
Effects of the Licorice Isoflavans Licoricidin and Glabridin on the Growth, Adherence Properties, and Acid Production of Streptococcus mutans, and Assessment of Their Biocompatibility

Abstract
:1. Introduction
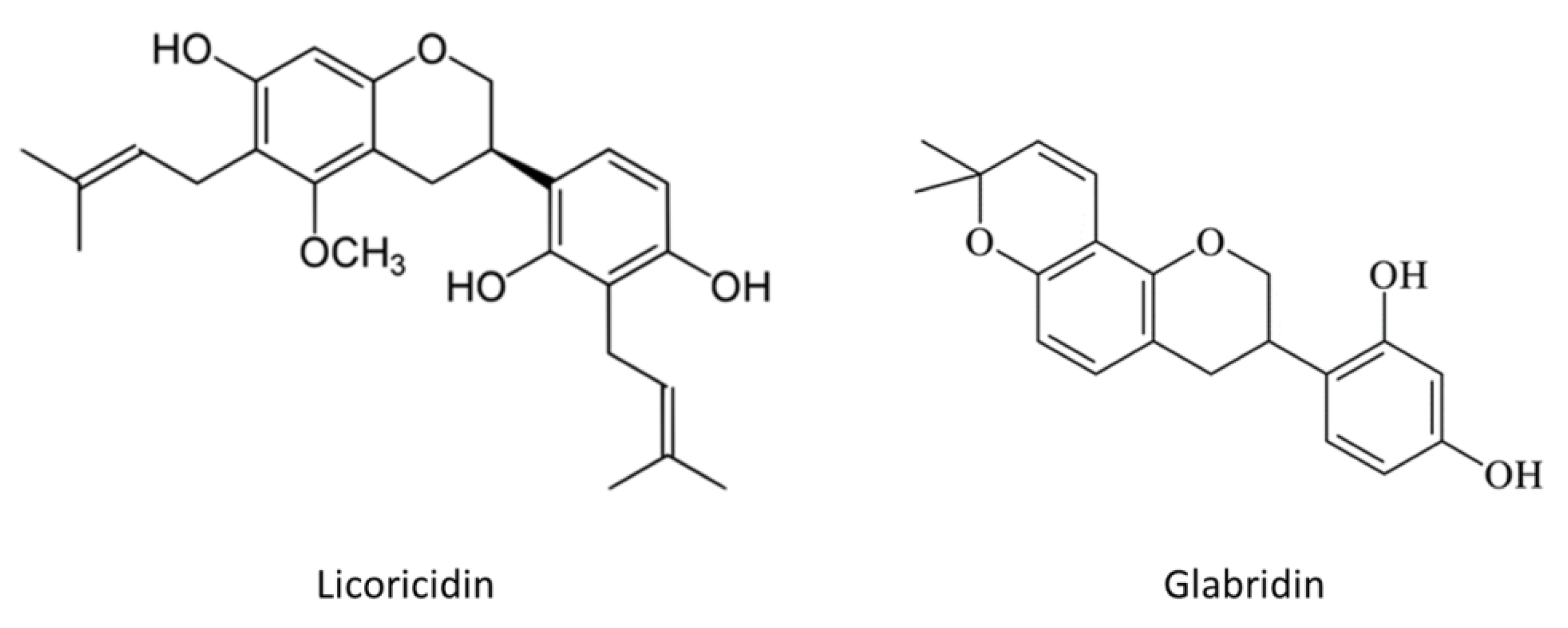
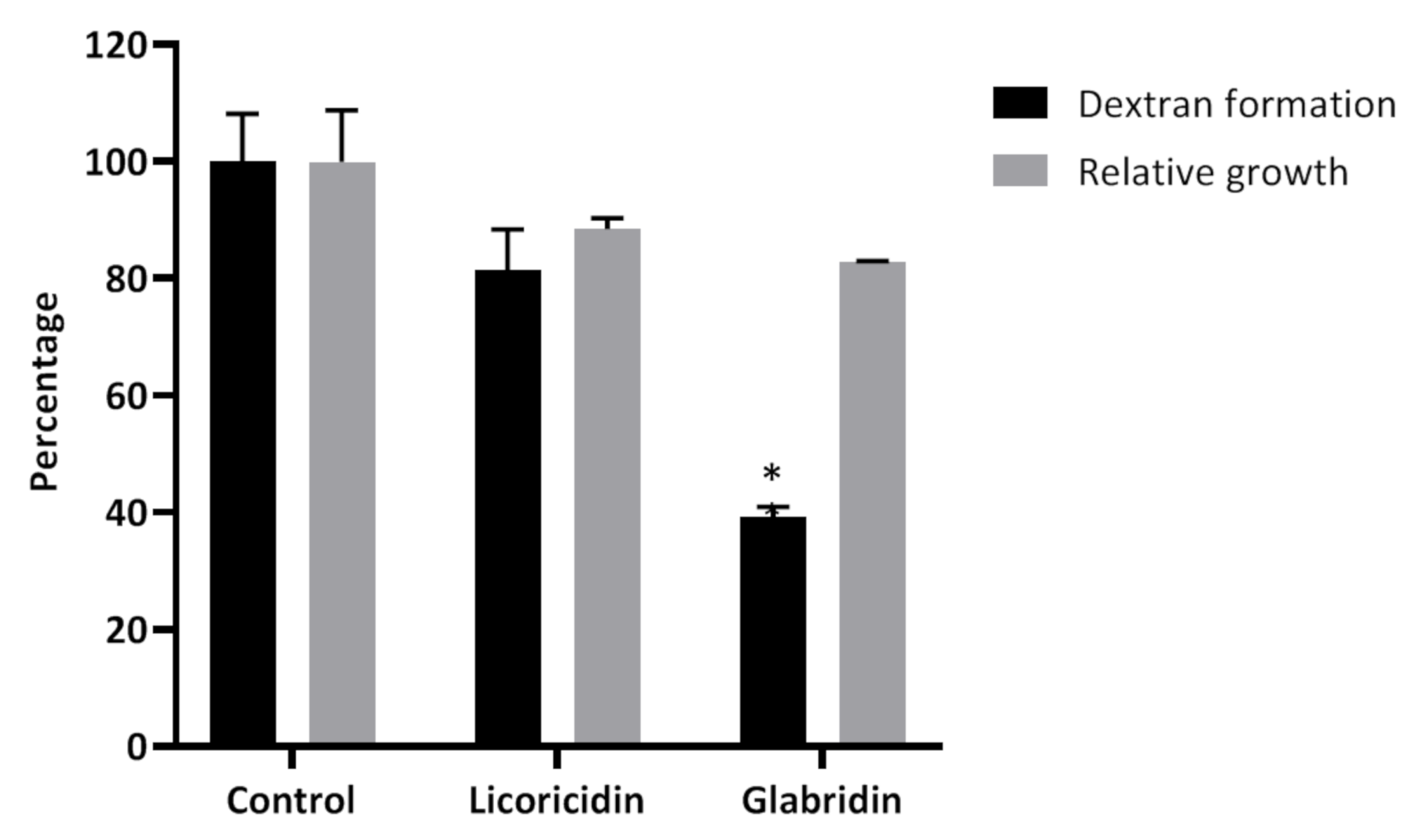
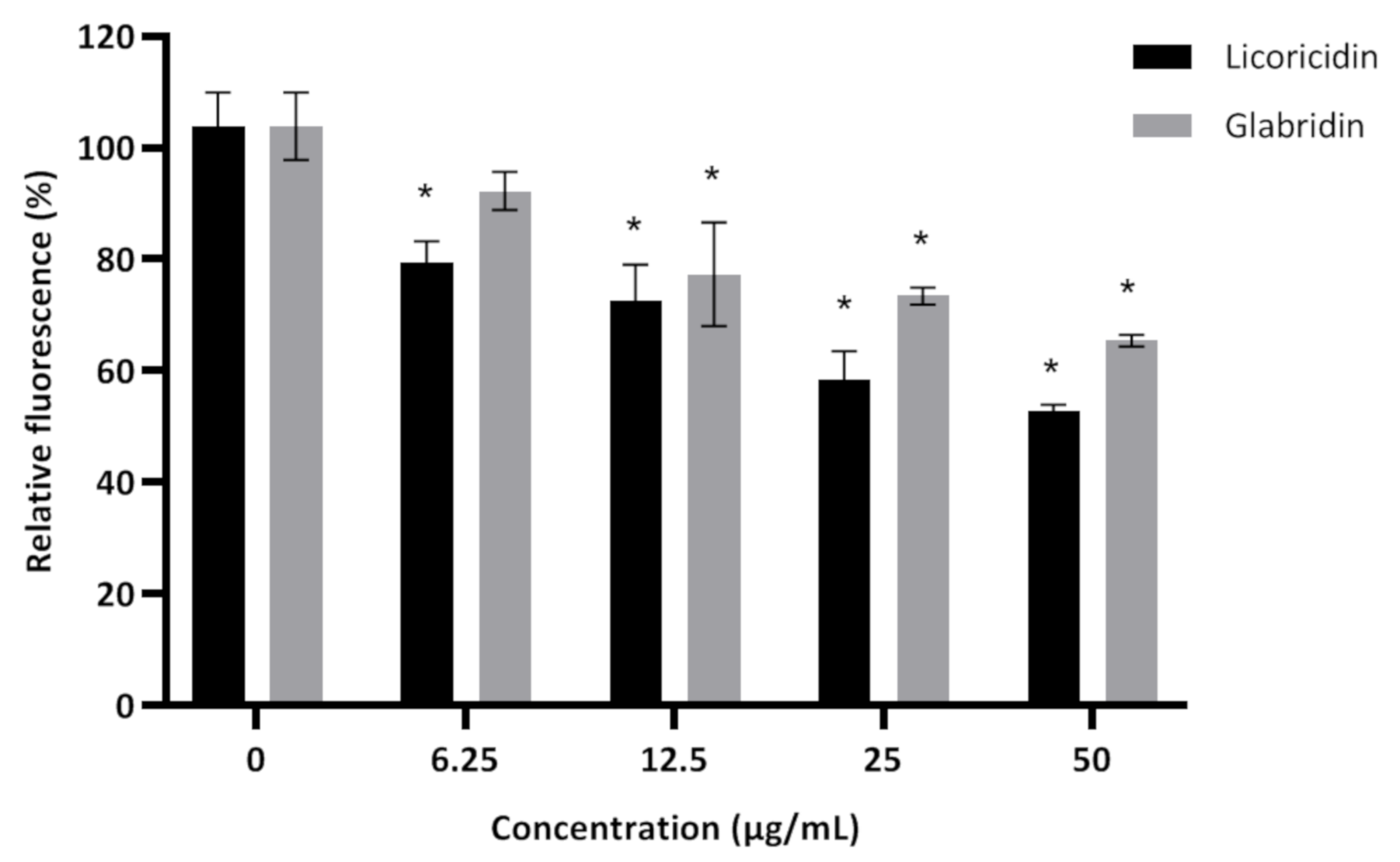
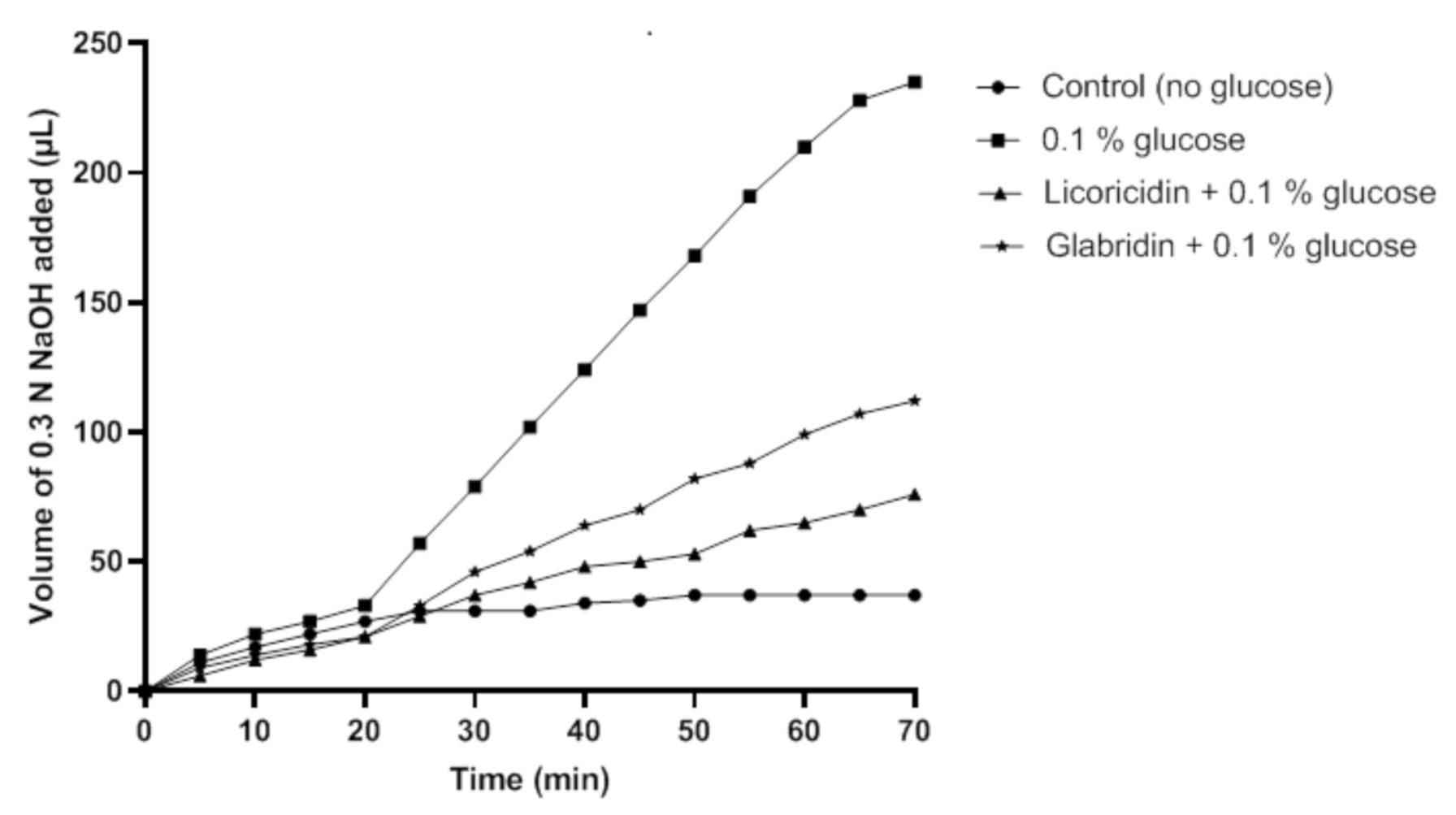
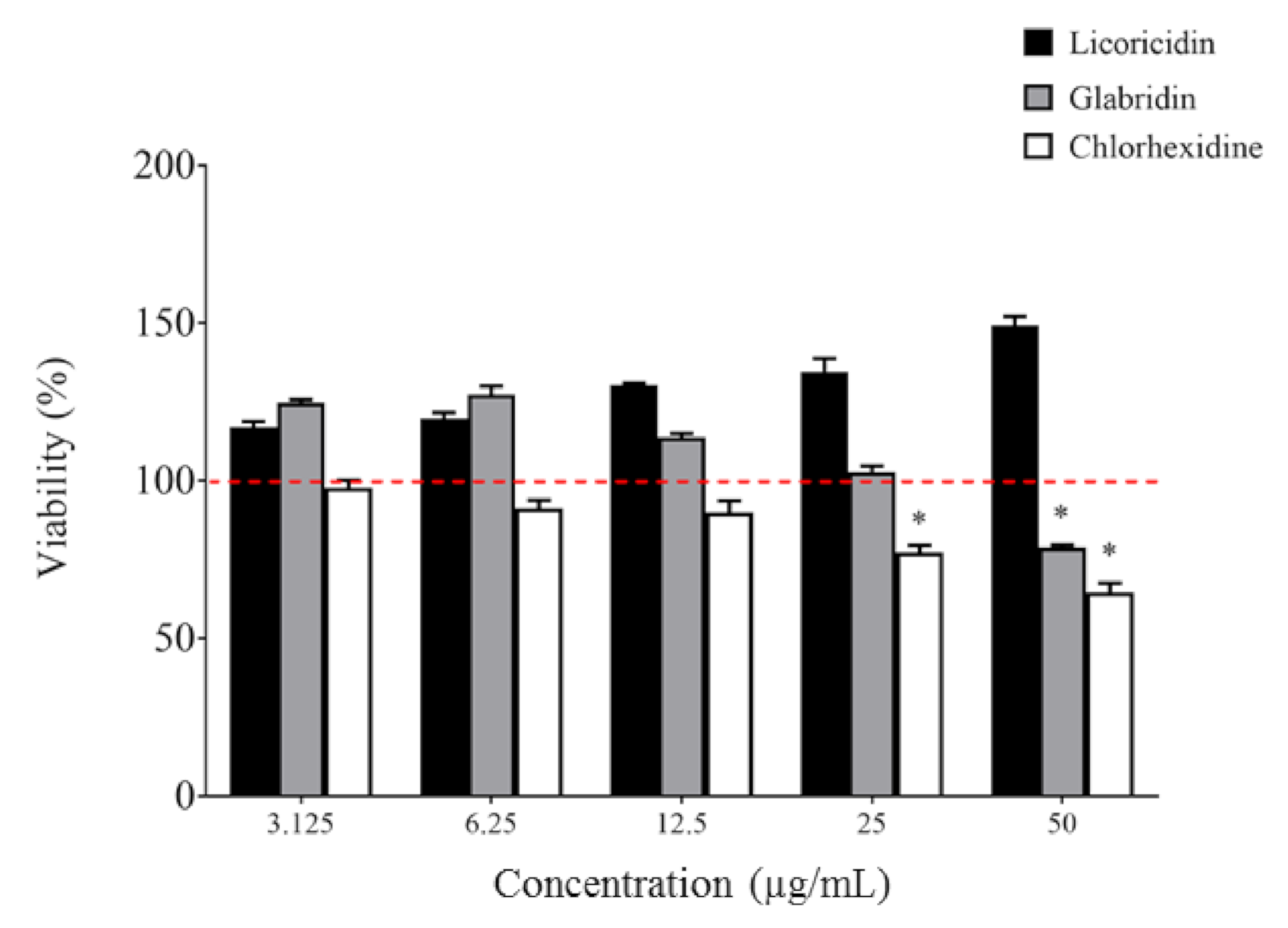
2. Results and Discussion
3. Materials and Methods
3.1. Compounds
3.2. Bacteria and Growth Conditions
3.3. Determination of Minimum Inhibitory and Minimum Bactericidal Concentrations
3.4. Biofilm Biomass and Viability
3.5. Dextran Formation
3.6. Adherence to Hydroxylapatite
3.7. Glycolytic pH Drop Assay
3.8. In Vitro Biocompatibility
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pitts, N.B.; Zero, D.T.; Marsh, P.D.; Ekstrand, K.; Weintraub, J.A.; Ramos-Gomez, F.; Tagami, J.; Twetman, S.; Tsakos, G.; Ismail, A. Dental caries. Nat. Rev. Dis. Primers 2017, 3, 17030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, N.; Nyvad, B. The role of bacteria in the caries process: Ecological perspectives. J. Dent. Res. 2011, 90, 294–303. [Google Scholar] [CrossRef]
- Koo, H.; Xiao, J.; Klein, M.I. Extracellular polysaccharides matrix—An often forgotten virulence factor in oral biofilm research. Int. J. Oral Sci. 2009, 1, 229–234. [Google Scholar] [CrossRef]
- Addy, M. Oral hygiene products: Potential for harm to oral and systemic health? Periodontol. 2000 2008, 48, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, G.M.; Kumar, S.; Fornari, C.D.; Corti, E.; Connelly, S.T. Mouthwashes in the 21st century: A narrative review about active molecules and effectiveness on the periodontal outcomes. Expert Opin. Drug Deliv. 2017, 14, 973–982. [Google Scholar] [CrossRef] [Green Version]
- Tartaglia, G.M.; Tadakamadla, S.K.; Connelly, S.T.; Sforza, C.; Martin, C. Adverse events associated with home use of mouthrinses: A systematic review. Ther. Adv. Drug Saf. 2019, 10, 1–6. [Google Scholar] [CrossRef]
- Zanatta, F.B.; Antoniazzi, R.P.; Rosing, C.K. Staining and calculus formation after 0.12% chlorhexidine rinses in plaque-free and plaque covered surfaces: A randomized trial. J. Appl. Oral Sci. 2010, 18, 515–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Daliri, E.B.M.; Kim, N.; Kim, J.R.; Yoo, D.; Oh, D.H. Microbial etiology and prevention of dental caries: Exploiting natural products to inhibit cariogenic biofilms. Pathogens 2020, 9, 569. [Google Scholar] [CrossRef]
- Jeon, J.G.; Rosalen, P.L.; Falsetta, M.L.; Koo, H. Natural products in caries research: Current (limited) knowledge, challenges and future perspective. Caries Res. 2011, 45, 243–263. [Google Scholar] [CrossRef]
- Davis, E.A.; Morris, D.J. Medicinal uses of licorice through the millennia: The good and plenty of it. Mol. Cell Endocrinol. 1991, 78, 1–6. [Google Scholar] [CrossRef]
- Jiang, M.; Zhao, S.; Yang, S.; Lin, X.; He, X.; Wei, X.; Song, Q.; Li, R.; Fu, C.; Zhang, J.; et al. An “essential herbal medicine”—Licorice: A review of phytochemicals and its effects in combination preparations. J. Ethnopharmacol. 2020, 249, 112439. [Google Scholar] [CrossRef]
- Isbrucker, R.A.; Burdock, G.A. Risk and safety assessment on the consumption of licorice root (Glycyrrhiza sp.), its extract and powder as a food ingredient, with emphasis on the pharmacology and toxicology of glycyrrhizin. Reg. Toxicol. Pharmacol. 2006, 26, 167–192. [Google Scholar] [CrossRef]
- Yang, R.; Wang, L.Q.; Yuan, B.C.; Liu, Y. The pharmacological activities of licorice. Planta Med. 2015, 81, 1654–1669. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, J.; Zhou, W.; Wang, Y.; Yang, L. Systems approaches and polypharmacology for drug discovery from herbal medicines: An example using licorice. J. Ethnopharmacol. 2013, 146, 773–793. [Google Scholar] [CrossRef]
- Nassiri Asl, M.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar]
- Messier, C.; Epifano, F.; Genovese, S.; Grenier, D. Licorice and its potential beneficial effects in common oro-dental diseases. Oral Dis. 2012, 18, 32–38. [Google Scholar] [CrossRef]
- He, J.; Chen, L.; Heber, D.; Shi, W.; Lu, Q.Y. Antibacterial compounds from Glycyrrhiza uralensis. J. Nat. Prod. 2006, 69, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Nuvvula, S.; Nunna, M.; Almaz, M.E.; Mallineni, S.K. Efficacy of licorice lollipops in reducing dental caries in a paediatric population: A systematic review. Oral Health Prev. Dent. 2020, 18, 97–102. [Google Scholar]
- Segal, R.; Pisanty, S.; Wormser, R.; Azaz, E.; Sela, M.N. Anticariogenic activity of licorice and glycyrrhizine I: Inhibition of in vitro plaque formation by Streptococcus mutans. J. Pharm. Sci. 1985, 74, 79–81. [Google Scholar] [CrossRef]
- Sela, M.N.; Steinberg, D.; Segal, R. Inhibition of the activity of glucosyltransferase from Streptococcus mutans by glycyrrhizin. Oral Microbiol. Immunol. 1987, 2, 125–128. [Google Scholar] [CrossRef]
- Tanabe, S.; Desjardins, J.; Bergeron, C.; Gafner, S.; Villinski, J.R.; Grenier, D. Reduction of bacterial volatile sulfur compound production by licoricidin and licorisoflavan A from licorice. J. Breath Res. 2012, 6, 016006. [Google Scholar] [CrossRef] [PubMed]
- Grenier, D.; Marcoux, E.; Azelmat, J.; Ben Lagha, A.; Gauthier, P. Biocompatible combinations of nisin and licorice polyphenols exert synergistic bactericidal effects against Enterococcus faecalis and inhibit NF-κB activation in monocytes. AMB Expr. 2020, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Araya-Cloutier, C.; Vincken, J.P.; van de Schans, M.G.M.; Hageman, J.; Schaftenaar, G.; den Besten, H.M.W.; Gruppen, H. QSAR-based molecular signatures of prenylated (iso)flavonoids underlying antimicrobial potency against and membrane-disruption in Gram positive and Gram negative bacteria. Sci. Rep. 2018, 8, 9267. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Park, S.N.; Lee, Y.J.; Cho, E.J.; Lim, Y.K.; Li, X.M.; Choi, M.H.; Seo, Y.W.; Kook, J.K. In vitro antimicrobial activities of 1-methoxyficifolinol, licorisoflavan A, and 6,8-diprenylgenistein against Streptococcus mutans. Caries Res. 2015, 49, 78–89. [Google Scholar] [CrossRef]
- La, V.D.; Tanabe, S.I.; Bergeron, C.; Gafner, S.; Grenier, D. Modulation of matrix metalloproteinase and cytokine production by licorice isolates licoricidin and licorisoflavan A: Potential therapeutic approach for periodontitis. J. Periodontol. 2011, 82, 122–128. [Google Scholar] [CrossRef]
- National Committee for Clinical Laboratory Standards. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Document M7-A6; NCCLS: Wayne, PA, USA, 2003. [Google Scholar]
- LeBel, G.; Vaillancourt, K.; Bercier, P.; Grenier, D. Antibacterial activity against porcine respiratory bacterial pathogens and in vitro biocompatibility of essential oils. Arch. Microbiol. 2019, 201, 833–840. [Google Scholar] [CrossRef]
- Drago, L.; Agrappi, S.; Bortolin, M.; Toscano, M.; Romano, C.L.; De Vecchi, E. How to study biofilms after microbial colonization of materials used in orthopaedic implants. Int. J. Mol. Sci. 2016, 17, 293. [Google Scholar] [CrossRef]
- Ben Lagha, A.; Haas, B.; Grenier, D. Tea polyphenols inhibit the growth and virulence properties of Fusobacterium nucleatum. Sci. Rep. 2017, 7, 44805. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, M.; Millhouse, E.; Culshaw, S.; Edwards, C.A.; Ramage, G.; Comber, E. Selected dietary (poly)phenols inhibit periodontal pathogen growth and biofilm formation. Food Funct. 2015, 6, 719–729. [Google Scholar] [CrossRef] [Green Version]
- Groeger, S.; Michel, J.; Meyle, J. Establishment and characterization of immortalized human gingival keratinocyte cell lines. J. Periodont. Res. 2008, 43, 604–614. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | S. mutans Strains | ||||
|---|---|---|---|---|---|
| 25175 | 12A | 33A | INB | T8 | |
| Licoricidin | |||||
| MIC (µg/mL) | 6.25 | 6.25 | 6.25 | 6.25 | 6.25 |
| MBC (µg/mL) | 25 | 6.25 | 12.5 | 12.5 | 25 |
| Glabridin | |||||
| MIC (µg/mL) | 12.5 | 6.25 | 12.5 | 12.5 | 12.5 |
| MBC (µg/mL) | 25 | 6.25 | 12.5 | 25 | 25 |
| Strains | Biofilm Viability | ||||
|---|---|---|---|---|---|
| Control | Licoricidin | Glabridin | |||
| MIC | 2× MIC | MIC | 2× MIC | ||
| 25175 | 100 ± 0.4 | 98.1 ± 0.6 | 92.7 ± 2.1 * | 93.7 ± 1.6 * | 82.8 ± 1.3 * |
| 12A | 100 ± 1.6 | 100.2 ± 0.5 | 97.3 ± 1.1 | 100 ± 0.9 | 95.9 ± 1.4 |
| 33A | 100 ± 1.5 | 91.5 ± 1.6 * | 84.6 ± 1.9 * | 82.7 ± 5.4 * | 71.0 ± 2.8 * |
| INB | 100 ± 2.1 | 96.1 ± 2.1 | 92.8 ± 2.2 | 92.4 ± 0.9 | 87.1 ± 1.8 * |
| T8 | 100 ± 2.0 | 99.6 ± 1.1 | 98.1 ± 1.3 | 94.9 ± 1.7 | 82.5 ± 0.8 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaillancourt, K.; LeBel, G.; Pellerin, G.; Ben Lagha, A.; Grenier, D. Effects of the Licorice Isoflavans Licoricidin and Glabridin on the Growth, Adherence Properties, and Acid Production of Streptococcus mutans, and Assessment of Their Biocompatibility. Antibiotics 2021, 10, 163. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020163
Vaillancourt K, LeBel G, Pellerin G, Ben Lagha A, Grenier D. Effects of the Licorice Isoflavans Licoricidin and Glabridin on the Growth, Adherence Properties, and Acid Production of Streptococcus mutans, and Assessment of Their Biocompatibility. Antibiotics. 2021; 10(2):163. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020163
Chicago/Turabian StyleVaillancourt, Katy, Geneviève LeBel, Geneviève Pellerin, Amel Ben Lagha, and Daniel Grenier. 2021. "Effects of the Licorice Isoflavans Licoricidin and Glabridin on the Growth, Adherence Properties, and Acid Production of Streptococcus mutans, and Assessment of Their Biocompatibility" Antibiotics 10, no. 2: 163. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020163