
Endolysin, a Promising Solution against Antimicrobial Resistance

 ,
,  and
and
Abstract
:1. Introduction
2. Architecture of Endolysins
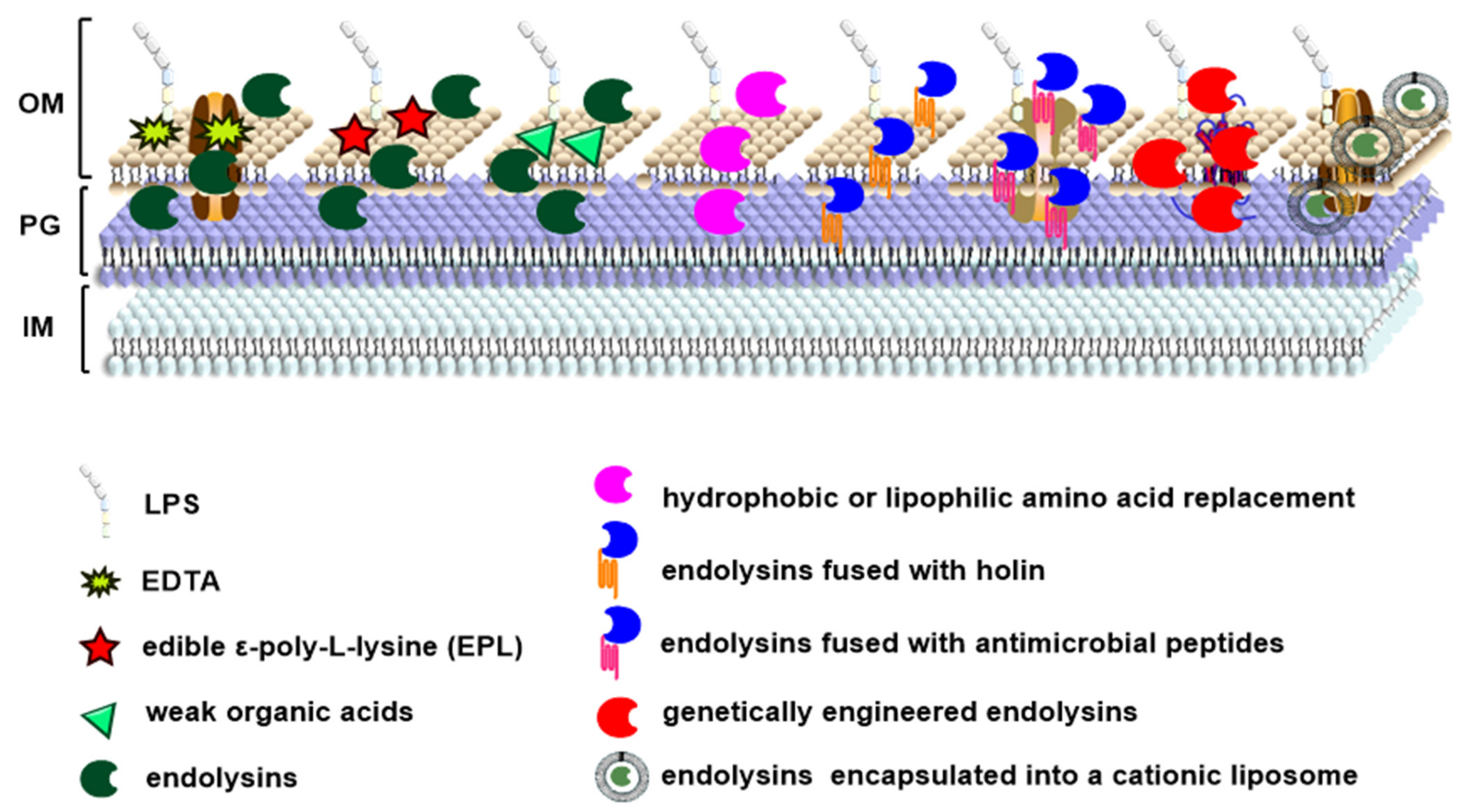
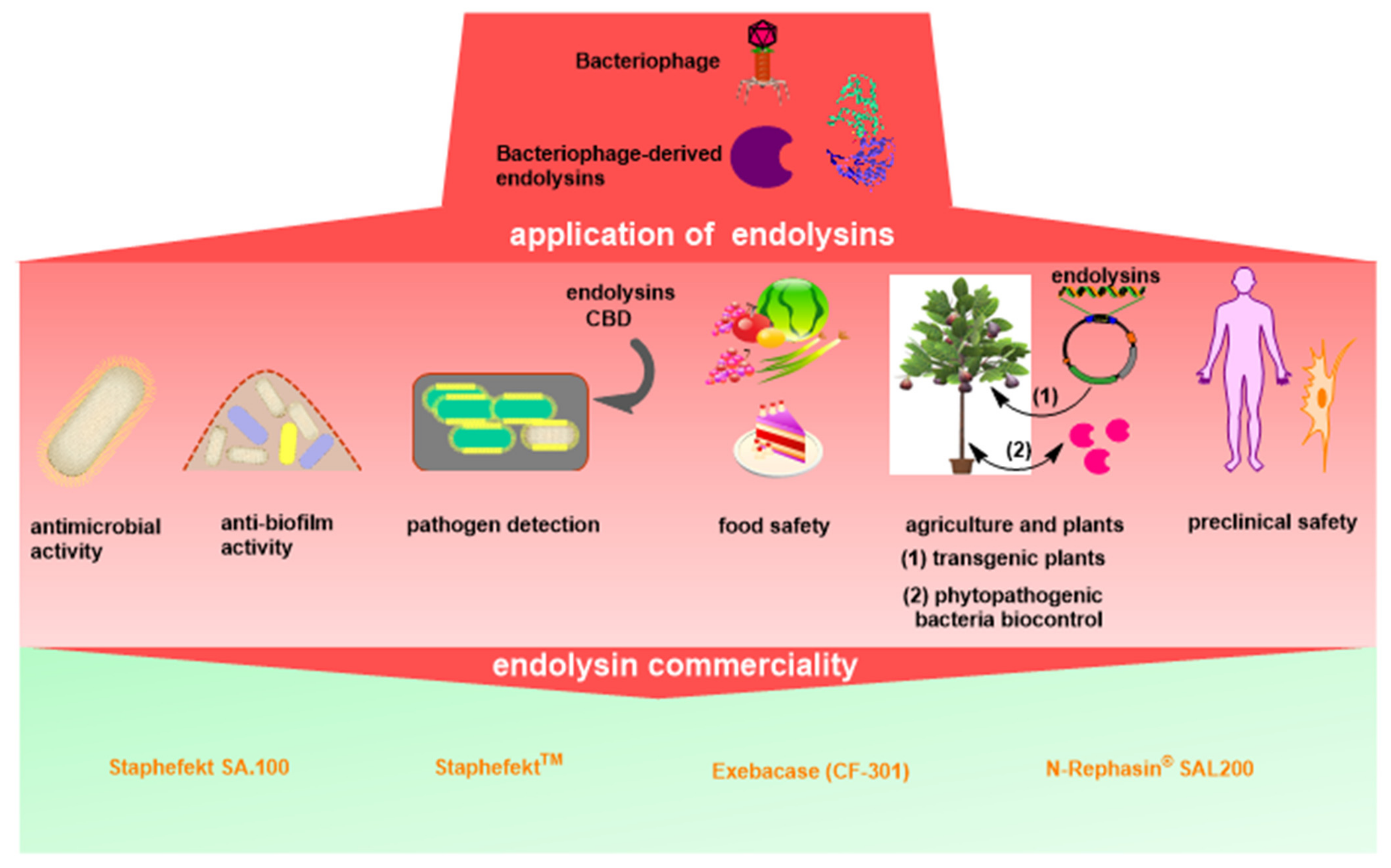
3. Antimicrobial Activity of Endolysins
4. Anti-Biofilm Activity of Endolysins
5. Endolysin Application for Pathogen Detection
6. Endolysin Application in Food Safety
7. Endolysin Application in Agriculture
8. Immunogenicity, Toxicity and Safety of Endolysins
9. Commerciality of Endolysins
10. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hadizadeh, M.; Norouzi, A.; Taghadosi, R.; Mohebi, S.; Mohammadi, M.; Hasanzade, A.; Moghadam, M.T. Prevalence of qnr, intI, and intII genes in extendedspectrum beta-lactamase (ESBL)-producing Escherichia coli isolated from clinical samples in Iran. Trop. J. Pharm. Res. 2017, 16, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-S.; Wei, W.; Hu, X.-X.; Tang, S.; Pang, J.; You, X.-F.; Fan, T.-Y.; Wang, Y.-X.; Song, D.-Q. Evolution and antibacterial evaluation of 8-hydroxy-cycloberberine derivatives as a novel family of antibacterial agents against MRSA. Molecules 2019, 24, 984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summers, W. Bacteriophage discovered. In Felix d1Herelle and the Origins of Molecular Biology; Yale University Press: New Haven, CT, USA, 1999; pp. 47–59. ISBN 9780300071276. [Google Scholar]
- Clark, J.R.; March, J.B. Bacteriophages and biotechnology: Vaccines, gene therapy and antibacterials. Trends Biotechnol. 2006, 24, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Dabrowska, K.; Switała-Jelen, K.; Opolski, A.; Weber-Dabrowska, B.; Gorski, A. Bacteriophage penetration in vertebrates. J. Appl. Microbiol. 2005, 98, 7–13. [Google Scholar] [CrossRef]
- Clark, J.R.; March, J.B. Bacterial viruses as human vaccines? Expert Rev. Vaccines 2004, 3, 463–476. [Google Scholar] [CrossRef]
- López, R.; García, E.; García, P. Enzymes for anti-infective therapy: Phage lysins. Drug Discov. Today Ther. Strateg. 2004, 1, 469–474. [Google Scholar] [CrossRef]
- Fischetti, V.A. Bacteriophage lytic enzymes: Novel anti-infectives. Trends Microbiol. 2005, 13, 491–496. [Google Scholar] [CrossRef]
- Borysowski, J.; Weber-Dąbrowska, B.; Górski, A. Bacteriophage endolysins as a novel class of antibacterial agents. Exp. Biol. Med. 2006, 231, 366–377. [Google Scholar] [CrossRef]
- Briers, Y.; Walmagh, M.; Van Puyenbroeck, V.; Cornelissen, A.; Cenens, W.; Aertsen, A.; Oliveira, H.; Azeredo, J.; Verween, G.; Pirnay, J. Engineered endolysin-based “Artilysins” to combat multidrug-resistant gram-negative pathogens. MBio 2014, 5, e01379-14. [Google Scholar] [CrossRef] [Green Version]
- Vollmer, W.; Bertsche, U. Murein (peptidoglycan) structure, architecture and biosynthesis in Escherichia coli. Biochim. Biophys. Acta Biomembr. 2008, 1778, 1714–1734. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.; Loomis, L.; Fischetti, V.A. Prevention and elimination of upper respiratory colonization of mice by group A streptococci by using a bacteriophage lytic enzyme. Proc. Natl. Acad. Sci. USA 2001, 98, 4107–4112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashani, H.H.; Schmelcher, M.; Sabzalipoor, H.; Hosseini, E.S.; Moniri, R. Recombinant endolysins as potential therapeutics against antibiotic-resistant Staphylococcus aureus: Current status of research and novel delivery strategies. Clin. Microbiol. Rev. 2018, 31, e00071-17. [Google Scholar]
- López, R.; García, E.; García, P.; García, J.L. The pneumococcal cell wall degrading enzymes: A modular design to create new lysins? Microb. Drug Resist. 1997, 3, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y.; Schmelcher, M.; Loessner, M.J.; Hendrix, J.; Engelborghs, Y.; Volckaert, G.; Lavigne, R. The high-affinity peptidoglycan binding domain of Pseudomonas phage endolysin KZ144. Biochem. Biophys. Res. Commun. 2009, 383, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Linden, S.B.; Wang, J.; Yu, J.; Nelson, D.C.; Wei, H. A chimeolysin with extended-spectrum streptococcal host range found by an induced lysis-based rapid screening method. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenech, M.; García, E.; Moscoso, M. In vitro destruction of Streptococcus pneumoniae biofilms with bacterial and phage peptidoglycan hydrolases. Antimicrob. Agents Chemother. 2011, 55, 4144–4148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuch, R.; Nelson, D.; Fischetti, V.A. A bacteriolytic agent that detects and kills Bacillus anthracis. Nature 2002, 418, 884–889. [Google Scholar] [CrossRef]
- Attai, H.; Rimbey, J.; Smith, G.P.; Brown, P.J. Expression of a peptidoglycan hydrolase from lytic bacteriophages Atu_ph02 and Atu_ph03 triggers lysis of Agrobacterium tumefaciens. Appl. Environ. Microbiol. 2017, 83, e01498-17. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, R.I.; Bustos, P.; Sepúlveda-Robles, O.; Lozano, L.; Rodríguez, C.; Fernández, J.L.; Juárez, S.; Kameyama, L.; Guarneros, G.; Dávila, G.; et al. Narrow-host-range bacteriophages that infect Rhizobium etli associate with distinct genomic types. App. Environ. Microbiol. 2014, 80, 446–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’flaherty, S.; Coffey, A.; Meaney, W.; Fitzgerald, G.; Ross, R.P. The recombinant phage lysin LysK has a broad spectrum of lytic activity against clinically relevant staphylococci, including methicillin-resistant Staphylococcus aureus. J. Bacteriol. 2005, 187, 7161–7164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmelcher, M.; Powell, A.M.; Becker, S.C.; Camp, M.J.; Donovan, D.M. Chimeric phage lysins act synergistically with lysostaphin to kill mastitis-causing Staphylococcus aureus in murine mammary glands. Appl. Environ. Microbiol. 2012, 78, 2297–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelrahman, F.; Easwaran, M.; Daramola, O.I.; Ragab, S.; Lynch, S.; Oduselu, T.J.; Khan, F.M.; Ayobami, A.; Adnan, F.; Torrents, E.; et al. Phage-Encoded Endolysins. Antibiotics 2021, 10, 124. [Google Scholar] [CrossRef] [PubMed]
- Low, L.Y.; Yang, C.; Perego, M.; Osterman, A.; Liddington, R. Role of net charge on catalytic domain and influence of cell wall binding domain on bactericidal activity, specificity, and host range of phage lysins. J. Biol. Chem. 2011, 286, 34391–34403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, H.; Melo, L.D.; Santos, S.B.; Nóbrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, H.; Azeredo, J.; Lavigne, R.; Kluskens, L.D. Bacteriophage endolysins as a response to emerging foodborne pathogens. Trends Food Sci. Technol. 2012, 28, 103–115. [Google Scholar] [CrossRef] [Green Version]
- García, P.; García, J.; García, E.; Sánchez-Puelles, J.; López, R. Modular organization of the lytic enzymes of Streptococcus pneumoniae and its bacteriophages. Gene 1990, 86, 81–88. [Google Scholar] [CrossRef]
- Kong, M.; Na, H.; Ha, N.C.; Ryu, S. LysPBC2, a Novel Endolysin Harboring a Bacillus cereus Spore Binding Domain. Appl. Environ. Microbiol. 2019, 85, e02462-18. [Google Scholar] [CrossRef] [Green Version]
- Oechslin, F.; Menzi, C.; Moreillon, P.; Resch, G. The multidomain architecture of a bacteriophage endolysin enables intramolecular synergism and regulation of bacterial lysis. J. Biol. Chem. 2021, 296, 100639. [Google Scholar] [CrossRef]
- São-José, C. Engineering of Phage-Derived Lytic Enzymes: Improving Their Potential as Antimicrobials. Antibiotics 2018, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, B.; Kong, M.; Cha, Y.; Bai, J.; Ryu, S. Simultaneous Control of Staphylococcus aureus and Bacillus cereus Using a Hybrid Endolysin LysB4EAD-LysSA11. Antibiotics 2020, 9, 906. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.C.; Swift, S.; Korobova, O.; Schischkova, N.; Kopylov, P.; Donovan, D.M.; Abaev, I. Lytic activity of the staphylolytic Twort phage endolysin CHAP domain is enhanced by the SH3b cell wall binding domain. FEMS Microbiol. Lett. 2015, 362, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciejewska, B.; Źrubek, K.; Espaillat, A.; Wiśniewska, M.; Rembacz, K.P.; Cava, F.; Dubin, G.; Drulis-Kawa, Z. Modular endolysin of Burkholderia AP3 phage has the largest lysozyme-like catalytic subunit discovered to date and no catalytic aspartate residue. Sci. Rep. 2017, 7, 14501. [Google Scholar] [CrossRef]
- Walmagh, M.; Briers, Y.; dos Santos, S.B.; Azeredo, J.; Lavigne, R. Characterization of modular bacteriophage endolysins from Myoviridae phages OBP, 201φ2-1 and PVP-SE1. PLoS ONE 2012, 7, e36991. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.; Schuch, R.; Chahales, P.; Zhu, S.; Fischetti, V.A. PlyC: A multimeric bacteriophage lysin. Proc. Natl. Acad. Sci. USA 2006, 103, 10765–10770. [Google Scholar] [CrossRef] [Green Version]
- McGowan, S.; Buckle, A.M.; Mitchell, M.S.; Hoopes, J.T.; Gallagher, D.T.; Heselpoth, R.D.; Shen, Y.; Reboul, C.F.; Law, R.H.; Fischetti, V.A. X-ray crystal structure of the streptococcal specific phage lysin PlyC. Proc. Natl. Acad. Sci. USA 2012, 109, 12752–12757. [Google Scholar] [CrossRef] [Green Version]
- Dunne, M.; Mertens, H.D.; Garefalaki, V.; Jeffries, C.M.; Thompson, A.; Lemke, E.A.; Svergun, D.I.; Mayer, M.J.; Narbad, A.; Meijers, R. The CD27L and CTP1L endolysins targeting Clostridia contain a built-in trigger and release factor. PLoS Pathog. 2014, 10, e1004228. [Google Scholar] [CrossRef] [Green Version]
- Dunne, M.; Leicht, S.; Krichel, B.; Mertens, H.D.; Thompson, A.; Krijgsveld, J.; Svergun, D.I.; Gómez-Torres, N.; Garde, S.; Uetrecht, C. Crystal structure of the CTP1L endolysin reveals how its activity is regulated by a secondary translation product. J. Biol. Chem. 2016, 291, 4882–4893. [Google Scholar] [CrossRef] [Green Version]
- Proença, D.; Velours, C.; Leandro, C.; Garcia, M.; Pimentel, M.; São-José, C. A two-component, multimeric endolysin encoded by a single gene. Mol. Microbiol. 2015, 95, 739–753. [Google Scholar] [CrossRef]
- Sanz-Gaitero, M.; Keary, R.; Garcia-Doval, C.; Coffey, A.; van Raaij, M.J. Crystallization of the CHAP domain of the endolysin from Staphylococcus aureus bacteriophage K. Acta Crystallogr. Sect. F 2013, 69, 1393–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broendum, S.S.; Buckle, A.M.; McGowan, S. Catalytic diversity and cell wall binding repeats in the phage-encoded endolysins. Mol. Microbiol. 2018, 110, 879–896. [Google Scholar] [CrossRef] [Green Version]
- Sutcliffe, I.C. A phylum level perspective on bacterial cell envelope architecture. Trends Microbiol. 2010, 18, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Navarre, W.W.; Ton-That, H.; Faull, K.F.; Schneewind, O. Multiple enzymatic activities of the murein hydrolase from staphylococcal phage φ11: Identification of a D-alanyl-glycine endopeptidase activity. J. Biol. Chem. 1999, 274, 15847–15856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loessner, M.J. Bacteriophage endolysins—Current state of research and applications. Curr. Opin. Microbiol. 2005, 8, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xu, D.; Wang, L.; Qu, M.; Li, F.; Tan, Z.; Yao, L. Characterization of a broad-spectrum endolysin LysSP1 encoded by a Salmonella bacteriophage. Appl. Microbiol. Biotechnol. 2021, 105, 5461–5470. [Google Scholar] [CrossRef]
- Ni, P.; Wang, L.; Deng, B.; Jiu, S.; Ma, C.; Zhang, C.; Almeida, A.; Wang, D.; Xu, W.; Wang, S. Characterization of a Lytic Bacteriophage against Pseudomonas syringae pv. actinidiae and Its Endolysin. Viruses 2021, 13, 631. [Google Scholar] [CrossRef]
- Han, H.; Li, X.; Zhang, T.; Wang, X.; Zou, J.; Zhang, C.; Tang, H.; Zou, Y.; Cheng, B.; Wang, R. Bioinformatic analyses of a potential Salmonella-virus-FelixO1 biocontrol phage BPS15S6 and the characterisation and anti-Enterobacteriaceae-pathogen activity of its endolysin LyS15S6. Antonie Van Leeuwenhoek 2019, 112, 1577–1592. [Google Scholar] [CrossRef]
- Oliveira, H.; Vilas Boas, D.; Mesnage, S.; Kluskens, L.D.; Lavigne, R.; Sillankorva, S.; Secundo, F.; Azeredo, J. Structural and Enzymatic Characterization of ABgp46, a Novel Phage Endolysin with Broad Anti-Gram-Negative Bacterial Activity. Front. Microbiol. 2016, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.; Yang, R.; Fan, K.; Dong, H.; Gao, C.; Wang, S.; Yu, L.; Cheng, Z.; Lei, L. External lysis of Escherichia coli by a bacteriophage endolysin modified with hydrophobic amino acids. AMB Express 2019, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Zampara, A.; Sørensen, M.C.H.; Grimon, D.; Antenucci, F.; Briers, Y.; Brøndsted, L. Innolysins: A novel approach to engineer endolysins to kill Gram-negative bacteria. BioRxiv 2018, 408948. [Google Scholar] [CrossRef]
- Xu, D.; Zhao, S.; Dou, J.; Xu, X.; Zhi, Y.; Wen, L. Engineered endolysin-based "artilysins" for controlling the gram-negative pathogen Helicobacter pylori. AMB Express 2021, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Antonova, N.P.; Vasina, D.V.; Rubalsky, E.O.; Fursov, M.V.; Savinova, A.S.; Grigoriev, I.V.; Usachev, E.V.; Shevlyagina, N.V.; Zhukhovitsky, V.G.; Balabanyan, V.U.; et al. Modulation of Endolysin LysECD7 Bactericidal Activity by Different Peptide Tag Fusion. Biomolecules 2020, 10, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, J.; Yang, E.; Chang, P.-S.; Ryu, S. Preparation and characterization of endolysin-containing liposomes and evaluation of their antimicrobial activities against gram-negative bacteria. Enzym. Microb. Technol. 2019, 128, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Hu, K.; Xie, Y.; Liu, Y.; Mu, D.; Guo, H.; Zhang, Z.; Zhang, Y.; Chang, D.; Shi, Y. A Novel Phage PD-6A3, and Its Endolysin Ply6A3, with Extended Lytic Activity Against Acinetobacter baumannii. Front. Microbiol. 2018, 9, 3302. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.M.; Gondil, V.S.; Li, C.; Jiang, M.; Li, J.; Yu, J.; Wei, H.; Yang, H. A Novel Acinetobacter baumannii Bacteriophage Endolysin LysAB54 With High Antibacterial Activity Against Multiple Gram-Negative Microbes. Front. Cell. Infect. Microbiol. 2021, 11, 70. [Google Scholar] [CrossRef]
- Plotka, M.; Kapusta, M.; Dorawa, S.; Kaczorowska, A.K.; Kaczorowski, T. Ts2631 Endolysin from the Extremophilic Thermus scotoductus Bacteriophage vB_Tsc2631 as an Antimicrobial Agent against Gram-Negative Multidrug-Resistant Bacteria. Viruses 2019, 11, 657. [Google Scholar] [CrossRef] [Green Version]
- Kaur, J.; Kour, A.; Panda, J.J.; Harjai, K.; Chhibber, S. Exploring Endolysin-Loaded Alginate-Chitosan Nanoparticles as Future Remedy for Staphylococcal Infections. AAPS PharmSciTech 2020, 21, 233. [Google Scholar] [CrossRef] [PubMed]
- Portilla, S.; Fernández, L.; Gutiérrez, D.; Rodríguez, A.; García, P. Encapsulation of the Antistaphylococcal Endolysin LysRODI in pH-Sensitive Liposomes. Antibiotics 2020, 9, 242. [Google Scholar] [CrossRef]
- Kuiper, J.W.P.; Hogervorst, J.M.A.; Herpers, B.L.; Bakker, A.D.; Klein-Nulend, J.; Nolte, P.A.; Krom, B.P. The novel endolysin XZ.700 effectively treats MRSA biofilms in two biofilm models without showing toxicity on human bone cells in vitro. Biofouling 2021, 37, 184–193. [Google Scholar] [CrossRef]
- Kim, S.; Jin, J.S.; Choi, Y.J.; Kim, J. LysSAP26, a New Recombinant Phage Endolysin with a Broad Spectrum Antibacterial Activity. Viruses 2020, 12, 1340. [Google Scholar] [CrossRef]
- Ning, H.; Lin, H.; Wang, J.; He, X.; Lv, X.; Ju, L. Characterizations of the endolysin Lys84 and its domains from phage qdsa002 with high activities against Staphylococcus aureus and its biofilms. Enzym. Microb. Technol. 2021, 148, 109809. [Google Scholar] [CrossRef]
- Yu, J.H.; Park, D.W.; Lim, J.A.; Park, J.H. Characterization of staphylococcal endolysin LysSAP33 possessing untypical domain composition. J. Microbiol. 2021, 59, 840–847. [Google Scholar] [CrossRef]
- Imanishi, I.; Uchiyama, J.; Tsukui, T.; Hisatsune, J.; Ide, K.; Matsuzaki, S.; Sugai, M.; Nishifuji, K. Therapeutic Potential of an Endolysin Derived from Kayvirus S25-3 for Staphylococcal impetigo. Viruses 2019, 11, 769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.Y.; Jun, K.I.; Kang, C.K.; Song, K.H.; Choe, P.G.; Bang, J.H.; Kim, E.S.; Park, S.W.; Kim, H.B.; Kim, N.J.; et al. Efficacy of Intranasal Administration of the Recombinant Endolysin SAL200 in a Lethal Murine Staphylococcus aureus Pneumonia Model. Antimicrob. Agents Chemother. 2019, 63, e02009-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, Y.; Son, B.; Ryu, S. Effective removal of staphylococcal biofilms on various food contact surfaces by Staphylococcus aureus phage endolysin LysCSA13. Food Microbiol. 2019, 84, 103245. [Google Scholar] [CrossRef] [PubMed]
- Son, B.; Kong, M.; Lee, Y.; Ryu, S. Development of a Novel Chimeric Endolysin, Lys109 With Enhanced Lytic Activity Against Staphylococcus aureus. Front. Microbiol. 2020, 11, 615887. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wang, Y.; Wang, J.; Zhao, Y.; Zhong, Q.; Li, G.; Fu, Z.; Lu, S. Phage Endolysin LysP108 Showed Promising Antibacterial Potential Against Methicillin-resistant Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2021, 11, 668430. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, J.; Son, B.; Ryu, S. Development of Advanced Chimeric Endolysin to Control Multidrug-Resistant Staphylococcus aureus through Domain Shuffling. ACS Infect. Dis. 2021, 7, 2081–2092. [Google Scholar] [CrossRef]
- Knaack, D.; Idelevich, E.A.; Schleimer, N.; Molinaro, S.; Kriegeskorte, A.; Peters, G.; Becker, K. Bactericidal activity of bacteriophage endolysin HY-133 against Staphylococcus aureus in comparison to other antibiotics as determined by minimum bactericidal concentrations and time-kill analysis. Diagn. Microbiol. Infect. Dis. 2019, 93, 362–368. [Google Scholar] [CrossRef]
- Chun, J.; Bai, J.; Ryu, S. Yeast Surface Display System for Facilitated Production and Application of Phage Endolysin. ACS Synth. Biol. 2020, 9, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Muharram, M.M.; Abulhamd, A.T.; Aldawsari, M.F.; Alqarni, M.H.; Labrou, N.E. Development of Staphylococcus Enzybiotics: The Ph28 Gene of Staphylococcus epidermidis Phage PH15 Is a Two-Domain Endolysin. Antibiotics 2020, 9, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.D.; Oliveira, H.; Faustino, A.; Sillankorva, S. Characterization of MSlys, the endolysin of Streptococcus pneumoniae phage MS1. Biotechnol. Rep. 2020, 28, e00547. [Google Scholar] [CrossRef]
- Kim, H.; Lee, H.G.; Kwon, I.; Seo, J. Characterization of Endolysin LyJH307 with Antimicrobial Activity Against Streptococcus bovis. Animals 2020, 10, 963. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, T.; Kwon, I.; Seo, J. Specific inhibition of Streptococcus bovis by endolysin LyJH307 supplementation shifts the rumen microbiota and metabolic pathways related to carbohydrate metabolism. J. Anim. Sci. Biotechnol. 2021, 12, 93. [Google Scholar] [CrossRef]
- Broendum, S.S.; Williams, D.E.; Hayes, B.K.; Kraus, F.; Fodor, J.; Clifton, B.E.; Geert Volbeda, A.; Codee, J.D.C.; Riley, B.T.; Drinkwater, N.; et al. High avidity drives the interaction between the streptococcal C1 phage endolysin, PlyC, with the cell surface carbohydrates of Group A Streptococcus. Mol. Microbiol. 2021, 116, 397–415. [Google Scholar] [CrossRef]
- Lu, N.; Sun, Y.; Wang, Q.; Qiu, Y.; Chen, Z.; Wen, Y.; Wang, S.; Song, Y. Cloning and characterization of endolysin and holin from Streptomyces avermitilis bacteriophage phiSASD1 as potential novel antibiotic candidates. Int. J. Biol. Macromol. 2020, 147, 980–989. [Google Scholar] [CrossRef]
- Sarjoughian, M.R.; Rahmani, F.; Abolmaali, S.; Astaneh, S.D.A. Bacillus phage endolysin, lys46, bactericidal properties against Gram-negative bacteria. Iran. J. Microbiol. 2020, 12, 607–615. [Google Scholar]
- Skorynina, A.V.; Piligrimova, E.G.; Kazantseva, O.A.; Kulyabin, V.A.; Baicher, S.D.; Ryabova, N.A.; Shadrin, A.M. Bacillus-infecting bacteriophage Izhevsk harbors thermostable endolysin with broad range specificity. PLoS ONE 2020, 15, e0242657. [Google Scholar] [CrossRef]
- Lee, K.O.; Kong, M.; Kim, I.; Bai, J.; Cha, S.; Kim, B.; Ryu, K.S.; Ryu, S.; Suh, J.Y. Structural Basis for Cell-Wall Recognition by Bacteriophage PBC5 Endolysin. Structure 2019, 27, 1355–1365.e1354. [Google Scholar] [CrossRef]
- Schuch, R.; Pelzek, A.J.; Nelson, D.C.; Fischetti, V.A. The PlyB Endolysin of Bacteriophage vB_BanS_Bcp1 Exhibits Broad-Spectrum Bactericidal Activity against Bacillus cereus Sensu Lato Isolates. Appl. Environ. Microbiol. 2019, 85, e00003-19. [Google Scholar] [CrossRef] [Green Version]
- Garde, S.; Calzada, J.; Sánchez, C.; Gaya, P.; Narbad, A.; Meijers, R.; Mayer, M.J.; Ávila, M. Effect of Lactococcus lactis expressing phage endolysin on the late blowing defect of cheese caused by Clostridium tyrobutyricum. Int. J. Food Microbiol. 2020, 329, 108686. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Kwon, J.G.; O’Sullivan, D.J.; Ryu, S.; Lee, J.H. Development of an endolysin enzyme and its cell wall-binding domain protein and their applications for biocontrol and rapid detection of Clostridium perfringens in food. Food Chem. 2021, 345, 128562. [Google Scholar] [CrossRef]
- Mondal, S.I.; Akter, A.; Draper, L.A.; Ross, R.P.; Hill, C. Characterization of an Endolysin Targeting Clostridioides difficile That Affects Spore Outgrowth. Int. J. Mol. Sci. 2021, 22, 5690. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, H.; Okada, M.; Tamai, E.; Shimamoto, T.; Shimamoto, T.; Nariya, H. A Putative Amidase Endolysin Encoded by Clostridium perfringens St13 Exhibits Specific Lytic Activity and Synergizes with the Muramidase Endolysin Psm. Antibiotics 2021, 10, 245. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zhen, X.; Zhou, H.; Zhao, F.; Fan, C.; Perčulija, V.; Tong, Y.; Mi, Z.; Ouyang, S. Structural and functional insights into a novel two-component endolysin encoded by a single gene in Enterococcus faecalis phage. PLoS Pathog. 2020, 16, e1008394. [Google Scholar] [CrossRef]
- Zhang, H.; Buttaro, B.A.; Fouts, D.E.; Sanjari, S.; Evans, B.S.; Stevens, R.H. Bacteriophage φEf11 ORF28 Endolysin, a Multifunctional Lytic Enzyme with Properties Distinct from All Other Identified Enterococcus faecalis Phage Endolysins. Appl. Environ. Microbiol. 2019, 85, e00555-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Chen, Y.; Sun, E.; Hua, L.; Peng, Z.; Wu, B. Characterization of a Lytic Bacteriophage vB_EfaS_PHB08 Harboring Endolysin Lys08 Against Enterococcus faecalis Biofilms. Microorganisms 2020, 8, 1332. [Google Scholar] [CrossRef]
- Matsui, H.; Uchiyama, J.; Ogata, M.; Nasukawa, T.; Takemura-Uchiyama, I.; Kato, S.I.; Murakami, H.; Higashide, M.; Hanaki, H. Use of Recombinant Endolysin to Improve Accuracy of Group B Streptococcus Tests. Microbiol. Spectr. 2021, 9, e0007721. [Google Scholar] [CrossRef]
- Oh, H.K.; Hwang, Y.J.; Hong, H.W.; Myung, H. Comparison of Enterococcus faecalis Biofilm Removal Efficiency among Bacteriophage PBEF129, Its Endolysin, and Cefotaxime. Viruses 2021, 13, 426. [Google Scholar] [CrossRef]
- Landlinger, C.; Tisakova, L.; Oberbauer, V.; Schwebs, T.; Muhammad, A.; Latka, A.; Van Simaey, L.; Vaneechoutte, M.; Guschin, A.; Resch, G.; et al. Engineered Phage Endolysin Eliminates Gardnerella Biofilm without Damaging Beneficial Bacteria in Bacterial Vaginosis Ex Vivo. Pathogens 2021, 10, 54. [Google Scholar] [CrossRef]
- Lu, S.Y.; Bischoff, K.M.; Rich, J.O.; Liu, S.; Skory, C.D. Recombinant bacteriophage LysKB317 endolysin mitigates Lactobacillus infection of corn mash fermentations. Biotechnol. Biofuels 2020, 13, 157. [Google Scholar] [CrossRef] [PubMed]
- Pennone, V.; Sanz-Gaitero, M.; O’Connor, P.; Coffey, A.; Jordan, K.; van Raaij, M.J.; McAuliffe, O. Inhibition of L. monocytogenes Biofilm Formation by the Amidase Domain of the Phage vB_LmoS_293 Endolysin. Viruses 2019, 11, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, G.; Jain, V. Separation of Mycobacterium smegmatis From a Mixed Culture Using the Cell Wall Binding Domain of D29 Mycobacteriophage Endolysin. Front. Microbiol. 2020, 11, 1119. [Google Scholar] [CrossRef]
- Islam, M.R.; Son, N.; Lee, J.; Lee, D.W.; Sohn, E.J.; Hwang, I. Production of bacteriophage-encoded endolysin, LysP11, in Nicotiana benthamiana and its activity as a potent antimicrobial agent against Erysipelothrix rhusiopathiae. Plant Cell Rep. 2019, 38, 1485–1499. [Google Scholar] [CrossRef]
- Santos, S.B.; Oliveira, A.; Melo, L.D.R.; Azeredo, J. Identification of the first endolysin Cell Binding Domain (CBD) targeting Paenibacillus larvae. Sci. Rep. 2019, 9, 2568. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, D.W.; Jin, J.S.; Kim, J. Antimicrobial activity of LysSS, a novel phage endolysin, against Acinetobacter baumannii and Pseudomonas aeruginosa. J. Glob. Antimicrob. Resist. 2020, 22, 32–39. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, Y.; Huang, C.; Wang, J.; Wang, X. An Endolysin LysSE24 by Bacteriophage LPSE1 Confers Specific Bactericidal Activity against Multidrug-Resistant Salmonella Strains. Microorganisms 2020, 8, 737. [Google Scholar] [CrossRef]
- Bai, J.; Lee, S.; Ryu, S. Identification and in vitro Characterization of a Novel Phage Endolysin that Targets Gram-Negative Bacteria. Microorganisms 2020, 8, 447. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Huang, H.H.; Duc, H.M.; Masuda, Y.; Honjoh, K.I.; Miyamoto, T. Endolysin LysSTG2: Characterization and application to control Salmonella Typhimurium biofilm alone and in combination with slightly acidic hypochlorous water. Food Microbiol. 2021, 98, 103791. [Google Scholar] [CrossRef]
- Park, D.W.; Park, J.H. Characterization of Endolysin LysECP26 Derived from rV5-like Phage vB_EcoM-ECP26 for Inactivation of Escherichia coli O157:H7. J. Microbiol. Biotechnol. 2020, 30, 1552–1558. [Google Scholar] [CrossRef]
- Deng, S.; Xu, Q.; Fu, Y.; Liang, L.; Wu, Y.; Peng, F.; Gao, M. Genomic Analysis of a Novel Phage Infecting the Turkey Pathogen Escherichia coli APEC O78 and Its Endolysin Activity. Viruses 2021, 13, 1034. [Google Scholar] [CrossRef] [PubMed]
- Fursov, M.V.; Abdrakhmanova, R.O.; Antonova, N.P.; Vasina, D.V.; Kolchanova, A.D.; Bashkina, O.A.; Rubalsky, O.V.; Samotrueva, M.A.; Potapov, V.D.; Makarov, V.V.; et al. Antibiofilm Activity of a Broad-Range Recombinant Endolysin LysECD7: In vitro and in vivo Study. Viruses 2020, 12, 545. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Li, X.; Wang, L.; Li, G.; Cong, C.; Li, R.; Cui, H.; Murtaza, B.; Xu, Y. The endolysin of the Acinetobacter baumannii phage vB_AbaP_D2 shows broad antibacterial activity. Microb. Biotechnol. 2021, 14, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Hammond, R.W.; Swift, S.M.; Foster-Frey, J.A.; Kovalskaya, N.Y.; Donovan, D.M. Optimized production of a biologically active Clostridium perfringens glycosyl hydrolase phage endolysin PlyCP41 in plants using virus-based systemic expression. BMC Biotechnol. 2019, 19, 101. [Google Scholar] [CrossRef]
- Basit, A.; Qadir, S.; Qureshi, S.; Rehman, S.U. Cloning and expression analysis of fused holin-endolysin from RL bacteriophage; Exhibits broad activity against multi drug resistant pathogens. Enzym. Microb. Technol. 2021, 149, 109846. [Google Scholar] [CrossRef]
- Ning, H.Q.; Lin, H.; Wang, J.X. Synergistic effects of endolysin Lysqdvp001 and ε-poly-lysine in controlling Vibrio parahaemolyticus and its biofilms. Int. J. Food Microbiol. 2021, 343, 109112. [Google Scholar] [PubMed]
- Adamczyk-Popławska, M.; Tracz-Gaszewska, Z.; Lasota, P.; Kwiatek, A.; Piekarowicz, A. Haemophilus influenzae HP1 Bacteriophage Encodes a Lytic Cassette with a Pinholin and a Signal-Arrest-Release Endolysin. Int. J. Mol. Sci. 2020, 21, 4013. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilm formation: A clinically relevant microbiological process. Clin. Infect. Dis. 2001, 33, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Sharma, U.; Vipra, A.; Channabasappa, S. Phage-derived lysins as potential agents for eradicating biofilms and persisters. Drug Discov. Today 2018, 23, 848–856. [Google Scholar] [CrossRef]
- Vukotic, G.; Obradovic, M.; Novovic, K.; Di Luca, M.; Jovcic, B.; Fira, D.; Neve, H.; Kojic, M.; McAuliffe, O. Characterization, Antibiofilm, and Depolymerizing Activity of Two Phages Active on Carbapenem-Resistant Acinetobacter baumannii. Front. Med. 2020, 7, 426. [Google Scholar] [CrossRef]
- Shkodenko, L.; Kassirov, I.; Koshel, E. Metal Oxide Nanoparticles Against Bacterial Biofilms: Perspectives and Limitations. Microorganisms 2020, 8, 1545. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, I.; Miyaji, H.; Miyata, S.; Shitomi, K.; Sugaya, T.; Ushijima, N.; Akasaka, T.; Enya, S.; Saita, S.; Kawasaki, H. Antibacterial and Antibiofilm Photodynamic Activities of Lysozyme-Au Nanoclusters/Rose Bengal Conjugates. ACS Omega 2021, 6, 9279–9290. [Google Scholar] [CrossRef] [PubMed]
- Zagami, R.; Franco, D.; Pipkin, J.D.; Antle, V.; De Plano, L.; Patanè, S.; Guglielmino, S.; Monsù Scolaro, L.; Mazzaglia, A. Sulfobutylether-β-cyclodextrin/5,10,15,20-tetrakis(1-methylpyridinium-4-yl)porphine nanoassemblies with sustained antimicrobial phototherapeutic action. Int. J. Pharm. 2020, 585, 119487. [Google Scholar] [CrossRef]
- Tonin, M.H.; Brites, F.C.; Mariano, J.R.; Freitas, K.M.S.; Ortiz, M.A.L.; Salmeron, S. Low-Level Laser and Antimicrobial Photodynamic Therapy Reduce Peri-implantitis-related Microorganisms Grown In Vitro. Eur. J. Dent. 2021. [Google Scholar] [CrossRef]
- Kortright, K.E.; Doss-Gollin, S.; Chan, B.K.; Turner, P.E. Evolution of Bacterial Cross-Resistance to Lytic Phages and Albicidin Antibiotic. Front. Microbiol. 2021, 12, 658374. [Google Scholar] [CrossRef]
- Girigoswami, K. Toxicity of Metal Oxide Nanoparticles. In Cellular and Molecular Toxicology of Nanoparticles; Saquib, Q., Faisal, M., Al-Khedhairy, A.A., Alatar, A.A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 99–122. [Google Scholar] [CrossRef]
- Dehghan Esmatabadi, M.J.; Bozorgmehr, A.; Hajjari, S.N.; Sadat Sombolestani, A.; Malekshahi, Z.V.; Sadeghizadeh, M. Review of new insights into antimicrobial agents. Cell. Mol. Biol. 2017, 63, 40–48. [Google Scholar] [CrossRef]
- Xiang, M.; Zhou, Q.; Shi, Z.; Wang, X.; Li, M.; Jia, Y.; Li, S.; Yang, F.; Wang, W.; Chen, T.; et al. A Review of Light Sources and Enhanced Targeting for Photodynamic Therapy. Curr. Med. Chem. 2021, 28, 6437–6457. [Google Scholar] [CrossRef]
- Jansen, B.; Peters, G. Foreign body associated infection. J. Antimicrob. Chemother. 1993, 32 (Suppl. A), 69–75. [Google Scholar] [CrossRef]
- Nowakowska, J.; Landmann, R.; Khanna, N. Foreign Body Infection Models to Study Host-Pathogen Response and Antimicrobial Tolerance of Bacterial Biofilm. Antibiotics 2014, 3, 378–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idelevich, E.A.; Knaack, D.; Nugroho, N.T.; Peters, G.; Bisdas, T.; Molinaro, S.; Torsello, G.B.; Becker, K.; Herten, M. Comparative in vitro activity of bacteriophage endolysin HY-133 against Staphylococcus aureus attached to vascular graft surface. Med. Microbiol. Immunol. 2020, 209, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Sass, P.; Bierbaum, G. Lytic activity of recombinant bacteriophage φ11 and φ12 endolysins on whole cells and biofilms of Staphylococcus aureus. Appl. Environ. Microbiol. 2007, 73, 347–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, J.-S.; Lee, S.-J.; Jun, S.Y.; Yoon, S.J.; Kang, S.H.; Paik, H.R.; Kang, J.O.; Choi, Y.-J. Antibacterial and biofilm removal activity of a podoviridae Staphylococcus aureus bacteriophage SAP-2 and a derived recombinant cell-wall-degrading enzyme. Appl. Microbiol. Biotechnol. 2010, 86, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, D.; Ruas-Madiedo, P.; Martínez, B.; Rodríguez, A.; García, P. Effective removal of staphylococcal biofilms by the endolysin LysH5. PLoS ONE 2014, 9, e107307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, S.Y.; Jung, G.M.; Yoon, S.J.; Oh, M.-D.; Choi, Y.-J.; Lee, W.J.; Kong, J.-C.; Seol, J.G.; Kang, S.H. Antibacterial properties of a pre-formulated recombinant phage endolysin, SAL-1. Int. J. Antimicrob. Agents 2013, 41, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Linden, S.B.; Zhang, H.; Heselpoth, R.D.; Shen, Y.; Schmelcher, M.; Eichenseher, F.; Nelson, D.C. Biochemical and biophysical characterization of PlyGRCS, a bacteriophage endolysin active against methicillin-resistant Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2015, 99, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Fenton, M.; Keary, R.; McAuliffe, O.; Ross, R.P.; O’Mahony, J.; Coffey, A. Bacteriophage-derived peptidase eliminates and prevents Staphylococcal biofilms. Int. J. Microbiol. 2013, 2013, 625341. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Shi, Y.; Ji, W.; Meng, X.; Zhang, J.; Wang, H.; Lu, C.; Sun, J.; Yan, Y. Application of a bacteriophage lysin to disrupt biofilms formed by the animal pathogen Streptococcus suis. Appl. Environ. Microbiol. 2011, 77, 8272–8279. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Yang, H.; Bi, Y.; Li, W.; Wei, H.; Li, Y. Activity of the Chimeric Lysin ClyR against Common Gram-Positive Oral Microbes and Its Anticaries Efficacy in Rat Models. Viruses 2018, 10, 380. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Ruas-Madiedo, P.; Fernández, L.; Campelo, A.B.; Briers, Y.; Nielsen, M.W.; Pedersen, K.; Lavigne, R.; García, P.; et al. Design and Selection of Engineered Lytic Proteins with Staphylococcus aureus Decolonizing Activity. Front. Microbiol. 2021, 12, 723834. [Google Scholar] [CrossRef]
- Vázquez, R.; García, P. Synergy Between Two Chimeric Lysins to Kill Streptococcus pneumoniae. Front. Microbiol. 2019, 10, 1251. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, H.; Thiagarajan, V.; Walmagh, M.; Sillankorva, S.; Lavigne, R.; Neves-Petersen, M.T.; Kluskens, L.D.; Azeredo, J. A thermostable Salmonella phage endolysin, Lys68, with broad bactericidal properties against gram-negative pathogens in presence of weak acids. PLoS ONE 2014, 9, e108376. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Feng, C.; Ren, J.; Zhuang, X.; Zhang, Y.; Zhu, Y.; Dong, K.; He, P.; Guo, X.; Qin, J. A Novel Antimicrobial Endolysin, LysPA26, against Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purevdorj-Gage, B.; Orr, M.E.; Stoodley, P.; Sheehan, K.B.; Hyman, L.E. The role of FLO11 in Saccharomyces cerevisiae biofilm development in a laboratory based flow-cell system. FEMS Yeast Res. 2007, 7, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Snel, G.; Malvisi, M.; Pilla, R.; Piccinini, R. Evaluation of biofilm formation using milk in a flow cell model and microarray characterization of Staphylococcus aureus strains from bovine mastitis. Vet. Microbiol. 2014, 174, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; Gilbert, P.; High, N.J.; Kolenbrander, P.E.; Handley, P.S. Bacterial coaggregation: An integral process in the development of multi-species biofilms. Trends Microbiol. 2003, 11, 94–100. [Google Scholar] [CrossRef]
- Chorianopoulos, N.; Tsoukleris, D.; Panagou, E.; Falaras, P.; Nychas, G.-J. Use of titanium dioxide (TiO2) photocatalysts as alternative means for Listeria monocytogenes biofilm disinfection in food processing. Food Microbiol. 2011, 28, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Kim, Y.-T.; Ryu, S.; Lee, J.-H. Biocontrol and rapid detection of food-borne pathogens using bacteriophages and endolysins. Front. Microbiol. 2016, 7, 474. [Google Scholar] [CrossRef] [PubMed]
- Schmelcher, M.; Loessner, M.J. Bacteriophage endolysins: Applications for food safety. Curr. Opin. Biotechnol. 2016, 37, 76–87. [Google Scholar] [CrossRef]
- Kretzer, J.W.; Lehmann, R.; Schmelcher, M.; Banz, M.; Kim, K.-P.; Korn, C.; Loessner, M.J. Use of high-affinity cell wall-binding domains of bacteriophage endolysins for immobilization and separation of bacterial cells. Appl. Environ. Microbiol. 2007, 73, 1992–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagens, S.; Loessner, M.J. Application of bacteriophages for detection and control of foodborne pathogens. Appl. Microbiol. Biotechnol. 2007, 76, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Idelevich, E.A.; Walther, T.; Molinaro, S.; Li, X.; Xia, G.; Wieser, A.; Peters, G.; Peschel, A.; Becker, K. Bacteriophage-based latex agglutination test for rapid identification of Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 3394–3398. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, M.G.; Carnazza, S.; De Plano, L.; Franco, D.; Nicolo, M.; Zammuto, V.; Petralia, S.; Calabrese, G.; Gugliandolo, C.; Conoci, S.; et al. Rapid detection of bacterial pathogens in blood through engineered phages-beads and integrated Real-Time PCR into MicroChip. Sens. Actuators B Chem. 2021, 329, 129227. [Google Scholar] [CrossRef]
- Ohlsson, P.; Evander, M.; Petersson, K.; Mellhammar, L.; Lehmusvuori, A.; Karhunen, U.; Soikkeli, M.; Seppä, T.; Tuunainen, E.; Spangar, A.; et al. Integrated Acoustic Separation, Enrichment, and Microchip Polymerase Chain Reaction Detection of Bacteria from Blood for Rapid Sepsis Diagnostics. Anal. Chem. 2016, 88, 9403–9411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, H.J.; Shield, C.G.; Swift, B.M. The Application of Bacteriophage Diagnostics for Bacterial Pathogens in the Agricultural Supply Chain: From Farm-to-Fork. PHAGE 2020, 1, 176–188. [Google Scholar] [CrossRef]
- Schmelcher, M.; Shabarova, T.; Eugster, M.R.; Eichenseher, F.; Tchang, V.S.; Banz, M.; Loessner, M.J. Rapid multiplex detection and differentiation of Listeria cells by use of fluorescent phage endolysin cell wall binding domains. Appl. Environ. Microbiol. 2010, 76, 5745–5756. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Wang, S.; Meng, X.; Wu, A.; Li, Q.; Song, Y.; Zhao, R.; Qiao, J. Lysin cell-binding domain-functionalized magnetic beads for detection of Staphylococcus aureus via inhibition of fluorescence of Amplex Red/hydrogen peroxide assay by intracellular catalase. Anal. Bioanal. Chem. 2019, 411, 7177–7185. [Google Scholar] [CrossRef]
- Fujinami, Y.; Hirai, Y.; Sakai, I.; Yoshino, M.; Yasuda, J. Sensitive detection of Bacillus anthracis using a binding protein originating from gamma-phage. Microbiol. Immunol. 2007, 51, 163–169. [Google Scholar] [CrossRef]
- Sainathrao, S.; Mohan, K.V.K.; Atreya, C. Gamma-phage lysin PlyG sequence-based synthetic peptides coupled with Qdot-nanocrystals are useful for developing detection methods for Bacillus anthracis by using its surrogates, B. anthracis-Sterne and B. cereus-4342. BMC Biotechnol. 2009, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tolba, M.; Ahmed, M.U.; Tlili, C.; Eichenseher, F.; Loessner, M.J.; Zourob, M. A bacteriophage endolysin-based electrochemical impedance biosensor for the rapid detection of Listeria cells. Analyst 2012, 137, 5749–5756. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, Y.; Li, H.; Yang, H.; Wei, H. Sensitive and rapid detection of Staphylococcus aureus in milk via cell binding domain of lysin. Biosens. Bioelectron. 2016, 77, 366–371. [Google Scholar] [CrossRef]
- Kong, M.; Sim, J.; Kang, T.; Nguyen, H.H.; Park, H.K.; Chung, B.H.; Ryu, S. A novel and highly specific phage endolysin cell wall binding domain for detection of Bacillus cereus. Eur. Biophys. J. 2015, 44, 437–446. [Google Scholar] [CrossRef]
- Kong, M.; Shin, J.H.; Heu, S.; Park, J.-K.; Ryu, S. Lateral flow assay-based bacterial detection using engineered cell wall binding domains of a phage endolysin. Biosens. Bioelectron. 2017, 96, 173–177. [Google Scholar] [CrossRef]
- Kwon, S.J.; Kim, D.; Lee, I.; Nam, J.; Kim, J.; Dordick, J.S. Sensitive multiplex detection of whole bacteria using self-assembled cell binding domain complexes. Anal. Chim. Acta 2018, 1030, 156–165. [Google Scholar] [CrossRef]
- Chang, Y.; Kim, M.; Ryu, S. Characterization of a novel endolysin LysSA11 and its utility as a potent biocontrol agent against Staphylococcus aureus on food and utensils. Food Microbiol. 2017, 68, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yoon, H.; Kang, D.-H.; Chang, P.-S.; Ryu, S. Endolysin LysSA97 is synergistic with carvacrol in controlling Staphylococcus aureus in foods. Int. J. Food Microbiol. 2017, 244, 19–26. [Google Scholar] [CrossRef]
- Misiou, O.; van Nassau, T.J.; Lenz, C.A.; Vogel, R.F. The preservation of Listeria-critical foods by a combination of endolysin and high hydrostatic pressure. Int. J. Food Microbiol. 2018, 266, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Sánchez, L.A.; Van Tassell, M.L.; Miller, M.J. Antimicrobial behavior of phage endolysin PlyP100 and its synergy with nisin to control Listeria monocytogenes in Queso Fresco. Food Microbiol. 2018, 72, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y. Bacteriophage-Derived Endolysins Applied as Potent Biocontrol Agents to Enhance Food Safety. Microorganisms 2020, 8, 724. [Google Scholar] [CrossRef] [PubMed]
- Yoong, P.; Schuch, R.; Nelson, D.; Fischetti, V.A. Identification of a broadly active phage lytic enzyme with lethal activity against antibiotic-resistant Enterococcus faecalis and Enterococcus faecium. J. Bacteriol. 2004, 186, 4808–4812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Bao, H.; Billington, C.; Hudson, J.A.; Wang, R. Isolation and lytic activity of the Listeria bacteriophage endolysin LysZ5 against Listeria monocytogenes in soya milk. Food Microbiol. 2012, 31, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Van Nassau, T.J.; Lenz, C.A.; Scherzinger, A.S.; Vogel, R.F. Combination of endolysins and high pressure to inactivate Listeria monocytogenes. Food Microbiol. 2017, 68, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Love, M.J.; Bhandari, D.; Dobson, R.C.; Billington, C. Potential for bacteriophage endolysins to supplement or replace antibiotics in food production and clinical care. Antibiotics 2018, 7, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Gómez-Aldapa, C.A.; Torres-Vitela, M.d.R.; Villarruel-López, A.; Castro-Rosas, J. The role of foods in Salmonella infections. In Salmonella—A Dangerous Foodborne Pathogen; Mahmoud, B., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 21–46. ISBN 978-953-307-782-6. [Google Scholar] [CrossRef]
- Rodríguez-Rubio, L.; Gerstmans, H.; Thorpe, S.; Mesnage, S.; Lavigne, R.; Briers, Y. DUF3380 domain from a Salmonella phage endolysin shows potent N-acetylmuramidase activity. Appl. Environ. Microbiol. 2016, 82, 4975–4981. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-A.; Shin, H.; Kang, D.-H.; Ryu, S. Characterization of endolysin from a Salmonella Typhimurium-infecting bacteriophage SPN1S. Res. Microbiol. 2012, 163, 233–241. [Google Scholar] [CrossRef]
- Son, B.; Yun, J.; Lim, J.-A.; Shin, H.; Heu, S.; Ryu, S. Characterization of LysB4, an endolysin from the Bacillus cereus-infecting bacteriophage B4. BMC Microbiol. 2012, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Yun, J.; Lim, J.-A.; Kang, D.-H.; Ryu, S. Characterization of an endolysin, LysBPS13, from a Bacillus cereus bacteriophage. FEMS Microbiol. Lett. 2012, 332, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Loessner, M.J.; Maier, S.K.; Daubek-Puza, H.; Wendlinger, G.; Scherer, S. Three Bacillus cereus bacteriophage endolysins are unrelated but reveal high homology to cell wall hydrolases from different bacilli. J. Bacteriol. 1997, 179, 2845–2851. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.J.; Payne, J.; Gasson, M.J.; Narbad, A. Genomic sequence and characterization of the virulent bacteriophage ΦCTP1 from Clostridium tyrobutyricum and heterologous expression of its endolysin. Appl. Environ. Microbiol. 2010, 76, 5415–5422. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.J.; Gasson, M.J.; Narbad, A. Genomic sequence of bacteriophage ATCC 8074-B1 and activity of its endolysin and engineered variants against Clostridium sporogenes. Appl. Environ. Microbiol. 2012, 78, 3685–3692. [Google Scholar] [CrossRef] [Green Version]
- Strange, R.N.; Scott, P.R. Plant disease: A threat to global food security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- McManus, P.S.; Stockwell, V.O.; Sundin, G.W.; Jones, A.L. Antibiotic use in plant agriculture. Annu. Rev. Phytopathol. 2002, 40, 443–465. [Google Scholar] [CrossRef]
- Gaeng, S.; Scherer, S.; Neve, H.; Loessner, M.J. Gene Cloning and Expression and Secretion of Listeria monocytogenes Bacteriophage-Lytic Enzymes in Lactococcus lactis. Appl. Environ. Microbiol. 2000, 66, 2951–2958. [Google Scholar] [CrossRef] [Green Version]
- Hausbeck, M.; Bell, J.; Medina-Mora, C.; Podolsky, R.; Fulbright, D. Effect of bactericides on population sizes and spread of Clavibacter michiganensis subsp. michiganensis on tomatoes in the greenhouse and on disease development and crop yield in the field. Phytopathology 2000, 90, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Düring, K.; Porsch, P.; Fladung, M.; Lörz, H. Transgenic potato plants resistant to the phytopathogenic bacterium Erwinia carotovora. Plant J. 1993, 3, 587–598. [Google Scholar] [CrossRef]
- De Vries, J.; Harms, K.; Broer, I.; Kriete, G.; Mahn, A.; Düring, K.; Wackernagel, W. The bacteriolytic activity in transgenic potatoes expressing a chimeric T4 lysozyme gene and the effect of T4 lysozyme on soil-and phytopathogenic bacteria. Syst. Appl. Microbiol. 1999, 22, 280–286. [Google Scholar] [CrossRef]
- Dow, J.M.; Ma, G.-S.; Daniels, M.J. Cloning and characterization of the rpfC gene of Xanthomonas oryzae pv. oryzae: Involvement in exopolysaccharide production and virulence to rice. Mol. Plant Microbe Interact. 1996, 9, 664–666. [Google Scholar]
- Xu, Y.; Luo, Q.-Q.; Zhou, M.-G. Identification and characterization of integron-mediated antibiotic resistance in the phytopathogen Xanthomonas oryzae pv. oryzae. PLoS ONE 2013, 8, e55962. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-N.; Lin, J.-W.; Chow, T.-Y.; Tseng, Y.-H.; Weng, S.-F. A novel lysozyme from Xanthomonas oryzae phage ϕXo411 active against Xanthomonas and Stenotrophomonas. Protein Expr. Purif. 2006, 50, 229–237. [Google Scholar] [CrossRef]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [Green Version]
- Pulawska, J. Crown gall of stone fruits and nuts, economic significance and diversity of its causal agents: Tumorigenic agrobacterium spp. J. Plant Pathol. 2010, 92, S87–S98. [Google Scholar]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Oey, M.; Lohse, M.; Scharff, L.B.; Kreikemeyer, B.; Bock, R. Plastid production of protein antibiotics against pneumonia via a new strategy for high-level expression of antimicrobial proteins. Proc. Natl. Acad. Sci. USA 2009, 106, 6579–6584. [Google Scholar] [CrossRef] [Green Version]
- Oey, M.; Lohse, M.; Kreikemeyer, B.; Bock, R. Exhaustion of the chloroplast protein synthesis capacity by massive expression of a highly stable protein antibiotic. Plant J. 2009, 57, 436–445. [Google Scholar] [CrossRef]
- Kovalskaya, N.Y.; Herndon, E.E.; Foster-Frey, J.A.; Donovan, D.M.; Hammond, R.W. Antimicrobial activity of bacteriophage derived triple fusion protein against Staphylococcus aureus. AIMS Microbiol. 2019, 5, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Kovalskaya, N.; Foster-Frey, J.; Donovan, D.M.; Bauchan, G.; Hammond, R.W. Antimicrobial Activity of Bacteriophage Endolysin Produced in Nicotiana benthamiana Plants. J. Microbiol. Biotechnol. 2016, 26, 160–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, F.; Muniesa, M. Phages in the human body. Front. Microbiol. 2017, 8, 566. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Wang, S.; Gide, M.; Zhu, D.; Lamabadu Warnakulasuriya Patabendige, H.M.; Li, C.; Cai, J.; Sun, X. A novel bacteriophage lysin-human defensin fusion protein is effective in treatment of Clostridioides difficile infection in mice. Front. Microbiol. 2019, 9, 3234. [Google Scholar] [CrossRef] [Green Version]
- Dams, D.; Briers, Y. Enzybiotics: Enzyme-based antibacterials as therapeutics. Ther. Enzym. Funct. Clin. Implic. 2019, 233–253. [Google Scholar]
- Jun, S.Y.; Jang, I.J.; Yoon, S.; Jang, K.; Yu, K.-S.; Cho, J.Y.; Seong, M.-W.; Jung, G.M.; Yoon, S.J.; Kang, S.H. Pharmacokinetics and tolerance of the phage endolysin-based candidate drug SAL200 after a single intravenous administration among healthy volunteers. Antimicrob. Agents Chemother. 2017, 61, e02629-16. [Google Scholar] [CrossRef] [Green Version]
- Rashel, M.; Uchiyama, J.; Ujihara, T.; Uehara, Y.; Kuramoto, S.; Sugihara, S.; Yagyu, K.-I.; Muraoka, A.; Sugai, M.; Hiramatsu, K. Efficient elimination of multidrug-resistant Staphylococcus aureus by cloned lysin derived from bacteriophage ϕMR11. J. Infect. Dis. 2007, 196, 1237–1247. [Google Scholar] [CrossRef] [Green Version]
- Entenza, J.; Loeffler, J.; Grandgirard, D.; Fischetti, V.; Moreillon, P. Therapeutic effects of bacteriophage Cpl-1 lysin against Streptococcus pneumoniae endocarditis in rats. Antimicrob. Agents Chemother. 2005, 49, 4789–4792. [Google Scholar] [CrossRef] [Green Version]
- Daniel, A.; Euler, C.; Collin, M.; Chahales, P.; Gorelick, K.J.; Fischetti, V.A. Synergism between a novel chimeric lysin and oxacillin protects against infection by methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2010, 54, 1603–1612. [Google Scholar] [CrossRef] [Green Version]
- Witzenrath, M.; Schmeck, B.; Doehn, J.M.; Tschernig, T.; Zahlten, J.; Loeffler, J.M.; Zemlin, M.; Müller, H.; Gutbier, B.; Schütte, H. Systemic use of the endolysin Cpl-1 rescues mice with fatal pneumococcal pneumonia. Crit. Care Med. 2009, 37, 642–649. [Google Scholar] [CrossRef]
- Herpers, B.; Badoux, P.; Pietersma, F.; Eichenseher, F.; Loessner, M. Specific lysis of methicillin susceptible and resistant Staphylococcus aureus by the endolysin Staphefekt SA. 100 TM. In Proceedings of the 24th European Congress of Clinical Microbiology and Infectious Diseases (ECCMID), Barcelona, Spain, 10–13 May 2014. [Google Scholar]
- Totté, J.E.; van Doorn, M.B.; Pasmans, S.G. Successful treatment of chronic Staphylococcus aureus-related dermatoses with the topical endolysin Staphefekt SA. 100: A report of 3 cases. Case Rep. Dermatol. 2017, 9, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Totté, J.; de Wit, J.; Pardo, L.; Schuren, F.; van Doorn, M.; Pasmans, S. Targeted anti-staphylococcal therapy with endolysins in atopic dermatitis and the effect on steroid use, disease severity and the microbiome: Study protocol for a randomized controlled trial (MAAS trial). Trials 2017, 18, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelkader, K.; Gerstmans, H.; Saafan, A.; Dishisha, T.; Briers, Y. The Preclinical and Clinical Progress of Bacteriophages and Their Lytic Enzymes: The Parts are Easier than the Whole. Viruses 2019, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Traczewski, M.M.; Ambler, J.E.; Schuch, R. Determination of MIC Quality Control Parameters for Exebacase, a Novel Lysin with Antistaphylococcal Activity. J. Clin. Microbiol. 2021, 59, e0311720. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| No. | Endolysin Name | Original Phage | Targeted Pathogens | Effective Concentration | Features of Endolysin | Reference |
|---|---|---|---|---|---|---|
| 1 | LysMR-5 | S. aureus phage | S. aureus, S. epidermidis | 500 μg/mL | encapsulation in alginate-chitosan nanoparticles | [59] |
| 2 | LysRODI | S. aureus phage | S. aureus | 20 µg/mL | encapsulation in pH-sensitive liposomes, and effective at pH 5 | [60] |
| 3 | XZ.700 | S. aureus phage | S. aureus | 250 µg/mL | chimeric endolysin and effective against S. aureus biofilms | [61] |
| 4 | LysSAP26 | S. aureus phage SAP-26 | A. baumannii, E. coli, K. pneumoniae, P. aeruginosa, S. aureus, E. faecium | 5–80 µg/mL | there was 40% protection rate in A. baumannii-infected mouse model | [62] |
| 5 | Lys84 | S. aureus phage qdsa002 | S. aureus | 10 μM | effective against biofilms | [63] |
| 6 | LysSAP33 | S. aureus phage SAP33 | S. aureus | / | higher activity against biofilms than LysK-like endolysin | [64] |
| 7 | S25-3 | S. aureus kayvirus S25-3 | S. aureus | / | genus-specific against staphylococci, particularly S. aureus | [65] |
| 8 | SAL200 | S. aureus phage | S. aureus | / | effective against severe pneumonia caused by S. aureus in a lethal murine model | [66] |
| 9 | LysCSA13 | S. aureus phage | S. aureus | 300 nM | effective against staphylococcal biofilms on various food contact surfaces | [67] |
| 10 | Lys109 | S. aureus phage | S. aureus | 100 nM | chimeric endolysin | [68] |
| 11 | LysP108 | S. aureus phage | S. aureus | / | / | [69] |
| 12 | ClyC | S. aureus phage | S. aureus | / | chimeric endolysin | [70] |
| 13 | HY-133 | S. aureus phage | S. aureus | 0.12–0.5 μg/mL | chimeric endolysin | [71] |
| 14 | LysSA11 | staphylococcal phage SA11 | S. aureus | / | expressed and surface-displayed in Saccharomyces cerevisiae | [72] |
| 15 | Ph28 | S. epidermidis phage PH15 | S. epidermidis | / | / | [73] |
| 16 | MSlys | S. pneumoniae phage MS1 | S. pneumoniae | 2 μM | / | [74] |
| 17 | LyJH307 | Streptococcus bovis phage | S. bovis, E. faecalis, S. sanguinis | 50 µg/mL | highest efficacy at pH 5.5 at 39 °C | [75] |
| 18 | LyJH307 | S. bovis | S. bovis | / | as a specific modulator for rumen | [76] |
| 19 | PlyC | streptococcal C1 phage | group A, C, and E streptococci | / | recognition of Streptococcus Group A carbohydrate backbone | [77] |
| 20 | LytSD | S. avermitilis phage phiSASD1 | S. avermitilis, B. subtilis, S. aureus, S. lutea, E. faecalis | 10 μg/mL | / | [78] |
| 21 | lys46 | B. subtilis phage | K. pneumoniae, S. Typhimurium, Proteus, E. coli | / | / | [79] |
| 22 | Ply57 | broad-host-range temperate phage, Izhevsk | B. cereus group | 1 μM | thermostability at 55 °C | [80] |
| 23 | LysPBC5 | B. cereus phage PBC5 | B. cereus | / | / | [81] |
| 24 | PlyB | B. anthracis phage vB_BanS_Bcp1 | B. cereus sensu lato group species | 16 µg/mL | potent bacteriolytic activity against all B. cereus sensu lato isolates | [82] |
| 25 | LysB4EAD-LysSA11 | B. cereus phage B4 + S. aureus phage SA11 | S. aureus, B. cereus | 3.0 µM | a hybrid endolysin | [33] |
| 26 | LysPBC2 | B. cereus phage PBC2 | Bacillus, Listeria, Clostridium | / | harboring a B. cereus spore binding domain | [30] |
| 27 | CTP1L | C. tyrobutyricum phage ΦCTP1 | C. tyrobutyricum | / | the endolysin encoding gene was introduced into the nisin producer Lactococcus lactis subsp. lactis INIA 415 | [83] |
| 28 | LysCPAS15 | C. perfringens phage CPAS-15 | C. perfringens | 45 µg/mL | C. perfringens-specific, used for pathogen detection | [84] |
| 29 | CWH | C. difficile phage phiMMP01 | C. difficile | / | cell wall binding domain prevents C. difficile spore outgrowth | [85] |
| 30 | Psa | C. perfringens phage st13 | C. perfringens | / | an amidase endolysin that specifically lyses C. perfringens | [86] |
| 31 | LysIME-EF1 | E. faecalis phage | E. faecalis | / | a novel two-component endolysin encoded by a single gene | [87] |
| 32 | ORF28 endolysin | E. faecalis phage ϕEf11 | E. faecalis | 15–31 μg/mL | multifunctional lytic enzyme, effective against E. faecalis biofilm | [88] |
| 33 | Lys08 | E. faecalis phage PHB08 | E. faecalis | 0.5–1 µg/mL | effective against E. faecalis biofilms | [89] |
| 34 | EG-LYS | E. faecalis phage | E. faecalis | 0.1 mg/mL | specific to E. faecalis | [90] |
| 35 | PBEF129 endolysin | E. faecalis phage PBEF129 | E. faecalis | 4.8 µM | effective against biofilm | [91] |
| 36 | PM-477 | Gardnerella prophage | Gardnerella | 0.13–8 µg/mL | no effect on beneficial lactobacilli or other species of vaginal bacteria | [92] |
| 37 | LysKB317 | Lactobacillus phage EcoSau | Acetobacter, Lactobacillus, Pediococcus, Streptococcus, Weissella | 0.01–1 µM | broad activity and stability from pH 4.5–7.5 up to at least 48 h; maximum activity is observed at 50 °C up to at least 72 h | [93] |
| 38 | 293 endolysin | L. monocytogenes phage vB_LmoS_293 | L. monocytogenes 473 and 3099, a serotype 4b and serogroup 1/2b-3b-7 | / | amidase | [94] |
| 39 | LysA | mycobacteriophage D29 | M. smegmatis | / | separation of M. smegmatis from a mixed culture via the cell wall binding domain | [95] |
| 40 | LysP11 | Propionibacterium phage P1.1 | E. rhusiopathiae | / | binding specifically to the E. rhusiopathiae cell wall | [96] |
| 41 | PlyPl23 | P. larvae phage phiIBB_Pl23 | P. larvae | / | first highly specific CBD targeting exclusively P. larvae cells | [97] |
| No. | Endolysin Name | Original Phage | Targeted Pathogens | Effective Concentration | Features of Endolysin | Reference |
|---|---|---|---|---|---|---|
| 1 | LysSS | S. enterica serovar Enteritidis phage SS3e | Salmonella, E. coli, P. aeruginosa, A. baumannii, K. pneumoniae, S. aureus | 0.063–0.25 mg/mL | / | [98] |
| 2 | BSP16Lys | Salmonella phage | S. Typhimurium, E. coli | / | encapsulation into a cationic liposome | [55] |
| 3 | LysSE24 | Salmonella phage LPSE1 | S. enteritidis | 0.1 μM | very stable with different pH (4.0 to 10.0) at different temperatures (20 to 60 °C) | [99] |
| 4 | M4Lys | S. enterica serovar Typhimurium phage BSPM4 | S. enterica, E. coli O157:H7, P. aeruginosa | 1 mM | the lysis function was not dependent on either holin or the Sec pathway in vitro | [100] |
| 5 | LysSP1 | S. Typhimurium phage SLMP1 | S. Typhimurium | 50 μg/mL | the optimal activity was at 40 °C and was efficiently active at alkaline condition | [47] |
| 6 | LysSTG2 | Salmonella phage STG2 | Salmonella, E. coli, P. aeruginosa | 100 μg/mL | effective on S. Typhimurium biofilm | [101] |
| 7 | LyS15S6 | Salmonella-virus-FelixO1 phage BPS15S6 | 3 species of Enterobacteriaceae, Salmonella | 2 μM | edible ε-poly-L-lysine (EPL) can be used as an outer-membrane permeabilizer | [49] |
| 8 | LysECP26 | rV5-like phage | E. coli O157:H7, Salmonella spp. | 1 µg/mL | stable at 4–55 °C | [102] |
| 9 | Lysep3 | E. coli phage | E. coli | 1750 µg/mL | activity was enhanced by modification with hydrophobic amino acids | [51] |
| 10 | LysO78 | E. coli APEC O78 phage vB_EcoM_APEC | Klebsiella, Salmonella, Shigella, Burkholderia, Yersinia, Pseudomonas, C. arctica, E. coli, R. solanacearum, A. baumannii | / | the endolysin worked with the help of 50 mM EDTA as membrane permeabilizer | [103] |
| 11 | LysECD7 | coliphage | K. pneumoniae, Pseudomonas, Acinetobacter | 3000 µg/mL | effective against forming and mature biofilm | [104] |
| 12 | LysECD7-SMAP | coliphage | K. pneumoniae, Pseudomonas, Acinetobacter | 0.5 µg/mL | the endolysin was fused to either the N- or the C-terminus of membrane-destabilizing peptides | [54] |
| 13 | Ply6A3 | A. baumannii phage PD-6A3 | A. baumannii, E. coli, S. aureus | 1 mg/mL | effective in the mouse sepsis model | [56] |
| 14 | Abtn-4 | A. baumannii phage vB_AbaP_D2 | S. aureus, P. aeruginosa, K. pneumoniae, Enterococcus, Salmonella | 5 µM | / | [105] |
| 15 | LysAB54 | A. baumannii phage p54 | A. baumannii, P. aeruginosa, K. pneumoniae, E. coli | 100 μg/mL | / | [106] |
| 16 | LysPN09 | P. syringae pv. actinidiae phage PN09 | P. syringae pv. actinidiae | 12.5–400 µg/mL | only effective against the outer-membrane-permeabilized Psa strains | [48] |
| 17 | RL_Hlys | P. aeruginosa phage RL | P. aeruginosa, K. pneumoniae, Salmonella, methicillin resistant S. aureus | / | holin was fused at the N terminus of the endolysin | [107] |
| 18 | Lysqdvp001 | V. parahaemolyticus phage | V. parahaemolyticus | ≥60 U/mL | synergistic effects with ε-poly-lysine | [108] |
| 19 | artilysin | H. pylori phage KHP30 | H. pylori | 1000 µg/mL | there was a genetic linkage between an endolysin enzyme and a holin enzyme with a section of polypeptides | [53] |
| 20 | LysHP1 | H. influenzae phage HP1 | H. influenzae, E. coli | / | endolysin expression and release was regulated by signal-arrest-release (SAR) | [109] |
| 21 | Ts2631 | T. scotoductus Bacteriophage vB_Tsc2631 | the whole Enterobacteriaceae family | 1.23 µM | extremely broad antimicrobial activity, especially with EDTA | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.u.; Wang, W.; Sun, Q.; Shah, J.A.; Li, C.; Sun, Y.; Li, Y.; Zhang, B.; Chen, W.; Wang, S. Endolysin, a Promising Solution against Antimicrobial Resistance. Antibiotics 2021, 10, 1277. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111277
Rahman Mu, Wang W, Sun Q, Shah JA, Li C, Sun Y, Li Y, Zhang B, Chen W, Wang S. Endolysin, a Promising Solution against Antimicrobial Resistance. Antibiotics. 2021; 10(11):1277. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111277
Chicago/Turabian StyleRahman, Mujeeb ur, Weixiao Wang, Qingqing Sun, Junaid Ali Shah, Chao Li, Yanmei Sun, Yuanrui Li, Bailing Zhang, Wei Chen, and Shiwei Wang. 2021. "Endolysin, a Promising Solution against Antimicrobial Resistance" Antibiotics 10, no. 11: 1277. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111277