Pathogens from Historical Samples for Six Countries. Processes 2019, 7, 964. [Google Scholar] [CrossRef] [Green Version]
- Cui, K.; Gong, I.; Dong, A.; Yan, J.; Wang, M.; Huang, Z. Investigation of Virulence Genes Detected in Antimicrobial-Resistance Pathogens Isolates for Five Countries across the World. Processes 2020, 8, 1589. [Google Scholar] [CrossRef]
- Pei, R.; Zhang, L.; Duan, C.; Gao, M.; Feng, R.; Jia, Q.; Huang, Z. Investigation of Stress Response Genes in Antimicrobial Resistant Pathogens Sampled from Five Countries. Processes 2021, 9, 927. [Google Scholar] [CrossRef]
- Qin, J.S. Statistical process monitoring: Basics and beyond. J. Chemom. 2003, 17, 480–502. [Google Scholar] [CrossRef]
- Kourti, T. Application of latent variable methods to process control and multivariate statistical process control in industry. Int. J. Adapt. Control Signal Process 2005, 19, 213–246. [Google Scholar] [CrossRef]
- Arnau, V.; Mars, S.; Marín, I. Iterative Cluster Analysis of Protein Interaction Data. Bioinformatics 2005, 21, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Smith, K.; Hyndman, R. Characteristic-Based Clustering for Time Series Data. Data Min. Knowl. Disc. 2006, 13, 335–364. [Google Scholar] [CrossRef]
- Bar-Joseph, Z.; Demaine, E.D.; Gifford, D.K.; Hamel, A.M.; Jaakkola, T.S.; Srebro, N.H. K-ary clustering with optimal leaf ordering for gene expression data. Bioinformatics 2003, 19, 1070–1078. [Google Scholar] [CrossRef] [Green Version]
- Krüger, A.; Lucchesi, P.M. Shiga toxins and stx phages: Highly diverse entities. Microbiology 2015, 161 Pt 3, 451–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrametti, F.; Kresse, A.U.; Guzmán, C.A. Transcriptional regulation of the esp genes of enterohemorrhagic Escherichia coli. J. Bacteriol. 1999, 181, 3409–3418. [Google Scholar] [CrossRef] [Green Version]
- Coombes, B.K. Type III secretion systems in symbiotic adaptation of pathogenic and non-pathogenic bacteria. Trends Microbiol. 2009, 17, 89–94. [Google Scholar] [CrossRef]
- Dautin, N. Serine protease autotransporters of Enterobacteriaceae (SPATEs): Biogenesis and function. Toxins 2010, 2, 1179–1206. [Google Scholar] [CrossRef] [Green Version]
- Paton, A.W.; Paton, J.C. Escherichia coli Subtilase Cytotoxin. Toxins 2010, 2, 215–228. [Google Scholar] [CrossRef] [Green Version]
- Margolin, Y.; Barak, R.; Eisenbach, M. Arsenate arrests flagellar rotation in cytoplasm-free envelopes of bacteria. J. Bacteriol. 1994, 176, 5547–5549. [Google Scholar] [CrossRef] [Green Version]
- Leyton, D.L.; Sloan, J.; Hill, R.E.; Doughty, S.; Hartland, E.L. Transfer region of pO113 from enterohemorrhagic Escherichia coli: Similarity with R64 and identification of a novel plasmid-encoded autotransporter, EpeA. Infection and immunity. Infect. Immun. 2003, 71, 6307–6319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paton, A.W.; Srimanote, P.; Talbot, U.M.; Wang, H.; Paton, J.C. A new family of potent AB cytotoxins produced by Shiga toxigenic Escherichia coli. J. Exp. Med. 2004, 200, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bister, B.; Bischoff, D.; Nicholson, G.J.; Valdebenito, M.; Schneider, K.; Winkelmann, G.; Hantke, K.; Süssmuth, R.D. The structure of salmochelins: C-glucosylated enterobactins of Salmonella enterica. Biometals 2004, 17, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Mourão, J.; Rebelo, A.; Ribeiro, S.; Peixe, L.; Novais, C.; Antunes, P. Tolerance to arsenic contaminant among multidrug-resistant and copper-tolerant Salmonella successful clones is associated with diverse ars operons and genetic contexts. Environ. Microbiol. 2020, 22, 2829–2842. [Google Scholar] [CrossRef] [PubMed]
- AbuOun, M.; Suthers, P.F.; Jones, G.I.; Carter, B.R.; Saunders, M.P.; Maranas, C.D.; Woodward, M.J.; Anjum, M.F. Genome scale reconstruction of a Salmonella metabolic model: Comparison of similarity and differences with a commensal E. coli strain. J. Biol. Chem. 2009, 284, 29480–29488. [Google Scholar] [CrossRef] [Green Version]
- Winfield, M.D.; Groisman, E.A. Role of nonhost environments in the lifestyles of Salmonella and Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 3687–3694. [Google Scholar] [CrossRef] [Green Version]
- Ochman, H.; Groisman, E.A. The origin and evolution of species differences in Escherichia coli and Salmonella typhimurium. EXS 1994, 69, 479–493. [Google Scholar] [CrossRef]
- Iizumi, Y.; Sagara, H.; Kabe, Y.; Azuma, M.; Kume, K.; Ogawa, M.; Nagai, T.; Gillespie, P.G.; Sasakawa, C.; Handa, H. The enteropathogenic E. coli effector EspB facilitates microvillus effacing and antiphagocytosis by inhibiting myosin function. Cell Host Microbe 2007, 2, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; Kikuchi, T.; Tokunaga, T.; Iyoda, S.; Iguchi, A. Diversity of the Tellurite Resistance Gene Operon in Escherichia coli. Front. Microbiol. 2021, 12, 681175. [Google Scholar] [CrossRef]
- Chalmers, G.; Rozas, K.M.; Amachawadi, R.G.; Scott, H.M.; Norman, K.N.; Nagaraja, T.G.; Tokach, M.D.; Boerlin, P. Distribution of the pco Gene Cluster and Associated Genetic Determinants among Swine Escherichia coli from a Controlled Feeding Trial. Genes 2018, 9, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontel, L.B.; Audero, M.E.; Espariz, M.; Checa, S.K.; Soncini, F.C. GolS controls the response to gold by the hierarchical induction of Salmonella-specific genes that include a CBA efflux-coding operon. Mol. Biol. 2007, 66, 814–825. [Google Scholar] [CrossRef]
- Carrol, L.M.; Pierneef, R.; Mathole, M.; Matle, I. Genomic characterization of endemic and ecdemic non-typhoidal Salmonella enterica lineages circulating among animals and animal products in South Africa. Front. Microbiol. 2021, 12, 748611. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Collection Year | Type | Outbreak | fosA7 | mdsA | mdsB | … | tet(A) |
|---|---|---|---|---|---|---|---|---|
| 1 | 2019 | 2 | 0 | 0 | 1 | 1 | … | 0 |
| 1 | 2018 | 2 | 0 | 0 | 1 | 1 | … | 0 |
| 1 | 2016 | 2 | 1 | 0 | 1 | 1 | … | 0 |
| 1 | 2012 | 2 | 0 | 1 | 1 | 1 | … | 0 |
| 1 | 2014 | 2 | 0 | 1 | 1 | 1 | … | 0 |
| 1 | 2012 | 2 | 0 | 1 | 1 | 1 | … | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Wang, K.; Tang, A.; Tang, A.; Chen, A.; Huang, Z. Investigation of the Genes Involved in the Outbreaks of Escherichia coli and Salmonella spp. in the United States. Antibiotics 2021, 10, 1274. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101274
Li M, Wang K, Tang A, Tang A, Chen A, Huang Z. Investigation of the Genes Involved in the Outbreaks of Escherichia coli and Salmonella spp. in the United States. Antibiotics. 2021; 10(10):1274. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101274
Chicago/Turabian StyleLi, Michelle, Kyle Wang, Ashley Tang, Aaron Tang, Andrew Chen, and Zuyi Huang. 2021. "Investigation of the Genes Involved in the Outbreaks of Escherichia coli and Salmonella spp. in the United States" Antibiotics 10, no. 10: 1274. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101274