
First Global Report of Plasmid-Mediated mcr-1 and Extended-Spectrum Beta-Lactamase-Producing Escherichia coli from Sheep in Portugal

 ,
,  ,
,
Abstract
:1. Introduction
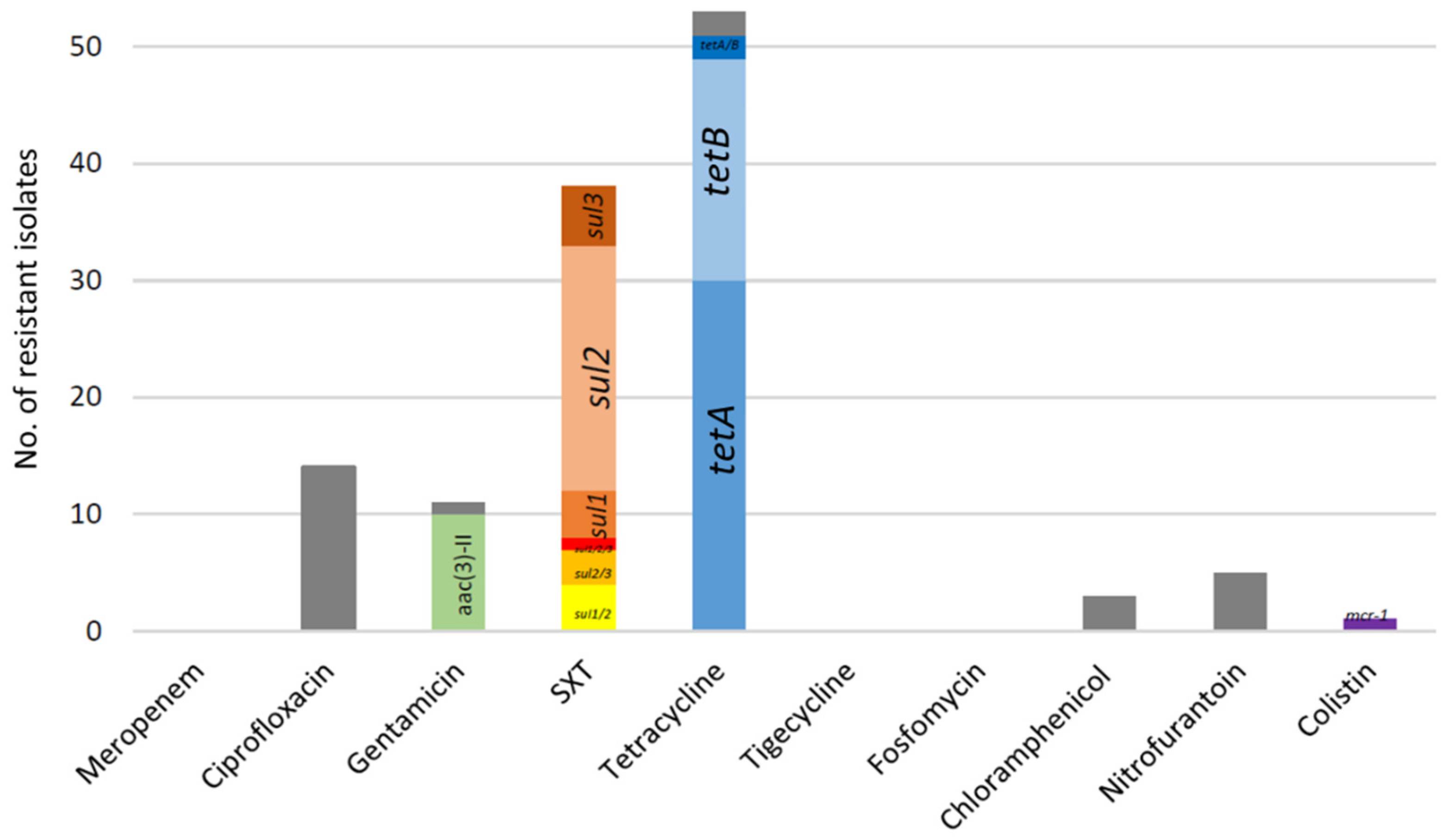
2. Results
2.1. ESBL-Producing Escherichia coli
2.1.1. Selection and Resistance Profile
2.1.2. Beta-Lactamases and Accessory Resistance Genes
2.1.3. Plasmid Content
2.1.4. Typing and Clonal Relation of ESBL-Producing E. coli
2.2. Detection of One mcr-1-Positive E. coli
3. Discussion
4. Materials and Methods
4.1. Sampling Scheme
4.2. Bacterial Isolation and Identification
4.3. Antimicrobial Susceptibility Testing and Identification
4.4. Antimicrobial Resistance Genes
4.5. E. coli Characterisation and Plasmid Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Naylor, N.R.; Atun, R.; Zhu, N.; Kulasabanathan, K.; Silva, S.; Chatterjee, A.; Knight, G.M.; Robotham, J.V. Estimating the burden of antimicrobial resistance: A systematic literature review. Antimicrob. Resist. Infect. Control. 2018, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [Green Version]
- Iramiot, J.S.; Kajumbula, H.; Bazira, J.; Kansiime, C.; Asiimwe, B.B. Antimicrobial resistance at the human–animal interface in the Pastoralist Communities of Kasese District, South Western Uganda. Sci. Rep. 2020, 10, 14737. [Google Scholar] [CrossRef] [PubMed]
- Booton, R.D.; Meeyai, A.; Alhusein, N.; Buller, H.; Feil, E.; Lambert, H.; Mongkolsuk, S.; Pitchforth, E.; Reyher, K.K.; Sakcamduang, W.; et al. One Health drivers of antibacterial resistance: Quantifying the relative impacts of human, animal and environmental use and transmission. One Health 2021, 12, 100220. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.T.; Carvalho, J.; Fernandes, J.; Palmeira, J.D.; Cunha, M.V.; Fonseca, C. Mapping the scientific knowledge of antimicrobial resistance in food-producing animals. One Health 2021, 13, 100324. [Google Scholar] [CrossRef]
- Palmeira, J.D.; Haenni, M.; Metayer, V.; Madec, J.Y.; Ferreira, H.M.N. Epidemic spread of IncI1/pST113 plasmid carrying the Extended-Spectrum Beta-Lactamase (ESBL) bla(CTX-M-8) gene in Escherichia coli of Brazilian cattle. Vet. Microbiol. 2020, 243, 108629. [Google Scholar] [CrossRef]
- Palmeira, J.D.; Ferreira, H.; Madec, J.Y.; Haenni, M. Draft genome of a ST443 mcr-1- and bla(CTX-M-2)-carrying Escherichia coli from cattle in Brazil. J. Glob. Antimicrob. Resist. 2018, 13, 269–270. [Google Scholar] [CrossRef]
- Dantas Palmeira, J.; Ferreira, H.; Madec, J.Y.; Haenni, M. Pandemic Escherichia coli ST648 isolate harbouring fosA3 and bla(CTX-M-8) on an IncI1/ST113 plasmid: A new successful combination for the spread of fosfomycin resistance? J. Glob. Antimicrob. Resist. 2018, 15, 254–255. [Google Scholar] [CrossRef]
- Torres, R.T.; Cunha, M.V.; Araujo, D.; Ferreira, H.; Fonseca, C.; Palmeira, J.D. Emergence of colistin resistance genes (mcr-1) in Escherichia coli among widely distributed wild ungulates. Environ. Pollut. 2021, 291, 118136. [Google Scholar] [CrossRef]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat. Commun. 2018, 9, 1179. [Google Scholar] [CrossRef] [Green Version]
- Canton, R.; Gonzalez-Alba, J.M.; Galán, J.C. CTX-M Enzymes: Origin and Diffusion. Front. Microbiol. 2012, 3, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th ed.; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Madec, J.-Y.; Haenni, M. Antimicrobial resistance plasmid reservoir in food and food-producing animals. Plasmid 2018, 99, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Coque, T.M.; Martínez, J.-L.; Aracil-Gisbert, S.; Lanza, V.F. Gene Transmission in the One Health Microbiosphere and the Channels of Antimicrobial Resistance. Front. Microbiol. 2019, 10, 2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolhouse, M.; Ward, M.; van Bunnik, B.; Farrar, J. Antimicrobial resistance in humans, livestock and the wider environment. Philos Trans. R. Soc. Lond B Biol. Sci. 2015, 370, 20140083. [Google Scholar] [CrossRef]
- Morris, S.T. 2—Overview of sheep production systems. In Advances in Sheep Welfare; Ferguson, D.M., Lee, C., Fisher, A., Eds.; Woodhead Publishing: Sawston, UK, 2017; pp. 19–35. [Google Scholar] [CrossRef]
- Silva, N.; Phythian, C.J.; Currie, C.; Tassi, R.; Ballingall, K.T.; Magro, G.; McNeilly, T.N.; Zadoks, R.N. Antimicrobial resistance in ovine bacteria: A sheep in wolf’s clothing? PLoS ONE 2020, 15, e0238708. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.; Menzies, P.; Reid-Smith, R.J.; Avery, B.P.; McEwen, S.A.; Moon, C.S.; Berke, O. Antimicrobial resistance in fecal generic Escherichia coli and Salmonella spp. obtained from Ontario sheep flocks and associations between antimicrobial use and resistance. Can. J. Vet. Res. 2012, 76, 109–119. [Google Scholar] [PubMed]
- Ramos, S.; Igrejas, G.; Silva, N.; Jones-Dias, D.; Capelo-Martinez, J.-L.; Caniça, M.; Poeta, P. First report of CTX-M producing Escherichia coli, including the new ST2526, isolated from beef cattle and sheep in Portugal. Food Control. 2013, 31, 208–210. [Google Scholar] [CrossRef]
- Seni, J.; Falgenhauer, L.; Simeo, N.; Mirambo, M.M.; Imirzalioglu, C.; Matee, M.; Rweyemamu, M.; Chakraborty, T.; Mshana, S.E. Multiple ESBL-Producing Escherichia coli Sequence Types Carrying Quinolone and Aminoglycoside Resistance Genes Circulating in Companion and Domestic Farm Animals in Mwanza, Tanzania, Harbor Commonly Occurring Plasmids. Front. Microbiol. 2016, 7, 142. [Google Scholar] [CrossRef]
- Geser, N.; Stephan, R.; Hächler, H. Occurrence and characteristics of extended-spectrum β-lactamase (ESBL) producing Enterobacteriaceae in food producing animals, minced meat and raw milk. Bmc Vet. Res. 2012, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Snow, L.C.; Wearing, H.; Stephenson, B.; Teale, C.J.; Coldham, N.G. Investigation of the presence of ESBL-producing Escherichia coli in the North Wales and West Midlands areas of the UK in 2007 to 2008 using scanning surveillance. Vet. Rec. 2011, 169, 656. [Google Scholar] [CrossRef]
- Benavides, J.A.; Salgado-Caxito, M.; Opazo-Capurro, A.; González Muñoz, P.; Piñeiro, A.; Otto Medina, M.; Rivas, L.; Munita, J.; Millán, J. ESBL-Producing Escherichia coli Carrying CTX-M Genes Circulating among Livestock, Dogs, and Wild Mammals in Small-Scale Farms of Central Chile. Antibiotics 2021, 10, 510. [Google Scholar] [CrossRef]
- Pehlivanoglu, F.; Turutoglu, H.; Ozturk, D.; Yardimci, H. Molecular Characterization of ESBL-Producing Escherichia coli Isolated from Healthy Cattle and Sheep. Acta Vet. 2016, 66, 520–533. [Google Scholar] [CrossRef]
- Mehmood, A.S.Q. Phenotypic and molecular characterization of esblproducing the enterobacteriaceae from animal fecal samples in southern punjab, pakistan. Sci. Int. 2021, 33, 45–48. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States; CDC: Atlanta, GA, USA, 2019.
- Dantas Palmeira, J.; Ferreira, H.M.N. Extended-spectrum beta-lactamase (ESBL)-producing Enterobacteriaceae in cattle production—A threat around the world. Heliyon 2020, 6, e03206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valsdottir, F.; Elfarsdottir Jelle, A.; Gudlaugsson, O.; Hilmarsdottir, I. Long-lasting outbreak due to CTX-M-15-producing Klebsiella pneumoniae ST336 in a rehabilitation ward: Report and literature review. J. Hosp. Infect. 2017, 97, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Pasanen, T.; Jalava, J.; Horsma, J.; Salo, E.; Pakarinen, M.; Tarkka, E.; Vaara, M.; Tissari, P. An outbreak of CTX-M-15-producing Escherichia coli, Enterobacter cloacae, and Klebsiella in a children’s hospital in Finland. Scand. J. Infect. Dis. 2014, 46, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Cottell, J.L.; Kanwar, N.; Castillo-Courtade, L.; Chalmers, G.; Scott, H.M.; Norby, B.; Loneragan, G.H.; Boerlin, P. blaCTX-M-32 on an IncN plasmid in Escherichia coli from beef cattle in the United States. Antimicrob. Agents Chemother. 2013, 57, 1096–1097. [Google Scholar] [CrossRef] [Green Version]
- Tamang, M.D.; Nam, H.-M.; Gurung, M.; Jang, G.-C.; Kim, S.-R.; Jung, S.-C.; Park, Y.H.; Lim, S.-K. Molecular Characterization of CTX-M β-Lactamase and Associated Addiction Systems in Escherichia coli Circulating among Cattle, Farm Workers, and the Farm Environment. Appl. Environ. Microbiol. 2013, 79, 3898–3905. [Google Scholar] [CrossRef] [Green Version]
- Lahlaoui, H.; De Luca, F.; Maradel, S.; Ben-Haj-Khalifa, A.; Ben Hamouda, H.; Kheder, M.; Ben Moussa, M.; Rossillini, G.-M.; Docquier, J.-D. Occurrence of conjugative IncF-type plasmids harboring the blaCTX-M-15 gene in Enterobacteriaceae isolates from newborns in Tunisia. Pediatr. Res. 2015, 77, 107–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.-H.; Hu, Z.-Q. Epidemiology and genetics of CTX-M extended-spectrum β-lactamases in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 79–101. [Google Scholar] [CrossRef]
- Novais, Â.; Cantón, R.; Moreira, R.; Peixe, L.; Baquero, F.; Coque, T.M. Emergence and Dissemination of Enterobacteriaceae Isolates Producing CTX-M-1-Like Enzymes in Spain Are Associated with IncFII (CTX-M-15) and Broad-Host-Range (CTX-M-1, -3, and -32) Plasmids. Antimicrob. Agents Chemother. 2007, 51, 796–799. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Zając, M.; Sztromwasser, P.; Bortolaia, V.; Leekitcharoenphon, P.; Cavaco, L.M.; Ziȩtek-Barszcz, A.; Hendriksen, R.S.; Wasyl, D. Occurrence and Characterization of mcr-1-Positive Escherichia coli Isolated from Food-Producing Animals in Poland, 2011–2016. Front. Microbiol. 2019, 10, 1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, S.; Urmi, U.L.; Rana, M.; Sultana, F.; Jahan, N.; Hossain, B.; Iqbal, S.; Hossain, M.M.; Mosaddek, A.S.M.; Nahar, S. High abundance of the colistin resistance gene mcr-1 in chicken gut-bacteria in Bangladesh. Sci. Rep. 2020, 10, 17292. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Fanning, S.; Gan, X.; Liu, C.; Nguyen, S.; Wang, M.; Wang, W.; Jiang, T.; Xu, J.; Li, F. Salmonella harbouring the mcr-1 gene isolated from food in China between 2012 and 2016. J. Antimicrob. Chemother. 2019, 74, 826–828. [Google Scholar] [CrossRef]
- Lv, L.; Cao, Y.; Yu, P.; Huang, R.; Wang, J.; Wen, Q.; Zhi, C.; Zhang, Q.; Liu, J.-H. Detection of mcr-1 Gene among Escherichia coli Isolates from Farmed Fish and Characterization of mcr-1-Bearing IncP Plasmids. Antimicrob. Agents Chemother. 2018, 62, e02378-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Ali, T.; Gao, J.; Ur Rahman, S.; Yu, D.; Barkema, H.W.; Huo, W.; Xu, S.; Shi, Y.; Kastelic, J.P.; et al. Co-Occurrence of Plasmid-Mediated Colistin Resistance (mcr-1) and Extended-Spectrum beta-Lactamase Encoding Genes in Escherichia coli from Bovine Mastitic Milk in China. Microb. Drug Resist. 2020, 26, 685–696. [Google Scholar] [CrossRef]
- Nagy, Á.; Székelyhidi, R.; Hanczné Lakatos, E.; Kapcsándi, V. Review on the occurrence of the mcr-1 gene causing colistin resistance in cow’s milk and dairy products. Heliyon 2021, 7, e06800. [Google Scholar] [CrossRef]
- Lei, L.; Wang, Y.; Schwarz, S.; Walsh, T.R.; Ou, Y.; Wu, Y.; Li, M.; Shen, Z. mcr-1 in Enterobacteriaceae from Companion Animals, Beijing, China, 2012–2016. Emerg. Infect. Dis. 2017, 23, 710–711. [Google Scholar] [CrossRef]
- FAO. Drivers, Dynamics and Epidemiology of Antimicrobial Resistance in Animal Production; FAO: Rome, Italy, 2016. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI: Annapolis Junction, MD, USA, 2017. [Google Scholar]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters; EUCAST: Vaxjo, Sweden, 2021. [Google Scholar]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance 2018, 23, 17-00672. [Google Scholar] [CrossRef] [PubMed]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum β-lactamases. J. Antimicrob. Chemother. 2005, 57, 154–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [Green Version]
- Saidani, M.; Messadi, L.; Chaouechi, A.; Tabib, I.; Saras, E.; Soudani, A.; Daaloul-Jedidi, M.; Mamlouk, A.; Ben Chehida, F.; Chakroun, C.; et al. High Genetic Diversity of Enterobacteriaceae Clones and Plasmids Disseminating Resistance to Extended-Spectrum Cephalosporins and Colistin in Healthy Chicken in Tunisia. Microb. Drug Resist. 2019, 25, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Cavaco, L.M.; Frimodt-Møller, N.; Hasman, H.; Guardabassi, L.; Nielsen, L.; Aarestrup, F.M. Prevalence of quinolone resistance mechanisms and associations to minimum inhibitory concentrations in quinolone-resistant Escherichia coli isolated from humans and swine in Denmark. Microb. Drug Resist. 2008, 14, 163–169. [Google Scholar] [CrossRef]
- Cattoir, V.; Weill, F.X.; Poirel, L.; Fabre, L.; Soussy, C.J.; Nordmann, P. Prevalence of qnr genes in Salmonella in France. J. Antimicrob. Chemother. 2007, 59, 751–754. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Robicsek, A.; Jacoby, G.A.; Sahm, D.; Hooper, D.C. Prevalence in the United States of aac(6’)-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrob. Agents Chemother. 2006, 50, 3953–3955. [Google Scholar] [CrossRef] [Green Version]
- Vliegenthart, J.S.; Ketelaar-van Gaalen, P.A.; van de Klundert, J.A. Nucleotide sequence of the aacC2 gene, a gentamicin resistance determinant involved in a hospital epidemic of multiply resistant members of the family Enterobacteriaceae. Antimicrob. Agents Chemother. 1989, 33, 1153–1159. [Google Scholar] [CrossRef] [Green Version]
- Bräu, B.; Pilz, U.; Piepersberg, W. Genes for gentamicin-(3)-N-acetyltransferases III and IV: I. Nucleotide sequence of the AAC(3)-IV gene and possible involvement of an IS140 element in its expression. Mol. Gen. Genet. 1984, 193, 179–187. [Google Scholar] [CrossRef]
- Cameron, F.H.; Groot Obbink, D.J.; Ackerman, V.P.; Hall, R.M. Nucleotide sequence of the AAD(2″) aminoglycoside adenylyltransferase determinant aadB. Evolutionary relationship of this region with those surrounding aadA in R538-1 and dhfrII in R388. Nucleic. Acids Res. 1986, 14, 8625–8635. [Google Scholar] [CrossRef] [PubMed]
- Melano, R.; Corso, A.; Petroni, A.; Centrón, D.; Orman, B.; Pereyra, A.; Moreno, N.; Galas, M. Multiple antibiotic-resistance mechanisms including a novel combination of extended-spectrum beta-lactamases in a Klebsiella pneumoniae clinical strain isolated in Argentina. J. Antimicrob. Chemother. 2003, 52, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarestrup, F.M.; Lertworapreecha, M.; Evans, M.C.; Bangtrakulnonth, A.; Chalermchaikit, T.; Hendriksen, R.S.; Wegener, H.C. Antimicrobial susceptibility and occurrence of resistance genes among Salmonella enterica serovar Weltevreden from different countries. J. Antimicrob. Chemother. 2003, 52, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Oka, A.; Sugisaki, H.; Takanami, M. Nucleotide sequence of the kanamycin resistance transposon Tn903. J. Mol. Biol. 1981, 147, 217–226. [Google Scholar] [CrossRef]
- Waters, S.H.; Rogowsky, P.; Grinsted, J.; Altenbuchner, J.; Schmitt, R. The tetracycline resistance determinants of RP1 and Tn1721: Nucleotide sequence analysis. Nucleic. Acids Res. 1983, 11, 6089–6105. [Google Scholar] [CrossRef]
- Sengeløv, G.; Agersø, Y.; Halling-Sørensen, B.; Baloda, S.B.; Andersen, J.S.; Jensen, L.B. Bacterial antibiotic resistance levels in Danish farmland as a result of treatment with pig manure slurry. Environ. Int. 2003, 28, 587–595. [Google Scholar] [CrossRef]
- Rahmani, M.; Peighambari, S.M.; Svendsen, C.A.; Cavaco, L.M.; Agersø, Y.; Hendriksen, R.S. Molecular clonality and antimicrobial resistance in Salmonella entericaserovars Enteritidis and Infantis from broilers in three Northern regions of Iran. BMC Vet. Res. 2013, 9, 66. [Google Scholar] [CrossRef] [Green Version]
- Perreten, V.; Boerlin, P. A new sulfonamide resistance gene (sul3) in Escherichia coli is widespread in the pig population of Switzerland. Antimicrob. Agents Chemother. 2003, 47, 1169–1172. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]


{kind=link}
| ESBL Enzyme (n = 62) | Phylogroup (n = 62) 1 | No. of Animals (n = 21) | No. of PFGE Patterns | ||||
|---|---|---|---|---|---|---|---|
| n | % | A (n = 46) | B1 (n = 15) | D (n = 1) | |||
| CTX-M-32 | 25 | 40.3 | 19 | 5 | 1 | 15 | 15 |
| CTX-M-15 | 25 | 40.3 | 23 | 2 | 0 | 16 | 11 |
| CTX-M-1 | 5 | 8.1 | 1 | 4 | 0 | 4 | 4 |
| CTX-M-14 | 1 | 1.6 | 1 | 0 | 0 | 1 | 1 |
| CTX-M-98 | 1 | 1.6 | 1 | 0 | 0 | 1 | 1 |
| CTX-M-32 + SHV-12 | 1 | 1.6 | 0 | 1 | 0 | 1 | 1 |
| SHV-12 | 4 | 6.5 | 1 | 3 | 0 | 4 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dantas Palmeira, J.; Haenni, M.; Madec, J.-Y.; Ferreira, H.M.N. First Global Report of Plasmid-Mediated mcr-1 and Extended-Spectrum Beta-Lactamase-Producing Escherichia coli from Sheep in Portugal. Antibiotics 2021, 10, 1403. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111403
Dantas Palmeira J, Haenni M, Madec J-Y, Ferreira HMN. First Global Report of Plasmid-Mediated mcr-1 and Extended-Spectrum Beta-Lactamase-Producing Escherichia coli from Sheep in Portugal. Antibiotics. 2021; 10(11):1403. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111403
Chicago/Turabian StyleDantas Palmeira, Josman, Marisa Haenni, Jean-Yves Madec, and Helena Maria Neto Ferreira. 2021. "First Global Report of Plasmid-Mediated mcr-1 and Extended-Spectrum Beta-Lactamase-Producing Escherichia coli from Sheep in Portugal" Antibiotics 10, no. 11: 1403. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111403