
In Vitro Synergism of Azithromycin Combination with Antibiotics against OXA-48-Producing Klebsiella pneumoniae Clinical Isolates

Abstract
:1. Introduction
2. Results
2.1. Antibiotic Susceptibility of OXA-48-Producing K. pneumoniae Isolates
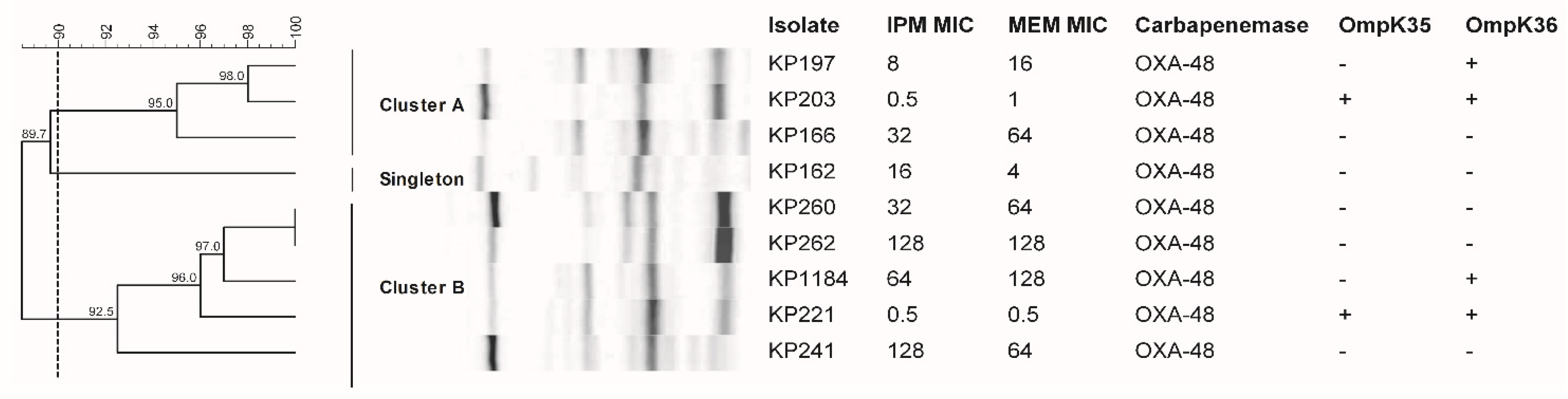
2.2. Two Major Clonalities of OXA-48-Producing K. pneumoniae
2.3. OXA-48 Expression Level among Different Carbapenem-Resistant K. pneumoniae
2.4. Impact of blaOXA-48-Carrying Plasmid in Carbapenem Susceptibility
2.5. Loss of OmpK35 and/or OmpK36 among blaOXA-48-Producing K. pneumoniae
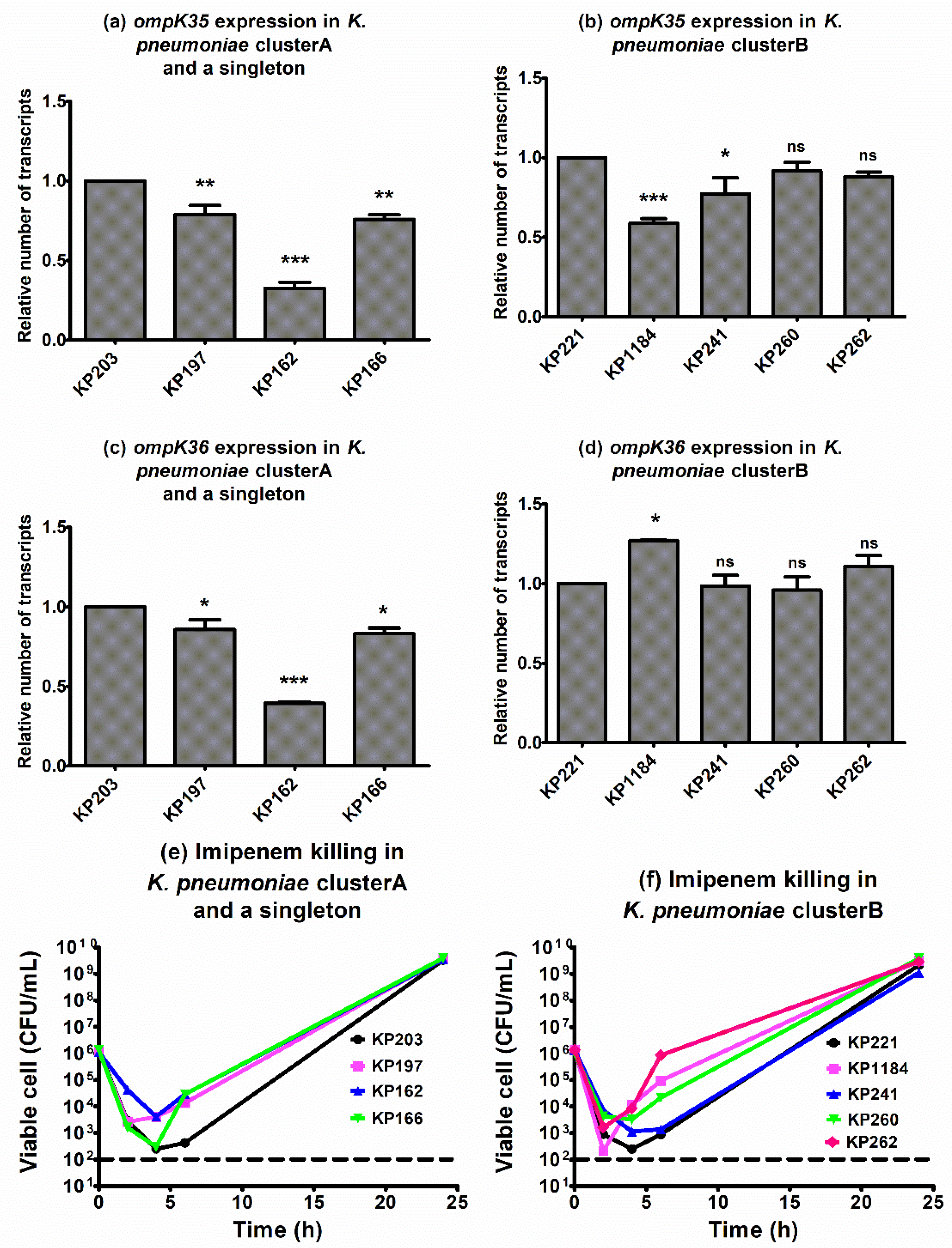
2.6. Ompk35 and Ompk36 Expression and Killing Effect of Imipenem against blaOXA-48-Carrying K. pneumoniae
2.7. Azithromycin and Fosfomycin Resistance and The Resistance Genes in blaOXA-48-Carrying K. pneumoniae
2.8. Synergistic Activity of Azithromycin with Other Antibiotics against blaOXA-48-Carrying K. pneumoniae
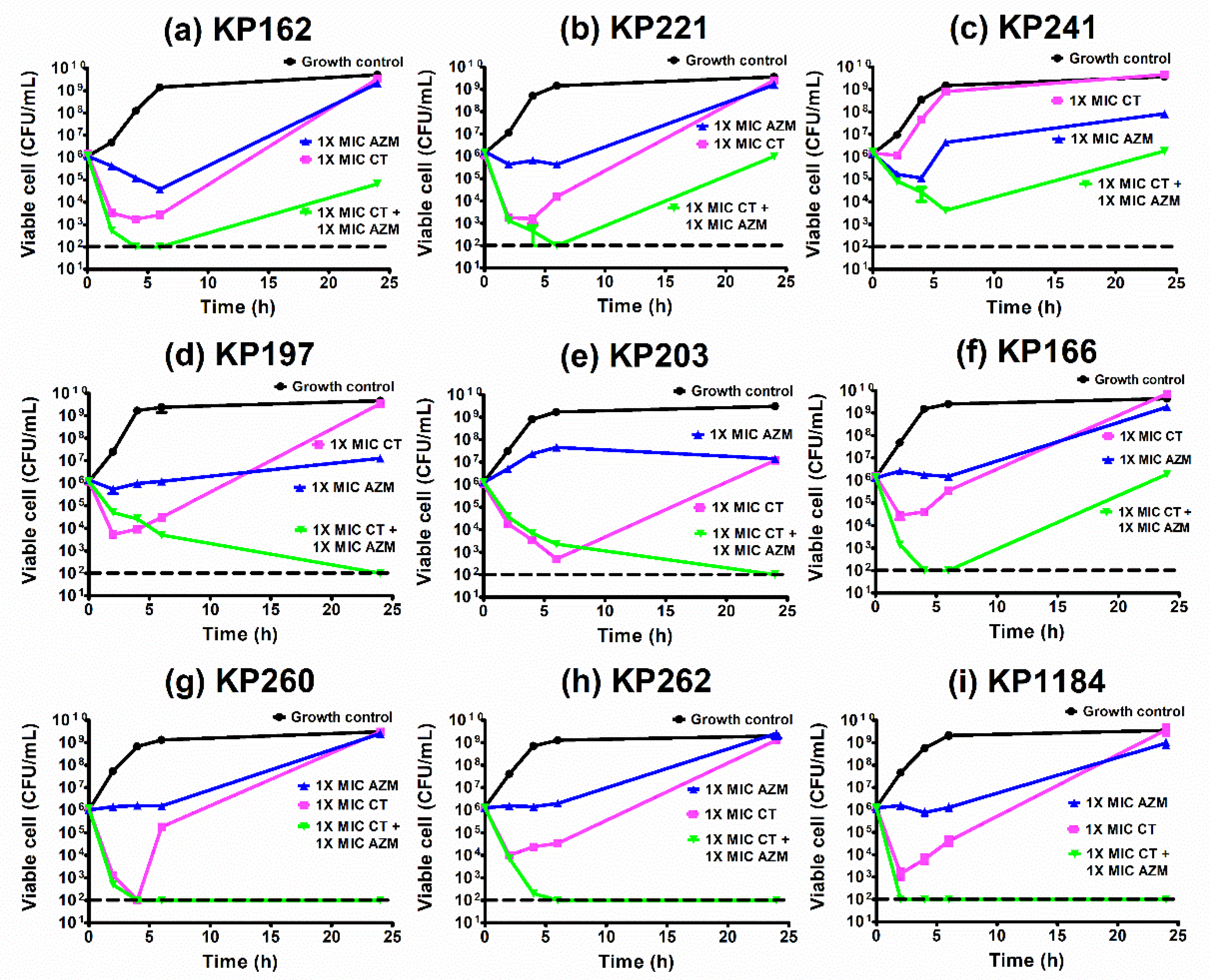
2.9. Time–Kill Curves of Azithromycin Combination with Colistin against K. pneumoniae
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Antibiotic Susceptibility Testing
4.3. Detection of Antibiotic Resistance Genes
4.4. Clonal Study
4.5. Expression Level of blaOXA-48, ompK35 and ompK36
4.6. Transformation of the blaOXA-48-Carrying Plasmids into E. coli DH5α
4.7. Outer Membrane Protein (OMP) Study
4.8. Checkerboard Assay
4.9. Time–Killing Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nordmann, P.; Naas, T.; Poirel, L. Global spread of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798. [Google Scholar] [CrossRef]
- Pitout, J.D.; Nordmann, P.; Poirel, L. Carbapenemase-Producing Klebsiella pneumoniae, a Key Pathogen Set for Global Nosocomial Dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC). Carbapenem-Resistant Enterobacteriaceae (CRE). Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/CRE-508.pdf (accessed on 1 October 2021).
- Iovleva, A.; Doi, Y. Carbapenem-Resistant Enterobacteriaceae. Clin. Lab. Med. 2017, 2, 303–315. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, K.S.; Kim, Y.B.; Jeong, B.C.; Lee, S.H. Global Dissemination of Carbapenemase-Producing Klebsiella pneumoniae: Epidemiology, Genetic Context, Treatment Options, and Detection Methods. Front. Microbiol. 2016, 7, 895. [Google Scholar] [CrossRef] [Green Version]
- Carrer, A.; Poirel, L.; Eraksoy, H.; Cagatay, A.A.; Badur, S.; Nordmann, P. Spread of OXA-48-positive carbapenem-resistant Klebsiella pneumoniae isolates in Istanbul, Turkey. Antimicrob. Agents Chemother. 2008, 52, 2950–2954. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606. [Google Scholar] [CrossRef] [Green Version]
- Evans, B.A.; Amyes, S.G. OXA beta-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [Green Version]
- Singkham-In, U.; Muhummudaree, N.; Chatsuwan, T. fosA3 overexpression with transporter mutations mediates high-level of fosfomycin resistance and silence of fosA3 in fosfomycin-susceptible Klebsiella pneumoniae producing carbapenemase clinical isolates. PLoS ONE 2020, 15, e0237474. [Google Scholar] [CrossRef]
- Tsai, Y.K.; Fung, C.P.; Lin, J.C.; Chen, J.H.; Chang, F.Y.; Chen, T.L.; Siu, L.K. Klebsiella pneumoniae outer membrane porins OmpK35 and OmpK36 play roles in both antimicrobial resistance and virulence. Antimicrob Agents Chemother. 2011, 55, 1485–1493. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A.; Mills, D.M.; Chow, N. Role of beta-lactamases and porins in resistance to ertapenem and other beta-lactams in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 3203–3206. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Martinez, L. Extended-spectrum beta-lactamases and the permeability barrier. Clin. Microbiol. Infect. 2008, 14, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jiang, X.; Wang, Y.; Li, G.; Tian, Y.; Liu, H.; Ai, F.; Ma, Y.; Wang, B.; Ruan, F.; et al. Contribution of beta-lactamases and porin proteins OmpK35 and OmpK36 to carbapenem resistance in clinical isolates of KPC-2-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2014, 58, 1214–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrill, H.J.; Pogue, J.M.; Kaye, K.S.; LaPlante, K.L. Treatment Options for Carbapenem-Resistant Enterobacteriaceae Infections. Open Forum. Infect. Dis. 2015, 2, ofv050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinos, G.P. The macrolide antibiotic renaissance. Br. J. Pharmacol. 2017, 174, 2967–2983. [Google Scholar] [CrossRef]
- Lin, L.; Nonejuie, P.; Munguia, J.; Hollands, A.; Olson, J.; Dam, Q.; Kumaraswamy, M.; Rivera, H., Jr.; Corriden, R.; Rohde, M.; et al. Azithromycin Synergizes with Cationic Antimicrobial Peptides to Exert Bactericidal and Therapeutic Activity Against Highly Multidrug-Resistant Gram-Negative Bacterial Pathogens. EBioMedicine 2015, 2, 690–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, J. Antimicrobial Resistance, from bench-to-publicside. Microbes Infect. Chemother. 2021, 1, e1182. [Google Scholar] [CrossRef]
- Zowawi, H.M.; Sartor, A.L.; Balkhy, H.H.; Walsh, T.R.; Al Johani, S.M.; AlJindan, R.Y.; Alfaresi, M.; Ibrahim, E.; Al-Jardani, A.; Al-Abri, S.; et al. Molecular characterization of carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in the countries of the Gulf cooperation council: Dominance of OXA-48 and NDM producers. Antimicrob. Agents Chemother. 2014, 58, 3085–3090. [Google Scholar] [CrossRef] [Green Version]
- Paveenkittiporn, W.; Lyman, M.; Biedron, C.; Chea, N.; Bunthi, C.; Kolwaite, A.; Janejai, N. Molecular epidemiology of carbapenem-resistant Enterobacterales in Thailand, 2016–2018. Antimicrob. Resist. Infect. Control 2021, 10, 88. [Google Scholar] [CrossRef]
- Van der Zwaluw, K.; Witteveen, S.; Wielders, L.; van Santen, M.; Landman, F.; de Haan, A.; Schouls, L.; Bosch, T.; Stuart, J.C.; Melles, D.; et al. Molecular characteristics of carbapenemase-producing Enterobacterales in the Netherlands; results of the 2014–2018 national laboratory surveillance. Clin. Microbiol. Infect. 2020, 26, 1412.e7–1412.e12. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.H.; Ma, L.; Huang, L.Y.; Yeh, K.M.; Lin, J.C.; Siu, L.K.; Chang, F.-Y. Molecular epidemiology and resistance patterns of blaOXA-48 Klebsiella pneumoniae and Escherichia coli: A nationwide multicenter study in Taiwan. J. Microbiol. Immunol. Infect. 2020, 54, 665–672. [Google Scholar] [CrossRef]
- Wiskirchen, D.E.; Nordmann, P.; Crandon, J.L.; Nicolau, D.P. Efficacy of humanized carbapenem and ceftazidime regimens against Enterobacteriaceae producing OXA-48 carbapenemase in a murine infection model. Antimicrob. Agents Chemother. 2014, 58, 1678–1683. [Google Scholar] [CrossRef] [Green Version]
- Mimoz, O.; Gregoire, N.; Poirel, L.; Marliat, M.; Couet, W.; Nordmann, P. Broad-spectrum beta-lactam antibiotics for treating experimental peritonitis in mice due to Klebsiella pneumoniae producing the carbapenemase OXA-48. Antimicrob. Agents Chemother. 2012, 56, 2759–2760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, A.; Harris, P.; Henderson, A.; Paterson, D. Treatment of Infections by OXA-48-Producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e01195–e01218. [Google Scholar] [CrossRef] [Green Version]
- Osei Sekyere, J.; Govinden, U.; Bester, L.A.; Essack, S.Y. Colistin and tigecycline resistance in carbapenemase-producing Gram-negative bacteria: Emerging resistance mechanisms and detection methods. J. Appl. Microbiol. 2016, 121, 601–617. [Google Scholar] [CrossRef]
- Rivera-Izquierdo, M.; Lainez-Ramos-Bossini, A.J.; Rivera-Izquierdo, C.; Lopez-Gomez, J.; Fernandez-Martinez, N.F.; Redruello-Guerrero, P.; Martín-delosReyes, L.M.; Martínez-Ruiz, V.; Moreno-Roldán, E.; Jiménez-Mejías, E. OXA-48 Carbapenemase-Producing Enterobacterales in Spanish Hospitals: An Updated Comprehensive Review on a Rising Antimicrobial Resistance. Antibiotics 2021, 10, 89. [Google Scholar] [CrossRef]
- Oueslati, S.; Nordmann, P.; Poirel, L. Heterogeneous hydrolytic features for OXA-48-like beta-lactamases. J. Antimicrob. Chemother. 2015, 70, 1059–1063. [Google Scholar]
- Pitout, J.D.D.; Peirano, G.; Kock, M.M.; Strydom, K.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin. Microbiol. Rev. 2019, 13, e00102-19. [Google Scholar] [CrossRef]
- Wise, M.G.; Horvath, E.; Young, K.; Sahm, D.F.; Kazmierczak, K.M. Global survey of Klebsiella pneumoniae major porins from ertapenem non-susceptible isolates lacking carbapenemases. J. Med. Microbiol. 2018, 67, 289–295. [Google Scholar] [CrossRef]
- Sugawara, E.; Kojima, S.; Nikaido, H. Klebsiella pneumoniae Major Porins OmpK35 and OmpK36 Allow More Efficient Diffusion of beta-Lactams than Their Escherichia coli Homologs OmpF and OmpC. J. Bacteriol. 2016, 198, 3200–3208. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.T.; Guo, M.K.; Ke, S.C.; Lin, Y.P.; Pang, Y.C.; Nguyen, H.V.; Chen, C.-M. Characterization of the Genetic Background of KPC-2-Producing Klebsiella pneumoniae with Insertion Elements Disrupting the ompK36 Porin Gene. Microb. Drug Resist. 2020, 26, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.L.C.; Romano, M.; Kerry, L.E.; Kwong, H.S.; Low, W.W.; Brett, S.J.; Clements, A.; Beis, K.; Frankel, G. OmpK36-mediated Carbapenem resistance attenuates ST258 Klebsiella pneumoniae in vivo. Nat. Commun. 2019, 10, 3957. [Google Scholar] [CrossRef] [Green Version]
- Leclercq, R. Mechanisms of resistance to macrolides and lincosamides: Nature of the resistance elements and their clinical implications. Clin. Infect. Dis. 2002, 34, 482–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, C.; Ruiz-Roldan, L.; Mateu, J.; Ochoa, T.J.; Ruiz, J. Azithromycin resistance levels and mechanisms in Escherichia coli. Sci. Rep. 2019, 9, 6089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Lin, X.; Yao, X.; Huang, Y.; Liu, W.; Ma, T.; Fang, B. Synergistic Antimicrobial Activity of Colistin in Combination with Rifampin and Azithromycin against Escherichia coli Producing MCR-1. Antimicrob. Agents Chemother. 2018, 62, e01631-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magreault, S.; Roy, C.; Launay, M.; Sermet-Gaudelus, I.; Jullien, V. Pharmacokinetic and Pharmacodynamic Optimization of Antibiotic Therapy in Cystic Fibrosis Patients: Current Evidences, Gaps in Knowledge and Future Directions. Clin. Pharmacokinet. 2021, 60, 409–445. [Google Scholar] [CrossRef]
- Reijnders, T.D.Y.; Saris, A.; Schultz, M.J.; van der Poll, T. Immunomodulation by macrolides: Therapeutic potential for critical care. Lancet Respir. Med. 2020, 8, 619–630. [Google Scholar] [CrossRef]
- Zhang, F.; Li, Q.; Bai, J.; Ding, M.; Yan, X.; Wang, G.; Zhu, B.; Zhou, Y. Heteroresistance to Amikacin in Carbapenem-Resistant Klebsiella pneumoniae Strains. Front. Microbiol. 2021, 12, 682239. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_10.0_Breakpoint_Tables.pdf (accessed on 13 October 2021).
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Ellington, M.J.; Kistler, J.; Livermore, D.M.; Woodford, N. Multiplex PCR for rapid detection of genes encoding acquired metallo-beta-lactamases. J. Antimicrob. Chemother. 2007, 59, 321–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutcliffe, J.; Grebe, T.; Tait-Kamradt, A.; Wondrack, L. Detection of erythromycin-resistant determinants by PCR. Antimicrob. Agents Chemother. 1996, 40, 2562–2566. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.C.; Chung, W.O.; Roe, D.; Xia, M.; Marquez, C.; Borthagaray, G.; Whittington, W.L.; Holmes, K.K. Erythromycin-resistant Neisseria gonorrhoeae and oral commensal Neisseria spp. carry known rRNA methylase genes. Antimicrob. Agents Chemother. 1999, 43, 1367–1372. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Nawaz, M.S.; Khan, A.A.; Cerniglia, C.E. Simultaneous detection of erythromycin-resistant methylase genes ermA and ermC from Staphylococcus spp. by multiplex-PCR. Mol. Cell Probes 1999, 13, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Ragheb, S.M.; Tawfick, M.M.; El-Kholy, A.A.; Abdulall, A.K. Phenotypic and Genotypic Features of Klebsiella pneumoniae Harboring Carbapenemases in Egypt: OXA-48-Like Carbapenemases as an Investigated Model. Antibiotics 2020, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- Singkham-In, U.; Chatsuwan, T. In vitro activities of carbapenems in combination with amikacin, colistin, or fosfomycin against carbapenem-resistant Acinetobacter baumannii clinical isolates. Diagn. Microbiol. Infect. Dis. 2018, 91, 169–174. [Google Scholar] [CrossRef]
- Lu, P.L.; Hsieh, Y.J.; Lin, J.E.; Huang, J.W.; Yang, T.Y.; Lin, L.; Tseng, S.P. Characterisation of fosfomycin resistance mechanisms and molecular epidemiology in extended-spectrum beta-lactamase-producing Klebsiella pneumoniae isolates. Int. J. Antimicrob. Agents 2016, 48, 564–568. [Google Scholar] [CrossRef]
- Hamzaoui, Z.; Ocampo-Sosa, A.; Martinez, M.F.; Landolsi, S.; Ferjani, S.; Maamar, E.; Saidani, M.; Slim, A.; Martinez-Martinez, L.; Boutiba-Ben Boubaker, I. Role of association of OmpK35 and OmpK36 alteration and blaESBL and/or blaampC in conferring carbapenem resistance among non-producing carbapenemase-Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2018, 52, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.F.; Chen, X.Y.; Yan, G.F.; Wang, Y.P.; Ying, C.M. Study of the correlation of imipenem resistance with efflux pumps AdeABC, AdeIJK, AdeDE and AbeM in clinical isolates of Acinetobacter baumannii. Chemotherapy 2012, 58, 152–158. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | MIC (Mg/L) (Interpretation) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| IMP | MEM | ETP | AMK | CIP | CRO | CT | FOF | AZM | |
| KP162 | 16 (R) | 4 (I) | 64 (R) | 4 (S) | 128 (R) | 128 (R) | 0.5 (S) | 128 (R) | 64 (R) |
| KP166 | 32 (R) | 64 (R) | 256 (R) | 4 (S) | 1 (R) | >256 (R) | 0.25 (S) | 256 (R) | 32 (R) |
| KP197 | 8 (R) | 16 (R) | 128 (R) | 16 (R) | 64 (R) | >256 (R) | 0.5 (S) | 32 (S) | 32 (R) |
| KP203 | 0.5 (S) | 1 (S) | 2 (R) | 4 (S) | 128 (R) | 128 (R) | 0.5 (S) | 16 (S) | 8 (S) |
| KP221 | 0.5 (S) | 0.5 (S) | 2 (R) | 1 (S) | 128 (R) | 128 (R) | 0.5 (S) | 8 (S) | 128 (R) |
| KP241 | 128 (R) | 64 (R) | 256 (R) | 2 (S) | 64 (R) | >256 (R) | 0.25 (S) | 128 (R) | 128 (R) |
| KP260 | 32 (R) | 64 (R) | 128 (R) | 4 (S) | >256 (R) | >256 (R) | 0.5 (S) | 256 (R) | 1024 (R) |
| KP262 | 128 (R) | 128 (R) | >256 (R) | 8 (S) | >256 (R) | >256 (R) | 0.25 (S) | 1024 (R) | 1024 (R) |
| KP1184 | 64 (R) | 128 (R) | >256 (R) | 64 (R) | 64 (R) | >256 (R) | 1 (S) | 16 (S) | 16 (S) |
| Isolate | MIC (mg/L) | ||||||
|---|---|---|---|---|---|---|---|
| Carbapenemase Gene | IPM | MEM | ETP | CRO | FOF | AMK | |
| E. coli DH5α | None | 0.25 | 0.015 | 0.015 | 0.015 | 0.5 | 0.5 |
| KP162 | blaOXA-48 | 16 | 4 | 64 | 128 | 128 | 4 |
| KP162_T | blaOXA-48 | 128 | 64 | >256 | >256 | >256 | 8 |
| KP197 | blaOXA-48 | 8 | 16 | 128 | >256 | 32 | 16 |
| KP197_T | blaOXA-48 | 8 | 16 | 16 | 32 | 2 | 1 |
| KP203 | blaOXA-48 | 0.5 | 1 | 2 | 128 | 16 | 4 |
| KP203_T | blaOXA-48 | 64 | 32 | 256 | >256 | >256 | 2 |
| KP166 | blaOXA-48 | 32 | 64 | 256 | >256 | 256 | 4 |
| KP166_T | blaOXA-48 | 64 | 32 | 256 | >256 | 256 | 2 |
| KP260 | blaOXA-48 | 32 | 64 | 12 | >256 | 256 | 4 |
| KP260_T | blaOXA-48 | 16 | 16 | 16 | 16 | 4 | 1 |
| KP262 | blaOXA-48 | 128 | 128 | >256 | >256 | 256 | 8 |
| KP262_T | blaOXA-48 | 64 | 32 | 256 | >256 | 128 | 2 |
| KP1184 | blaOXA-48 | 64 | 128 | >256 | >256 | >256 | 64 |
| KP1184_T | blaOXA-48 | 8 | 16 | 16 | 32 | 4 | 1 |
| KP221 | blaOXA-48 | 0.5 | 0.5 | 2 | 128 | 8 | 1 |
| KP221_T | blaOXA-48 | 16 | 16 | 16 | 16 | 4 | 1 |
| KP241 | blaOXA-48 | 128 | 64 | 256 | >256 | 128 | 2 |
| KP241_T | blaOXA-48 | 8 | 16 | 16 | 16 | 4 | 1 |
| Isolate | OMP | MIC (Mg/L) (Interpretation) | Antibiotic Resistance Gene | FICI (Interpretation) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ompk35 | Ompk36 | IPM | CT | FOF | AZM | fosa3 | fosa5 | ermB | ermC | AZM + IPM | AZM + CT | AZM + FOF | |
| KP162 | − | − | 16 (R) | 0.5 (S) | 128 (R) | 64 (R) | − | + | − | + | 0.75 (N) | 0.5 (Syn) | 0.38 (Syn) |
| KP197 | − | + | 8 (R) | 0.5 (S) | 32 (S) | 32 (R) | + | + | − | + | 2 (N) | 0.5 (Syn) | 0.5 (Syn) |
| KP203 | + | + | 0.5 (S) | 0.5 (S) | 16 (S) | 8 (S) | − | + | − | + | 2 (N) | 0.5 (Syn) | 0.38 (Syn) |
| KP166 | − | − | 32 (R) | 0.25 (S) | 256 (R) | 32 (R) | + | + | − | + | 0.75 (N) | 0.5 (Syn) | 0.38 (Syn) |
| KP260 | − | − | 32 (R) | 0.5 (S) | 256 (R) | 1024 (R) | − | + | + | + | 2 (N) | 0.38 (Syn) | 0.75 (N) |
| KP262 | − | − | 128 (R) | 0.25 (S) | 256 (R) | 1024 (R) | + | + | + | + | 0.75 (N) | 0.75 (N) | 0.75 (N) |
| KP1184 | − | + | 64 (R) | 1 (S) | 512 (R) | 16 (S) | + | + | − | + | 0.5 (Syn) | 0.38 (Syn) | 0.38 (Syn) |
| KP221 | + | + | 0.5 (S) | 0.5 (S) | 8 (S) | 128 (R) | − | + | − | + | 0.75 (N) | 0.38 (Syn) | 0.38 (Syn) |
| KP241 | − | − | 128 (R) | 0.25 (S) | 128 (R) | 128 (R) | + | + | − | + | 0.75 (N) | 0.5 (Syn) | 0.38 (Syn) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singkham-in, U.; Muhummudaree, N.; Chatsuwan, T. In Vitro Synergism of Azithromycin Combination with Antibiotics against OXA-48-Producing Klebsiella pneumoniae Clinical Isolates. Antibiotics 2021, 10, 1551. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121551
Singkham-in U, Muhummudaree N, Chatsuwan T. In Vitro Synergism of Azithromycin Combination with Antibiotics against OXA-48-Producing Klebsiella pneumoniae Clinical Isolates. Antibiotics. 2021; 10(12):1551. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121551
Chicago/Turabian StyleSingkham-in, Uthaibhorn, Netchanok Muhummudaree, and Tanittha Chatsuwan. 2021. "In Vitro Synergism of Azithromycin Combination with Antibiotics against OXA-48-Producing Klebsiella pneumoniae Clinical Isolates" Antibiotics 10, no. 12: 1551. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121551