
The Role of Staphylococcus aureus YycFG in Gene Regulation, Biofilm Organization and Drug Resistance


{kind=link}
Abstract
:1. Introduction
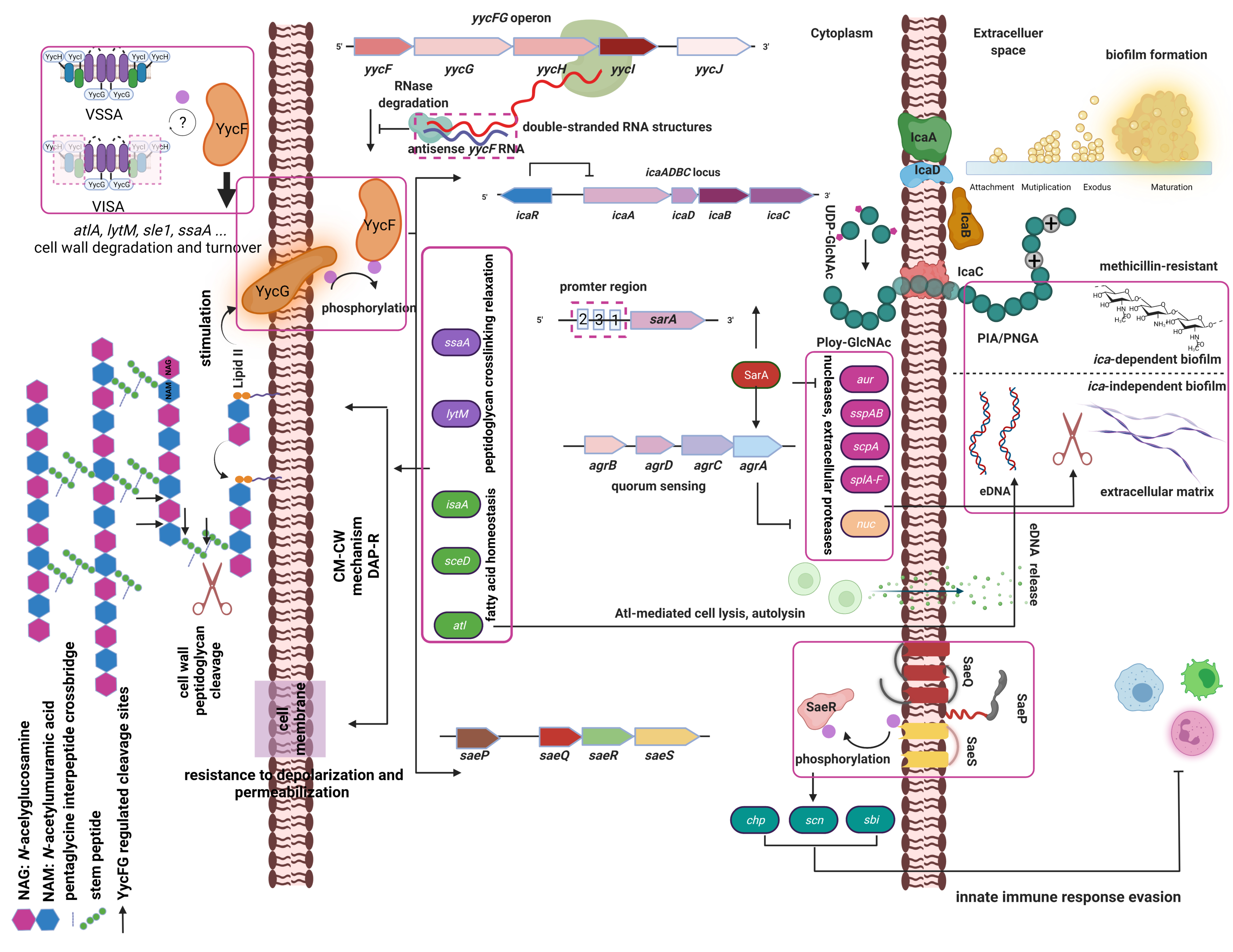
2. Regulation of S. aureus YycFG Two-Component Systems
2.1. S. aureus Two-Component Systems
2.2. Regulatory Roles of the YycFG TCS in Cell Wall Biosynthesis and Biofilm Formation
2.3. Regulatory Roles of the YycFG TCS in Response to Host Immunity
3. The Impact of YycFG Two-Component Systems on Antibiotic Resistance
3.1. S. aureus Biofilms and Antimicrobial Drug Resistance
3.2. YycFG TCS-Associated Cell Membrane and Cell Wall Biogenesis Involvement in Drug Resistance
4. Targeting the S. aureus Two-Component Systems
4.1. Molecular Targets
4.2. Non-Coding RNA Regulation
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Bhattacharya, M.; Wozniak, D.J.; Stoodley, P.; Hall-Stoodley, L. Prevention and treatment of Staphylococcus aureus biofilms. Expert Rev. Anti. Infect. Ther. 2015, 13, 1499–1516. [Google Scholar] [CrossRef] [Green Version]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiti, P.; Chatterjee, S.; Dey, R.; Kundu, A. Biofilms on indwelling urologic devices: Microbes and antimicrobial management prospect. Ann. Med. Heal. Sci. Res. 2014, 4, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Francischetto, O.; Da Silva, L.A.P.; E Senna, K.M.S.; Vasques, M.R.; Barbosa, G.F.; Weksler, C.; Ramos, R.G.; Golebiovski, W.F.; Lamas, C. Healthcare-Associated Infective Endocarditis: Case Series in Referral Hospital from 2006 to 2011. Arq. Bras. Cardiol. 2014, 103, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.-O.; Yoo, J.I.; Yoo, J.S.; Chung, H.-S.; Park, S.-H.; Kim, H.S.; Lee, Y.S.; Chung, G.T. Investigation of Biofilm Formation and its Association with the Molecular and Clinical Characteristics of Methicillin-resistant Staphylococcus aureus. Osong Public Heal Res. Perspect. 2013, 4, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, V.J.; Chopra, I.; O’Neill, A.J. Staphylococcus aureus Biofilms Promote Horizontal Transfer of Antibiotic Resistance. Antimicrob. Agents Chemother. 2013, 57, 1968–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattos-Graner, R.O.; Duncan, M.J. Two-component signal transduction systems in oral bacteria. J. Oral Microbiol. 2017, 9, 1400858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Wu, Y.; Lin, Z.; Bertram, R.; Götz, F.; Zhang, Y.; Qu, D. Identification of Genes Controlled by the Essential YycFG Two-Component System Reveals a Role for Biofilm Modulation in Staphylococcus epidermidis. Front. Microbiol. 2017, 8, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabret, C.; Hoch, J.A. A two-component signal transduction system essential for growth of Bacillus subtilis: Implications for anti-infective therapy. J. Bacteriol. 1998, 180, 6375–6383. [Google Scholar] [CrossRef]
- Martin, P.K.; Li, T.; Sun, D.; Biek, D.P.; Schmid, M.B. Role in Cell Permeability of an Essential Two-Component System in Staphylococcus aureus. J. Bacteriol. 1999, 181, 3666–3673. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, M.; García, B.; Valle, J.; Rapún, B.; Mozos, I.R.D.L.; Solano, C.; Martí, M.; Penadés, J.R.; Toledo-Arana, A.; Lasa, I. Sensory deprivation in Staphylococcus aureus. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dubrac, S.; Msadek, T. Identification of Genes Controlled by the Essential YycG/YycF Two-Component System of Staphylococcus aureus. J. Bacteriol. 2004, 186, 1175–1181. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Gerke, C.; Kraft, A.; Süssmuth, R.; Schweitzer, O.; Götz, F. Characterization of theN-Acetylglucosaminyltransferase Activity Involved in the Biosynthesis of the Staphylococcus epidermidis polysaccharide Intercellular Adhesin. J. Biol. Chem. 1998, 273, 18586–18593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilmann, C.; Schweitzer, O.; Gerke, C.; Vanittanakom, N.; Mack, D.; Götz, F. Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis. Mol. Microbiol. 1996, 20, 1083–1091. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Nguyen, T.H.; Otto, M. The staphylococcal exopolysaccharide PIA—Biosynthesis and role in biofilm formation, colonization, and infection. Comput. Struct Biotechnol. J. 2020, 18, 3324–3334. [Google Scholar] [CrossRef]
- Wu, S.; Huang, F.; Zhang, H.; Lei, L. Staphylococcus aureus biofilm organization modulated by YycFG two-component regulatory pathway. J. Orthop. Surg. Res. 2019, 14, 10. [Google Scholar] [CrossRef]
- O’Neill, E.; Pozzi, C.; Houston, P.; Smyth, D.; Humphreys, H.; Robinson, D.A.; O’Gara, J.P. Association between Methicillin Susceptibility and Biofilm Regulation in Staphylococcus aureus Isolates from Device-Related Infections. J. Clin. Microbiol. 2007, 45, 1379–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozzi, C.; Waters, E.; Rudkin, J.; Schaeffer, C.R.; Lohan, A.; Tong, P.; Loftus, B.; Pier, G.; Fey, P.D.; Massey, R.; et al. Methicillin Resistance Alters the Biofilm Phenotype and Attenuates Virulence in Staphylococcus aureus device-Associated Infections. PLoS Pathog. 2012, 8, e1002626. [Google Scholar] [CrossRef]
- Wu, S.; Liu, Y.; Lei, L.; Zhang, H. Antisense yycG modulates the susceptibility of Staphylococcus aureus to hydrogen peroxide via the sarA. BMC Microbiol. 2021, 21, 160. [Google Scholar] [CrossRef]
- Cheng, R.; Morcos, F.; Levine, H.; Onuchic, J.N. Toward rationally redesigning bacterial two-component signaling systems using coevolutionary information. Proc. Natl. Acad. Sci. USA 2014, 111, E563–E571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Cui, L.; Oguchi, A.; Aoki, K.; Nagai, Y.; et al. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Matsuo, M.; Kato, F.; Oogai, Y.; Kawai, T.; Sugai, M.; Komatsuzawa, H. Distinct two-component systems in methicillin-resistant Staphylococcus aureus can change the susceptibility to antimicrobial agents. J. Antimicrob. Chemother. 2010, 65, 1536–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulian, M. Two-component signaling circuit structure and properties. Curr. Opin. Microbiol. 2010, 13, 184–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Futur. Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef]
- Cameron, D.; Jiang, J.-H.; Kostoulias, X.; Foxwell, D.J.; Peleg, A.Y. Vancomycin susceptibility in methicillin-resistant Staphylococcus aureus is mediated by YycHI activation of the WalRK essential two-component regulatory system. Sci. Rep. 2016, 6, 30823. [Google Scholar] [CrossRef] [PubMed]
- Monk, I.R.; Shaikh, N.; Begg, S.L.; Gajdiss, M.; Sharkey, L.K.R.; Lee, J.Y.H.; Pidot, S.J.; Seemann, T.; Kuiper, M.; Winnen, B.; et al. Zinc-binding to the cytoplasmic PAS domain regulates the essential WalK histidine kinase of Staphylococcus aureus. Nat. Commun. 2019, 10, 3067. [Google Scholar] [CrossRef] [Green Version]
- Crosby, H.A.; Tiwari, N.; Kwiecinski, J.M.; Xu, Z.; Dykstra, A.; Jenul, C.; Fuentes, E.; Horswill, A.R. The Staphylococcus aureus ArlRS two-component system regulates virulence factor expression through MgrA. Mol. Microbiol. 2020, 113, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.; Zheng, F.; Chew, J.Y.; Pei, Y.; Zhou, J.; Wen, K.; Han, M.; Lemieux, M.J.; Hwang, P.M.; Wen, Y. Deciphering the activation and recognition mechanisms of Staphylococcus aureus response regulator ArlR. Nucleic Acids Res. 2019, 47, 11418–11429. [Google Scholar] [CrossRef]
- Tan, L.; Li, S.R.; Jiang, B.; Hu, X.M.; Li, S. Therapeutic targeting of the Staphylococcus aureus accessory gene regulator (agr) system. Front Microbiol. 2018, 9, 55. [Google Scholar] [CrossRef]
- Liu, L.; Shen, X.; Yu, J.; Cao, X.; Zhan, Q.; Guo, Y.; Yu, F. Subinhibitory Concentrations of Fusidic Acid May Reduce the Virulence of S. aureus by Down-Regulating sarA and saeRS to Reduce Biofilm Formation and α-Toxin Expression. Front. Microbiol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Valle, J.; Toledo-Arana, A.; Berasain, C.; Ghigo, J.M.; Amorena, B.; Penadés, J.R.; Lasa, I. SarA and not sigmaB is essential for biofilm development by Staphylococcus aureus. Mol. Microbiol. 2003, 48, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Tormo, M.A.; Martí, M.; Valle, J.; Manna, A.C.; Cheung, A.L.; Lasa, I.; Penadés, J.R. SarA Is an Essential Positive Regulator of Staphylococcus epidermidis Biofilm Development. J. Bacteriol. 2005, 187, 2348–2356. [Google Scholar] [CrossRef] [Green Version]
- Bayer, M.G.; Heinrichs, J.H.; Cheung, A.L. The molecular architecture of the sar locus in Staphylococcus aureus. J. Bacteriol. 1996, 178, 4563–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, A.L.; Eberhardt, K.; Heinrichs, J.H. Regulation of protein A synthesis by the sar and agr loci of Staphylococcus aureus. Infect. Immun. 1997, 65, 2243–2249. [Google Scholar] [CrossRef] [Green Version]
- Krismer, B.; Weidenmaier, C.; Zipperer, A.; Peschel, A. The commensal lifestyle of Staphylococcus aureus and its interactions with the nasal microbiota. Nat. Rev. Genet. 2017, 15, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Xu, D.; Xia, X.; Zhang, K.; Aadil, R.M.; Batool, Z.; Wang, J. Five major two components systems of Staphylococcus aureus for adaptation in diverse hostile environment. Microb. Pathog. 2021, 159, 105119. [Google Scholar] [CrossRef] [PubMed]
- Voyich, J.M.; Vuong, C.; Dewald, M.; Nygaard, T.; Kocianova, S.; Griffith, S.; Jones, J.; Iverson, C.; Sturdevant, D.E.; Braughton, K.R.; et al. The SaeR/S Gene Regulatory System Is Essential for Innate Immune Evasion by Staphylococcus aureus. J. Infect. Dis. 2009, 199, 1698–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karauzum, H.; Datta, S.K. Adaptive Immunity Against Staphylococcus aureus. Curr. Top. Microbiol. Immunol. 2017, 409, 419–439. [Google Scholar]
- Smith, E.J.; Visai, L.; Kerrigan, S.W.; Speziale, P.; Foster, T.J. The Sbi Protein Is a Multifunctional Immune Evasion Factor of Staphylococcus aureus. Infect. Immun. 2011, 79, 3801–3809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delauné, A.; Dubrac, S.; Blanchet, C.; Poupel, O.; Mäder, U.; Hiron, A.; LeDuc, A.; Fitting, C.; Nicolas, P.; Cavaillon, J.-M.; et al. The WalKR System Controls Major Staphylococcal Virulence Genes and Is Involved in Triggering the Host Inflammatory Response. Infect. Immun. 2012, 80, 3438–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moormeier, D.E.; Bose, J.L.; Horswill, A.R.; Bayles, K.W. Temporal and Stochastic Control of Staphylococcus aureus Biofilm Development. mBio 2014, 5, e01341-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuryastuti, T.; Krom, B.P. Ica-status of clinical Staphylococcus epidermidis strains affects adhesion and aggregation: A thermodynamic analysis. Antonie Van Leeuwenhoek 2017, 110, 1467–1474. [Google Scholar] [CrossRef]
- Schommer, N.N.; Christner, M.; Hentschke, M.; Ruckdeschel, K.; Aepfelbacher, M.; Rohde, H. Staphylococcus epidermidis Uses Distinct Mechanisms of Biofilm Formation To Interfere with Phagocytosis and Activation of Mouse Macrophage-Like Cells 774A.1. Infect. Immun. 2011, 79, 2267–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, V.; Matange, N. Adaptation and compensation in a bacterial gene regulatory network evolving under antibiotic selection. eLife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Fluckiger, U.; Ulrich, M.; Steinhuber, A.; Döring, G.; Mack, D.; Landmann, R.; Goerke, C.; Wolz, C. Biofilm Formation, icaADBC Transcription, and Polysaccharide Intercellular Adhesin Synthesis by Staphylococci in a Device-Related Infection Model. Infect. Immun. 2005, 73, 1811–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begun, J.; Gaiani, J.M.; Rohde, H.; Mack, D.; Calderwood, S.B.; Ausubel, F.M.; Sifri, C.D. Staphylococcal Biofilm Exopolysaccharide Protects against Caenorhabditis elegans Immune Defenses. PLoS Pathog. 2007, 3, e57. [Google Scholar] [CrossRef] [Green Version]
- Kropec, A.; Maira-Litran, T.; Jefferson, K.; Grout, M.; Cramton, S.E.; Götz, F.; Goldmann, D.A.; Pier, G.B. Poly- N -Acetylglucosamine Production in Staphylococcus aureus is Essential for Virulence in Murine Models of Systemic Infection. Infect. Immun. 2005, 73, 6868–6876. [Google Scholar] [CrossRef] [Green Version]
- Zago, C.E.; Silva, S.; Sanitá, P.V.; Barbugli, P.; Dias, C.M.I.; Lordello, V.B.; Vergani, C.E. Dynamics of Biofilm Formation and the Interaction between Candida albicans and Methicillin-Susceptible (MSSA) and -Resistant Staphylococcus aureus (MRSA). PLoS ONE 2015, 10, e0123206. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Hobby, G.L.; Meyer, K.; Chaffee, E. Observations on the Mechanism of Action of Penicillin. Exp. Biol. Med. 1942, 50, 281–285. [Google Scholar] [CrossRef]
- Waters, E.M.; Rowe, S.E.; O’Gara, J.P.; Conlon, B.P. Convergence of Staphylococcus aureus persister and biofilm research: Can biofilms be defined as communities of adherent persister cells? PLoS Pathog. 2016, 12, e1006012. [Google Scholar] [CrossRef] [PubMed]
- Schwartbeck, B.; Birtel, J.; Treffon, J.; Langhanki, L.; Mellmann, A.; Kale, D.; Kahl, J.; Hirschhausen, N.; Neumann, C.; Lee, J.; et al. Dynamic in vivo mutations within the ica operon during persistence of Staphylococcus aureus in the airways of cystic fibrosis patients. PLoS Pathog. 2016, 12, e1006024. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, H.; Pourhajibagher, M.; Chiniforush, N.; Soltanian, A.R.; Alikhani, M.Y.; Bahador, A. Biofilm formation and antibiotic resistance in meticillin-resistant and meticillin-sensitive Staphylococcus aureus isolated from burns. J. Wound Care 2019, 28, 66–73. [Google Scholar] [CrossRef]
- Kıvanç, S.A.; Arık, G.; Akova-Budak, B.; Kıvanç, M. Biofilm forming capacity and antibiotic susceptibility of Staphylococcus spp. with the icaA/icaD/bap genotype isolated from ocular surface of patients with diabetes. Malawi Med. J. 2018, 30, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Melo-Cristino, J.; Resina, C.; Manuel, V.; Lito, L.; Ramirez, M. First case of infection with vancomycin-resistant Staphylococcus aureus in Europe. Lancet 2013, 382, 205. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.; Eichenberger, E.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Genet. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Kourtis, A.P.; Hatfield, K.; Baggs, J.; Mu, Y.; See, I.; Epson, E.; Nadle, J.; Kainer, M.A.; Dumyati, G.; Petit, S.; et al. Vital Signs:Epidemiology and Recent Trends in Methicillin-Resistant and in Methicillin-Susceptible Staphylococcus aureus bloodstream Infections—United States. MMWR. Morb. Mortal. Wkly. Rep. 2019, 68, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Rajagopal, M.; Martin, M.J.; Santiago, M.; Lee, W.; Kos, V.N.; Meredith, T.; Gilmore, M.S.; Walker, S. Multidrug Intrinsic Resistance Factors in Staphylococcus aureus Identified by Profiling Fitness within High-Diversity Transposon Libraries. mBio 2016, 7, e00950-16. [Google Scholar] [CrossRef] [Green Version]
- Coe, K.A.; Lee, W.; Stone, M.C.; Komazin-Meredith, G.; Meredith, T.C.; Grad, Y.H.; Walker, S. Multi-strain Tn-Seq reveals common daptomycin resistance determinants in Staphylococcus aureus. PLoS Pathog. 2019, 15, e1007862. [Google Scholar] [CrossRef] [Green Version]
- Berti, A.D.; Baines, S.; Howden, B.; Sakoulas, G.; Nizet, V.; Proctor, R.A.; Rose, W.E. Heterogeneity of Genetic Pathways toward Daptomycin Nonsusceptibility in Staphylococcus aureus Determined by Adjunctive Antibiotics. Antimicrob. Agents Chemother. 2015, 59, 2799–2806. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Liu, Y.; Lei, L.; Zhang, H. Antisense yycG Regulation of Antibiotic Sensitivity of Methicillin-Resistant Staphylococcus aureus in Chronic Osteomyelitis. Surg. Infect. (Larchmt). 2019, 20, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, Y.; Lei, L.; Zhang, H. Virulence of methicillin-resistant Staphylococcus aureus modulated by the YycFG two-component pathway in a rat model of osteomyelitis. J. Orthop. Surg. Res. 2019, 14, 433. [Google Scholar] [CrossRef] [Green Version]
- Delaune, A.; Poupel, O.; Mallet, A.; Coic, Y.M.; Msadek, T.; Dubrac, S. Peptidoglycan crosslinking relaxation plays an important role in Staphylococcus aureus WalKR-dependent cell viability. PLoS ONE 2011, 6, e17054. [Google Scholar] [CrossRef] [Green Version]
- Dubrac, S.; Bisicchia, P.; Devine, K.M.; Msadek, T. A matter of life and death: Cell wall homeostasis and the WalKR (YycGF) essential signal transduction pathway. Mol. Microbiol. 2008, 70, 1307–1322. [Google Scholar] [CrossRef]
- Peng, H.; Rao, Y.; Yuan, W.; Zheng, Y.; Shang, W.; Hu, Z.; Yang, Y.; Tan, L.; Xiong, K.; Li, S.; et al. Reconstruction of the Vancomycin-Susceptible Staphylococcus aureus Phenotype From a Vancomycin-Intermediate S. aureus XN108. Front. Microbiol. 2018, 9, 2955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howden, B.P.; McEvoy, C.R.E.; Allen, D.L.; Chua, K.; Gao, W.; Harrison, P.; Bell, J.; Coombs, G.; Bennett-Wood, V.; Porter, J.L.; et al. Evolution of Multidrug Resistance during Staphylococcus aureus Infection Involves Mutation of the Essential Two Component Regulator WalKR. PLoS Pathog. 2011, 7, e1002359. [Google Scholar] [CrossRef] [PubMed]
- Jansen, A.; Türck, M.; Szekat, C.; Nagel, M.; Clever, I.; Bierbaum, G. Role of insertion elements and yycFG in the development of decreased susceptibility to vancomycin in Staphylococcus aureus. Int. J. Med. Microbiol. 2007, 297, 205–215. [Google Scholar] [CrossRef]
- Dubrac, S.; Boneca, I.G.; Poupel, O.; Msadek, T. New Insights into the WalK/WalR (YycG/YycF) Essential Signal Transduction Pathway Reveal a Major Role in Controlling Cell Wall Metabolism and Biofilm Formation in Staphylococcus aureus. J. Bacteriol. 2007, 189, 8257–8269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, P.C.; Barry, A.L.; Brown, S.D. In vitro bactericidal activity of daptomycin against staphylococci. J. Antimicrob. Chemother. 2002, 49, 467–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, A.S.; Schneider, T.; Sahl, H.-G. Mechanisms of daptomycin resistance in Staphylococcus aureus: Role of the cell membrane and cell wall. Ann. N. Y. Acad. Sci. 2012, 1277, 139–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paharik, A.E.; Horswill, A.R. The Staphylococcal Biofilm: Adhesins, Regulation, and Host Response. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, N.N.; McKinnell, J.; Yeaman, M.R.; Rubio, A.; Nast, C.C.; Chen, L.; Kreiswirth, B.N.; Bayer, A.S. In VitroCross-Resistance to Daptomycin and Host Defense Cationic Antimicrobial Peptides in Clinical Methicillin-Resistant Staphylococcus aureus Isolates. Antimicrob. Agents Chemother. 2011, 55, 4012–4018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasch, O.; Camoez, M.; Domínguez, M.A.; Padilla, B.; Pintado, V.; Almirante, B.; Martín, C.; López-Medrano, F.; de Gopegui, E.R.; Blanco, J.R.; et al. REIPI/GEIH study groups. Emergence of resistance to daptomycin in a cohort of patients with methicillin-resistant Staphylococcus aureus persistent bacteraemia treated with daptomycin. J. Antimicrob. Chemother. 2014, 69, 568–571. [Google Scholar] [CrossRef] [Green Version]
- Mitrophanov, A.Y.; Groisman, E.A. Signal integration in bacterial two-component regulatory systems. Genes Dev. 2008, 22, 2601–2611. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, A.M.; Beenken, K.E.; Byrum, S.D.; Tackett, A.J.; Shaw, L.N.; Gimza, B.D.; Smeltzer, M.S. SarA plays a predominant role in controlling the production of extracellular proteases in the diverse clinical isolates of Staphylococcus aureus LAC and UAMS-1. Virulence 2020, 11, 1738–1762. [Google Scholar] [CrossRef]
- Yu, J.; Jiang, F.; Zhang, F.; Pan, Y.; Wang, J.; Han, P.; Tang, J.; Shen, H. Virtual Screening for Novel SarA Inhibitors to Prevent Biofilm Formation of Staphylococcus aureus in Prosthetic Joint Infections. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Valverde, C.; Haas, D. Small RNAs Controlled by Two-Component Systems. Adv. Exp. Med. Biology 2008, 631, 54–79. [Google Scholar] [CrossRef]
- Brantl, S.; Müller, P. Cis- and Trans-Encoded Small Regulatory RNAs in Bacillus subtilis. Microorganisms 2021, 9, 1865. [Google Scholar] [CrossRef]
- Thomason, M.K.; Storz, G. Bacterial Antisense RNAs: How Many Are There, and What Are They Doing? Annu. Rev. Genet. 2010, 44, 167–188. [Google Scholar] [CrossRef] [Green Version]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Eckert, M.R.; Vogel, J.; Charpentier, E. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, L.; Stipp, R.; Chen, T.; Wu, S.; Hu, T.; Duncan, M. Activity of Streptococcus mutans VicR Is Modulated by Antisense RNA. J. Dent. Res. 2018, 97, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Jani, S.; Ramirez, M.; Tolmasky, M. Silencing Antibiotic Resistance with Antisense Oligonucleotides. Biomedicines 2021, 9, 416. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, Y.; Lei, L.; Zhang, H. An Antisense yycF RNA Modulates Biofilm Organization of Methicillin-Resistant Staphylococcus aureus and Pathogenicity in a Rat Model of Osteomyelitis. Antibiotics 2021, 10, 603. [Google Scholar] [CrossRef] [PubMed]
- Türck, M.; Bierbaum, G. Purification and Activity Testing of the Full-Length YycFGHI Proteins of Staphylococcus aureus. PLoS ONE 2012, 7, e30403. [Google Scholar] [CrossRef]
- Figueiredo, A.M.S.; Ferreira, F.A.; Beltrame, C.O.; Côrtes, M.F. The role of biofilms in persistent infections and factors involved in ica-independent biofilm development and gene regulation in Staphylococcus aureus. Crit. Rev. Microbiol. 2017, 43, 602–620. [Google Scholar] [CrossRef] [PubMed]
- Schilcher, K.; Horswill, A.R. Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies. Microbiol. Mol. Biol. Rev. 2020, 84, e00026-19. [Google Scholar] [CrossRef] [PubMed]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus Virulence. Microbiol. Spectr. 2019, 7, 10. [Google Scholar] [CrossRef]
- Loughran, A.J.; Atwood, D.N.; Anthony, A.C.; Harik, N.S.; Spencer, H.J.; Beenken, K.E.; Smeltzer, M.S. Impact of individual extracellular proteases on Staphylococcus aureus biofilm formation in diverse clinical isolates and their isogenic sarA mutants. MicrobiologyOpen 2014, 3, 897–909. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.-W.; Cho, H.; Jones, M.B.; Shatzkes, K.; Sun, F.; Ji, Q.; Liu, Q.; Peterson, S.N.; He, C.; Bae, T. The auxiliary protein complex SaePQ activates the phosphatase activity of sensor kinase SaeS in the SaeRS two-component system ofStaphylococcus aureus. Mol. Microbiol. 2012, 86, 331–348. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Zhang, J.; Peng, Q.; Liu, Y.; Lei, L.; Zhang, H. The Role of Staphylococcus aureus YycFG in Gene Regulation, Biofilm Organization and Drug Resistance. Antibiotics 2021, 10, 1555. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121555
Wu S, Zhang J, Peng Q, Liu Y, Lei L, Zhang H. The Role of Staphylococcus aureus YycFG in Gene Regulation, Biofilm Organization and Drug Resistance. Antibiotics. 2021; 10(12):1555. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121555
Chicago/Turabian StyleWu, Shizhou, Junqi Zhang, Qi Peng, Yunjie Liu, Lei Lei, and Hui Zhang. 2021. "The Role of Staphylococcus aureus YycFG in Gene Regulation, Biofilm Organization and Drug Resistance" Antibiotics 10, no. 12: 1555. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121555