2. Results and Discussion
In the current study, the quantitative analysis of the total flavonoid and phenolic contents of methanolic, water, and ethyl acetate extracts of leaves and stem bark of
B. ferruginea was carried out using standard colorimetric assays. For instance, clearly the extracts of the leaves of
B. ferruginea contained higher amounts of total flavonoids (24.37–42.31 mg/g) compared to the stem bark extracts (2.05–2.62 mg/g). However, the methanolic extract of the leaves showed the highest content of total flavonoids, followed by the leaves ethyl acetate and water extracts. Additionally, the extracts were found to possess relatively moderate to significant total phenolic content ranging from 26.07–193.58 mg/g. Notably, the methanolic and water extracts of the stem bark contained the highest phenolic content (193.58 ± 0.98 and 187.84 ± 1.88 mg/g, respectively), while the least phenolic contents were observed in the ethyl acetate extracts of both the leaves and stem bark, respectively (26.07 ± 0.48 and 26.90 ± 1.12 mg/g). Moreover, the methanolic and water extracts of the leaves were found to contain noteworthy phenolic levels (103.94 ± 2.00 and 85.05 ± 0.58, respectively) (
Table 1). The total phenolic and flavonoid contents of
n-hexane, ethyl acetate, and butanolic and residual aqueous fractions of
B. ferruginae leaves have also been investigated, with
n-hexane fraction yielding the highest phenolic and flavonoid contents [
14].
Indeed, extraction procedures and solvents greatly influence the solubility of the plants’ endogenous compounds. Moreover, plant components can be non-polar or polar in nature. Particularly, phenolic compounds are more soluble in polar organic solvents owing to the presence of a hydroxyl group, and particularly, methanol is used commonly as a preferred solvent for extraction [
15]. Interestingly, in the present study also, methanol was found to extract the highest phenolic and flavonoid contents from stem bark and leaves, respectively.
UHPLC/MS/MS technique was used for the chemical analysis of the different extracts, given that it provides high efficiency chromatographic separation, selectivity, and sensitivity, and the Orbitrap mass spectrometer allows precise mass measurement in order to characterize plant metabolites. UHPLC-MS analyses of all extracts were carried out in two chromatographic runs with recording of mass spectra in positive and negative ion mode, and protonated [M + H]+ or deprotonated [M − H]− molecules and their fragments were recorded. Many compounds having phenolic hydroxyl group(s) found in the extracts were ionized preferentially in negative electrospray ionization mode (ESI). In some cases, data recorded in positive ion mode were used to characterize components. The fragmentation of flavonoids in MS/MS is structure-specific, and thus can be useful for detection and/or determination of flavonoid structures. For instance, the loss of m/z 162 indicates O-hexosides, whereas the loss of m/z 120 suggests C-glycosides.
The number of characterized compounds and chemical composition in methanolic and aqueous extracts of stem bark and leaves of
B. ferruginea were similar (
Table 2). Details about metabolomic analysis are provided as
Supplementary Materials (Tables S1–S6; Figures S1–S7). Thirty-eight compounds from methanolic and thirty-nine compounds from aqueous extracts of stem bark were identified, while 48 and 38 compounds were characterized from methanolic and aqueous extracts of the leaves. Only 25 and 26 components were characterized from ethyl acetate extracts of stem bark and leaves (in
Supplemental Materials).
Organic acids (gallic, pantothenic, kynurenic, and chlorogenic), catechins (gallocatechin, catechin, epigallocatechin, epigallocatechin-3-O-gallate, and epicatechin), flavonoids (myricetin, apigenin, quercetin, luteolin, and kaempferol) and flavone derivatives (hydroxy, methoxy, rhamnosyl, C- and O-glucosides, and rutinoside) and their isomers were assigned in the extracts.
Ellagic acid (RT: 23.94 min) and its derivatives (O-hexoside 20.34 min, O-pentoside 23.27 min, ellagic acid-4-O-rhamnoside 23.58 min, 3-O-methyl-4′-O-xyloside 25.26 min and eschweilenol A or isomer 26.39 min) could be separated and characterized. All compounds showed characteristic product ions at 300,999, corresponding to the elimination of a molecule of hexose, and O-methyl derivatives produced fragment ions, corresponding to the loss of methyl group.
A typical extracted ion chromatogram of these compounds: a: ellagic acid; b: ellagic acid-O-hexoside; c: ellagic acid-O-pentoside; d: eschweilenol C (ellagic acid-4-O-rhamnoside); ducheside A (3-O-methylellagic acid-4′-O-xyloside); and e: eschweilenol A or isomer and their MS2 spectra in negative ion mode are shown in
Supplemental figures.
Phenolics and flavonoids are important antioxidant compounds present in plants. Particularly, the antioxidant activity of phenolics is attributed mainly because of their redox properties, enabling them to perform as reducing agents, in addition to donors of hydrogen atom. Furthermore, natural antioxidants are able to act as chain breakers, free-radical scavengers, complexers of pro-oxidant metal ions, as well as quenchers of singlet-oxygen formation [
16]. Similarly, the potential health benefits of dietary flavonoids have been garnering appreciable interests owing to their multiple beneficial effects. Indeed, several epidemiological evidences have established the recurrent intake of flavonoid-rich foods and beverages to play a significant role in the reduction or prevention of several ailments. For instance, they are known to exhibit antiviral, antimicrobial, antiatherosclerosis, cardioprotective, anti-neoplastic, anti-ulcerogenic, antioxidant, antidiabetic, mutagenic, antiinflammatory, anti-aging, anti-hepatotoxic, hypolipidaemic, anti-hypertensive, and anti-platelet effects amongst others [
17]. The mechanisms by which flavonoids display their therapeutic effects include the capacity to scavenge an array of reactive oxygen, chlorine, and nitrogen species such as superoxide as well as hydroxyl and peroxyl radicals, including hypochlorous and peroxynitrous acids. They also chelate ions, often reducing the metal ion pro-oxidant activity [
18,
19,
20,
21]. In fact, the pharmacological properties of flavonoids have been attributed to its antioxidant property [
22].
Additionally, several studies have revealed the presence of a correlation between total phenolic and flavonoid contents and antioxidant capacity [
23,
24,
25]. Importantly, it has also been reported that as antioxidants, phenolics and flavonoids are capable of exerting diverse biological effects such as antiallergic, anti-inflammatory, antidiabetic, antimicrobial, antipathogenic, antiviral, antithrombotic, and vasodilatory effects and averting disease conditions like cancer, Alzheimer’s disease, heart problems, cataracts, and eye disorders [
26].
The production of highly reactive oxygen species (ROS) induces oxidative stress, which plays a fundamental role in the pathogenesis of many physiological disorders, including cell injury, cancer, and hepatic, heart, neurodegenerative, and renal disorders [
15]. Thus, given the increasing risk factors of humans to different types of deadly diseases, there has been a global emerging trend towards the utility of medicinal and dietary plants as therapeutic antioxidants. In fact, an inverse relation between the dietary intake of antioxidant-rich medicinal plants and occurrence of human diseases has been established [
27].
In the current work, the in vitro antioxidant potentials of the extracts were evaluated by using a total of six assays, namely DPPH, ABTS, FRAP, CUPRAC, metal chelating, and phosphomolybdenum assays (
Table 3). Interestingly, the methanolic and water extracts of the leaves and stem bark displayed significant radical scavenging potential (DPPH: 95.26–491.59 mg/g; ABTS: 118.34–804.22 mg/g), in comparison to the ethyl acetate extracts of stem bark and leaves, which showed relatively low radical scavenging activity (DPPH: 31.68 ± 0.46; 10.71 ± 0.78 mg/g and ABTS: 22.98 ± 0.93; 2.97 ± 0.66 mg/g, respectively).
Remarkably, the same trend could be observed in CUPRAC and FRAP assays, whereby methanolic extracts of stem bark and leaves were significantly potent as reducing agents (CUPRAC: 1066.93 ± 12.02 and 395.81 ± 12.02 mg/g, respectively; FRAP: 633.44 ± 10.13 and 256.72 ± 3.39 mg/g, respectively), and then subsequently the water extracts of leaves and stem bark (136.26–328.08 mg/g), while the ethyl acetate extracts demonstrated comparatively lower reducing power (30.97–81.54 mg/g). Likewise, the methanolic and water extracts of stem bark demonstrated the highest total antioxidant capacity in the phosphomolybdenum assay (7.11 ± 0.39 and 6.92 ± 0.20 mmol/g, respectively). However, only moderate total antioxidant capacity was noted by the other extracts (2.03–3.17 mmol/g). Unlike in the other antioxidant assays conducted herein, the ethyl acetate extracts were exceptionally found to display the highest activity in the metal chelating assay (stem bark: 26.59 ± 0.25 and leaves: 19.57 ± 0.50 mg/g) relative to the other extracts (8.70–17.83 mg/g) (
Table 3).
Other studies such as the one conducted by Olaide et al. [
28] also assessed the in vitro antioxidant potentials of different extracts of stem bark of
B. ferruginea. In contrast to the present findings, the aqueous extract in that study was found to be a better radical scavenger, showing a significantly higher percent inhibition, as opposed to that of ethanol and ethyl acetate extracts in the DPPH method. Additionally, the aqueous extract was better in Fe
2+ chelating activity and contained higher total phenol than ethanolic and ethyl acetate extracts. The extracts were found to possess hydroxyl radical and nitric oxide scavenging activities as well [
28]. Furthermore, substantial antioxidant activity was revealed for the aqueous extract of
B. ferruginea bark, using six standard tests including ferric thiocyanate lipidic peroxidation inhibition and thiobarbituric acid reacting substances, reducing power, chelating power, and FRAP and DPPH reduction methods [
29].
Interestingly, the studied extracts, particularly the methanolic and water extracts, were observed to be rich in catechin and their derivatives (
Table 2). In fact, it has been reported that the number of hydroxyl groups and the presence of characteristic structural groups have a major impact on the antioxidant activity of catechins. Additionally, catechins exhibit the strong property of neutralizing reactive oxygen and nitrogen species [
30]. Hence, the good antioxidant potential of extracts obtained herein could be related to the presence of these compounds. According to the cholinergic hypothesis, memory impairment induced in Alzheimer’s disease takes place because of a deficiency of cholinergic function in the brain, therefore reducing cortical and hippocampal amounts of the neurotransmitter acetylcholine (ACh) and related enzyme choline transferase. Consequently, inhibition of cholinesterases is regarded as a valuable therapeutic approach for treating symptoms in patients with Alzheimer’s disease [
31]. Indeed, plants are highly rich sources of biologically active compounds, offering huge scope to modern pharmaceutical industry for drug designing. In fact, many synthetic drugs owe their origin to plant-based complementary medicine. Likewise, new treatment strategies based on medicinal plants have been established for diseases like Alzheimer’s disease, being among the most frequent cause of mortality globally [
32,
33].
In the present work, the methanolic extracts of the stem bark of
B. ferruginea showed the highest anti-cholinesterase activity, followed by the methanolic leaf extracts (AChE: 5.18 ± 0.04 and 4.64 ± 0.08; BChE: 12.79 ± 0.93 and 9.27 ± 1.08, respectively) (
Table 4). Nonetheless, relatively good to moderate cholinesterase inhibition was obtained by the other extracts (AChE: 2.39–4.37 mg/g; BChE: 3.15–7.70 mg/g) (
Table 4). Other
Bridelia species such as
B. speciosa have also been reported to act as cholinesterases’ inhibitors [
34].
Disorder in melanin formation has been found to cause a range of skin diseases in humans, for example hyperpigmentation, vitiligo, lentigo, and skin cancer. Additionally, the appearance of brown pigments in vegetables and fruits caused by tyrosinase activity is a leading reason for postharvest losses [
35]. Tyrosinase is a multi-copper enzyme widespread in different organisms that plays an essential role in melanogenesis and enzymatic browning. Accordingly, the discovery, isolation, synthesis, and characterization of new effective tyrosinase inhibitors have been a major focus of research for different applications in food, pharmaceutical, and cosmetics industries [
36].
In the present work, all the extracts were observed to inhibit tyrosinase (46.04–157.07 mg/g). In particular, the methanolic extracts could be demarcated as very potent tyrosinase inhibitors (stem bark: 157.07 ± 0.37 mg/g and leaves: 150.71 ± 0.57 mg/g), followed by water extract of stem bark and the ethyl acetate extracts (103.13–123.23 mg/g). In contrast, water extract of leaves showed the least anti-tyrosinase effect (46.04 ± 3.47 mg/g) (
Table 4).
Indeed, a particularly remarkable and delicate relationship exists between melanogenesis and antioxidant defense systems, linked to scavenging of reactive oxygen species. The synergistic effect in this relationship enhances the efficiency of antioxidants in their ability to scavenge free radicals while tyrosinase inhibitors work and in consequence decreases melanin synthesis [
37]. Additionally, tyrosinase inhibitory activity might rely on the hydroxyl groups of the phenolic compounds in the extracts that could form a hydrogen bond to a site of the enzyme, thereby resulting in reduced enzymatic action. On the other hand, some tyrosinase inhibitors are able to act via hydroxyl groups that bind to the active site of the enzyme, causing steric hindrance or altered conformation. However, antioxidant capacity may be one of the main reasons contributing to tyrosinase inhibition [
38]. Hence, the particularly high anti-tyrosinase effect of methanolic extracts obtained in the present study could be partly due to the relatively good total phenolic content and most likely to the high antioxidant potentials demonstrated in most antioxidant assays.
The inhibition of carbohydrate-hydrolyzing enzymes such as α-amylase and α-glucosidase represents undeniably one of the important therapeutic strategies considered in the treatment of type II diabetes mellitus [
39]. Hyperglycemia, which is one of the major characteristics of type II diabetes, is regarded as the principal reason for complications. A number of pharmacological interventions have been employed to enhance treatment of diabetes, using different modes of action, notably by stimulating insulin release, inhibiting gluconeogenesis, increasing the number of glucose transporters, as well as diminishing absorption of glucose from the intestine, which can be achieved with enzyme inhibitors like acarbose, miglitol, and voglibose [
40]. However, gastrointestinal adverse effects render these drugs less appealing as therapeutic agents, thus making researchers consider natural therapies as viable substitutes [
41], having made their mark throughout history. In fact, earlier studies have reported a number of medicinal plants and their derived compounds to possess the capability to retard enzymes involved in carbohydrate metabolism [
42,
43,
44].
While the studied ethyl acetate extracts displayed dual inhibition on the carbohydrate hydrolyzing enzymes (amylase: 0.94 ± 0.01 and 0.91 ± 0.04 mmol/g; glucosidase: 6.68 ± 0.01 and 6.24 ± 0.29 mmol/g), the other extracts selectively inhibited amylase (0.21–1.38 mmol/g). The methanolic extracts especially, showed the highest anti-amylase effect (1.38 ± 0.03 and 1.35 ± 0.05 mmol/g), followed by the water extract of stem bark (1.06 ± 0.04 mmol/g) (
Table 4).
Other reports have also studied the inhibitory effects of
B. ferruginea extracts on these enzymes. For instance, in the study of Bakoma et al. [
45], the ethyl acetate fraction of
B. ferruginea, possessing high phenolic contents and antioxidant potential, was shown to exhibit higher inhibition activities on α-glucosidase (IC
50 = 0.19 mg/mL) and α-amylase (IC
50 = 0.24 mg/mL) in contrast to acetone fraction and hydroalcoholic extract having lower phenolic content with low inhibition activity (IC
50 > 0.25 mg/mL), suggesting the phenolic compounds are likely the source of inhibition against both enzymes. Additionally, Kwon et al. [
46] reported phenolic-enriched extract to display high
α-glucosidase inhibition along with
α-amylase inhibitory property. Additionally, Bhandari et al. [
47] and Shanmugam et al. [
48] evaluated the inhibitory activities of phenolic compounds against α-glucosidase and α-amylase, whereby they were found to inhibit both enzymes significantly. Moreover, in agreement with the present findings, the ethyl acetate fraction of the
B. ferruginea stem bark in the study of Ojo et al. [
49] showed
α-glucosidase (IC
50: 4.52 ± 0.50 mg/mL) and
α-amylase (IC
50: 5.42 ± 1.10 mg/mL) inhibitory activities. Additionally, enzymes kinetics studies pointed out that the ethyl acetate fraction acted as a non-competitive inhibitor and a competitive inhibitor for glucosidase and amylase, respectively. The presence of phenols such as 2,2′ oxydiphenol and p-hydroxyphenyl ether were suggested to be the main bioactive compounds accountable for the enzyme inhibition activities [
49]. In furtherance, in a study by Mahomoodally et al. [
34], another species of the
Bridelia genus,
Bridelia speciosa Müll. Arg. stem bark extracts, were investigated. Similarly, only the ethyl acetate extract showed inhibition against glucosidase, which was in accordance with the present findings.
Considering the potential cytotoxicity of natural compounds extracted from
B. ferruginea [
11] and the intrinsic antimicrobial activity displayed by herbal extracts rich in total phenolic compounds [
50,
51], the present study also attempted to inspect the anti-proliferative and antimicrobial effects of
B. ferruginea. The anti-proliferative properties were evaluated against the human colon cancer HCT116 cell line. While antimicrobial effects were assayed against multiple Gram− (
E. coli,
P. aeruginosa,
S. tiphy), Gram+ (
B. subtilis,
B. cereus, and
S. aureus), and fungi (
C. albicans, C. tropicalis and
C. parapsilosis) strains, deeply involved in colon inflammation [
37,
52,
53,
54]. Antimycotic properties were also evaluated against different dermatophytes strains (
T. mentagrophytes,
T. tonsurans, T. rubrum, A. quadrifidum,
N. gypseum,
A. currei, and
A. insingulare). Specifically, the bio-pharmacological evaluations focused on the methanolic extract from stem bark that displayed the highest scavenging/reducing and enzyme inhibition properties. Initially, the biocompatibility limit of the extract was determined, in the concentration range of 0.1–20 mg/mL, through the brine shrimp
A. salina lethality test, an eco-toxicological assay regarded to be, at least partly, predictive of cytotoxicity [
55]. The LC
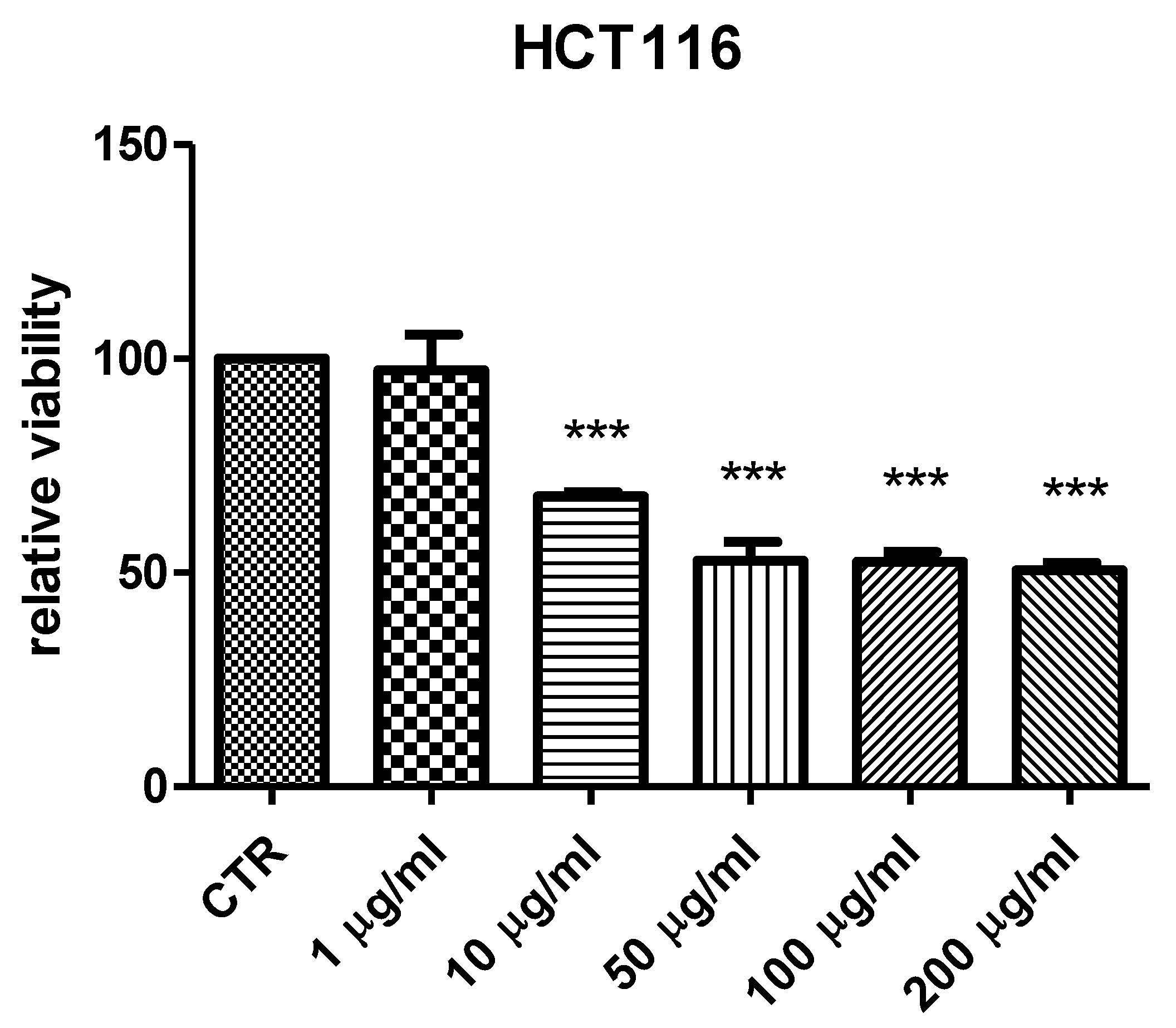
50 value resulting from brine shrimp test was <2 mg/mL. Therefore, at least ten-fold lower concentration was employed for the subsequent in vitro tests. Specifically, a concentration-dependent (1–200 µg/mL) inhibition of HCT116 cell viability was observed (
Figure 1), thus demonstrating an anti-proliferative effect that occurs within the range of biocompatibility yielded by the brine shrimp test. Recently, polar extracts from
B. speciosa were found to reduce the viability of liver cancer cells (HepG2) [
34]. The inhibition of cancer cell viability was related, at least in part, to the catechin fraction measured in the extracts. In analogy, it can be hypothesized that the current inhibitory effect induced by
B. ferruginea stem bark methanolic extract on HCT116 cell viability could depend, albeit partially, on the numerous catechin compounds detected by HPLC-MS qualitative analysis. This hypothesis is also substantiated by literature data highlighting the inhibitory effects of HCT116 cell viability induced by epigallocatechin-3-gallate, possibly mediated by the inhibition of MET receptor tyrosine kinase and DNA methyltransferases [
56,
57]. Catechin levels have also been related to the anti-proliferative effects induced by anti-inflammatory
Epilobium angustifolium and
Phyllantus niruri water extracts, in prostate cancer PC3 cells [
50]. However, the bioinformatics prediction based on the chemical composition of
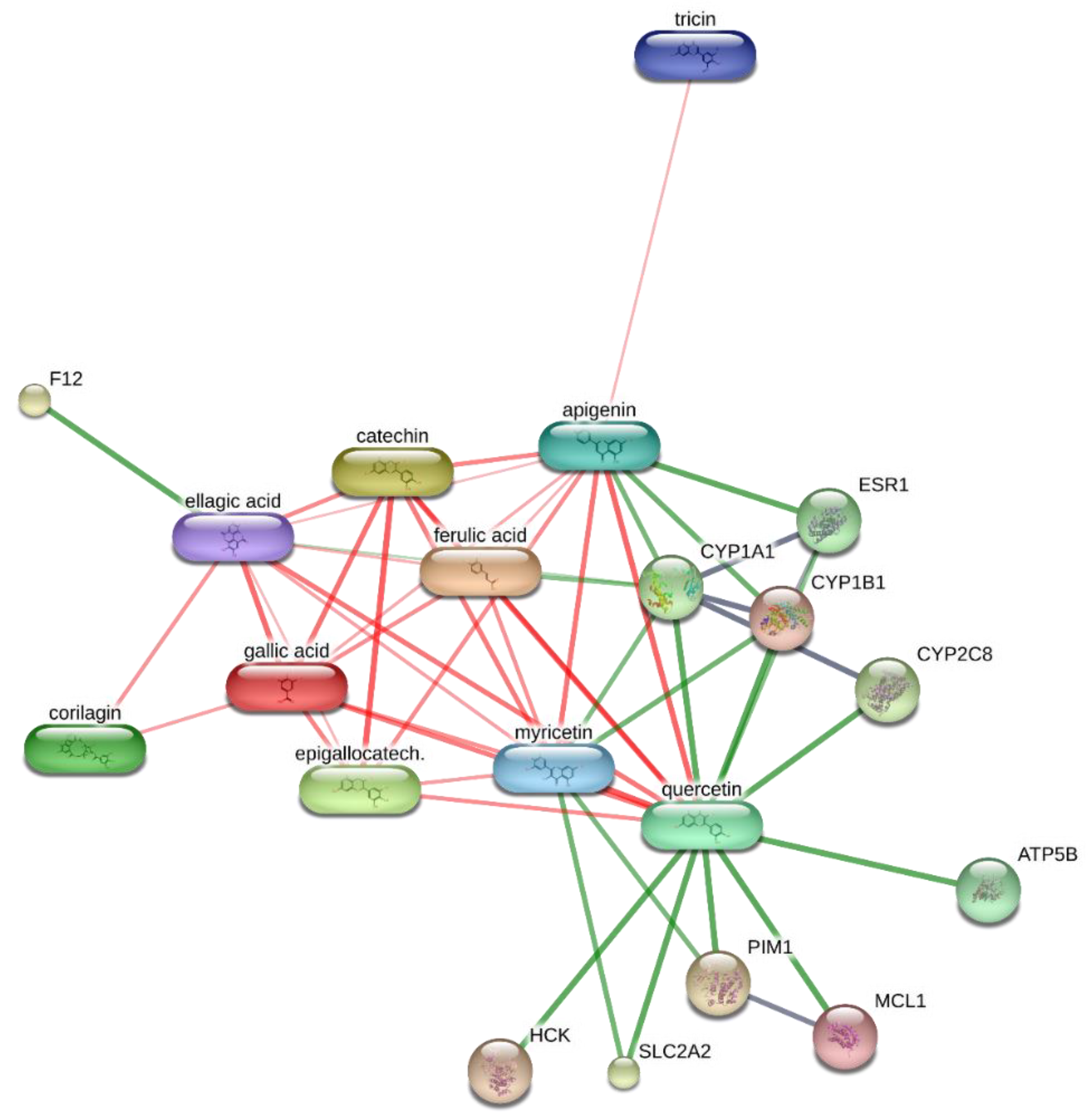
B. ferruginea methanolic extract, conducted via the STITCH platform (
http://stitch.embl.de/ accessed on 15 February 2021), pointed to the prominent position of quercetin in the scenario of the components–targets analysis (
Figure 2). In particular, quercetin was predicted to interact with pim-1 oncogene (PIM-1), a proto-oncogene involved in cell survival and proliferation [
58], whose expression was also observed in HCT116 cells [
59]. Overall, the past and present studies suggest the polar extracts from
Bridelia species as good candidates for future studies aimed to explore anticancer activity, in vivo. Regarding the antibacterial activity, the methanolic extract was effective against all the tested strains, except
P. aeruginosa. Intriguingly, the bacteriostatic effects induced by the extract occurred at concentrations (2.48–125 µg/mL) somewhat lower than the biocompatibility limit calculated by the brine shrimp test and in the concentration range effective in inducing the anti-proliferative effect on HCT116 cells (
Table 5). This suggests that the tested extract could also be a promising agent for counteracting dysbiosis-related inflammatory disorders in the colon. Among the assayed strains,
E. coli was the most sensitive to the bacterial growth inhibition induced by the extract that was also effective against all tested fungi and yeasts strains. Among the fungi, dermatophytes species showed higher sensitivity to the growth inhibitory effects induced by the extract (
Table 6). The inhibition of dermatophytes growth adds to the highest anti-tyrosinase effect (
Table 4) displayed by this extract, thus further supporting potential uses against skin hyperpigmentation, which could be also stimulated by dermatophytes infections [
60,
61]. Currently, the observed antimicrobial effects are consistent, albeit in part, with the presence of phenols and flavonoids [
62,
63]. Interestingly, the antibacterial mechanism and in particular, membrane disruption, in both Gram-positive and Gram-negative bacteria, that contributes to the antibacterial activity of most plant phenolics has been widely assessed, as reviewed by Rempe et al. [
64]. The present HPLC-MS fingerprint analysis and recent published data suggest that catechin fraction could play a pivotal role in the observed antibacterial effects, especially against
E. coli [
50,
65]. However, the bioinformatics prediction conducted to investigate the putative interactions of extracts’ phytocompounds against the dermatophytes species
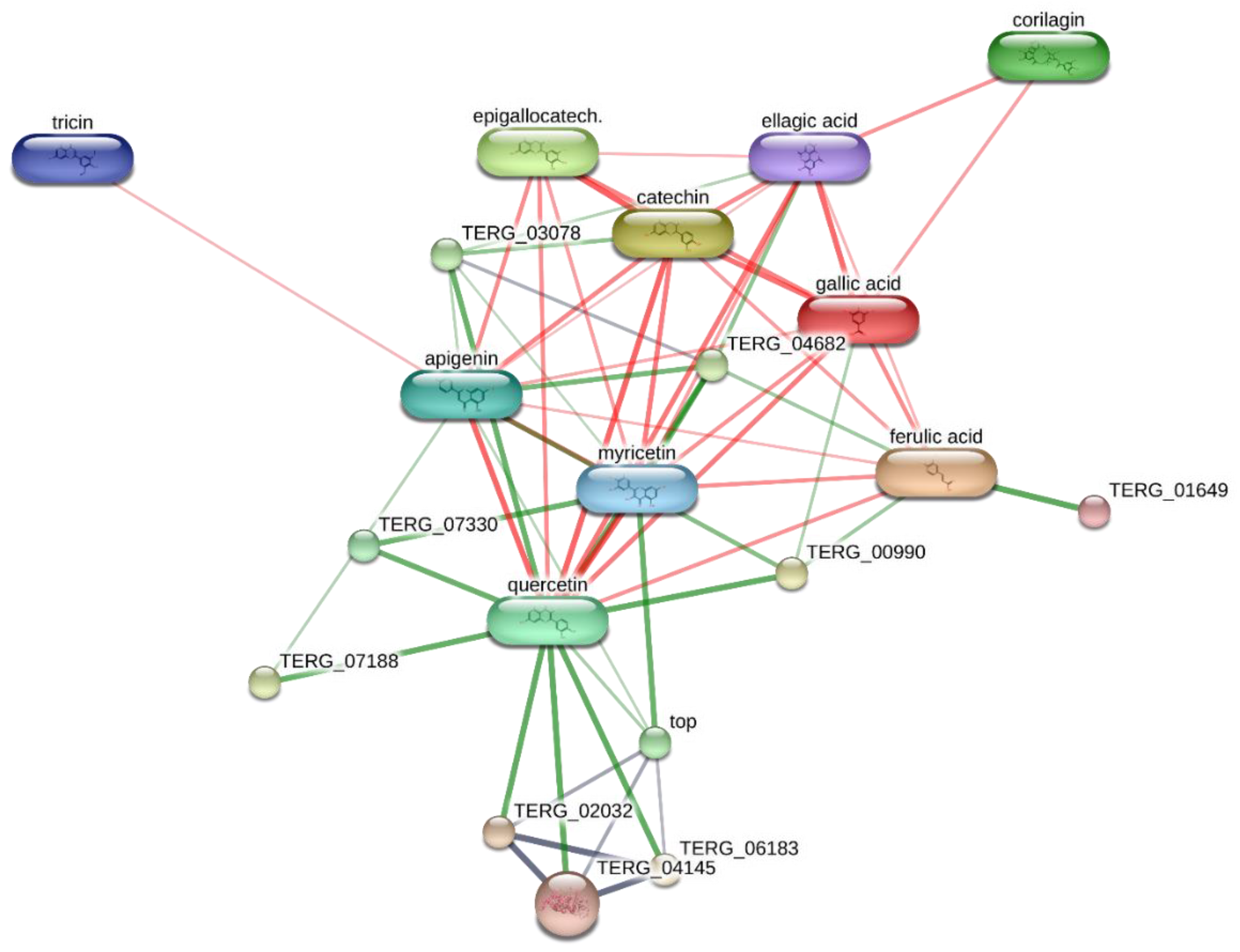
T. rubrum, chosen for its availability on the bioinformatics platform STITCH, suggested the interactions of epigallocatechin, quercetin, and gallic acid with multiple dermatophytes enzymes, including ATP synthase (TERG_07188), UDP-glucose:sterol glucosyltransferase (TERG_00990) and xanthine dehydrogenases (TERG_06183, TERG_04145, TERG02032) (
Figure 2 and
Figure 3), involved in the energetic metabolism. Overall, the reported bioinformatics prediction is consistent with the antifungal effects induced by the extract. Nevertheless, future investigations are essential to quantify the identified compounds in the extract and to confirm the present findings with independent biological models.



 ,
, 







{kind=link}
{kind=link}
{kind=link}