
Antibiotic Resistance in Shiga Toxigenic Escherichia coli Isolates from Surface Waters and Sediments in a Mixed Use Urban Agricultural Landscape

 , ,
, ,
Abstract
:1. Introduction
2. Results
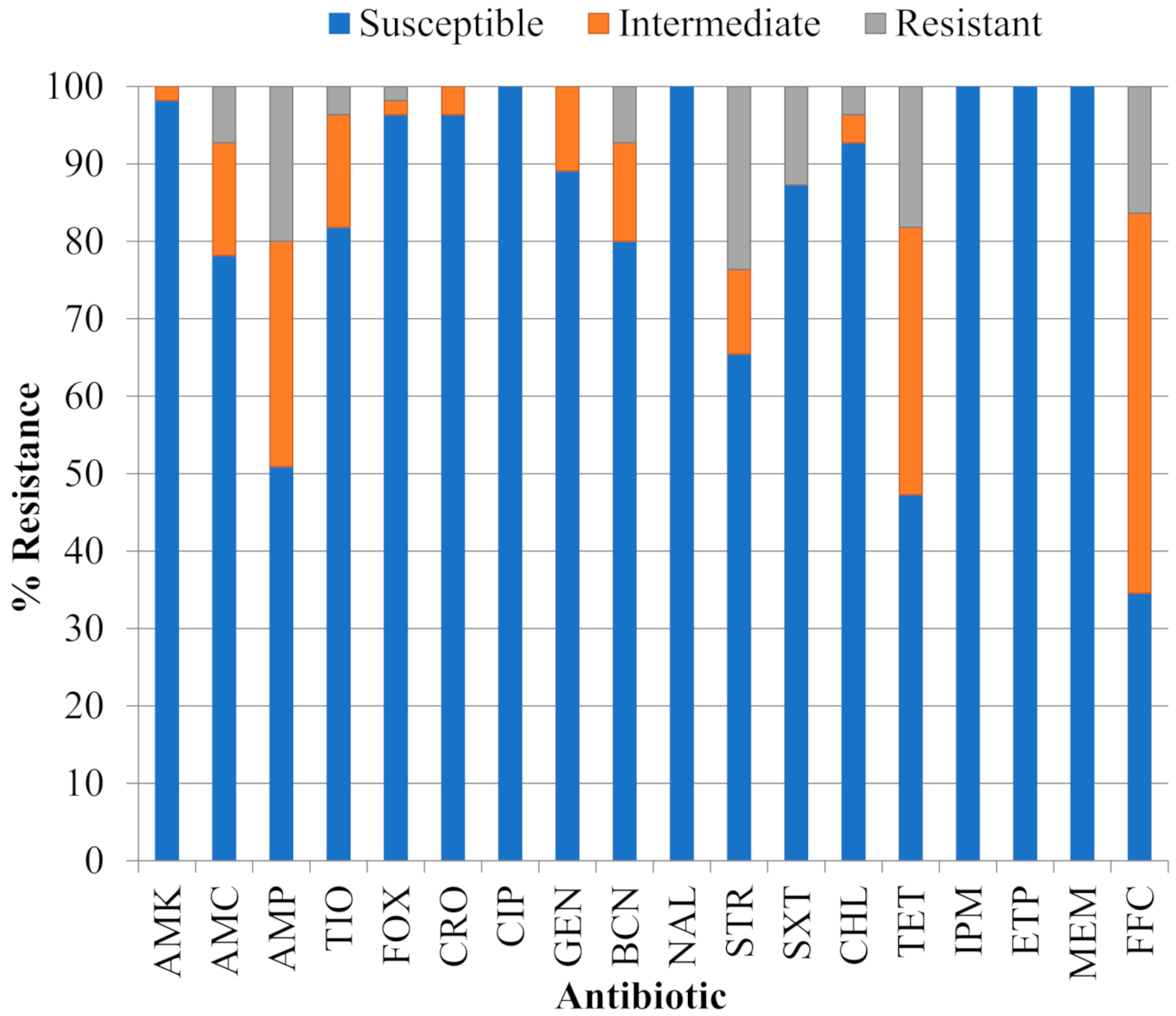
2.1. Antibiotic Resistance Phenotypes of STEC Isolates Recovered from Surface Water and Sediments
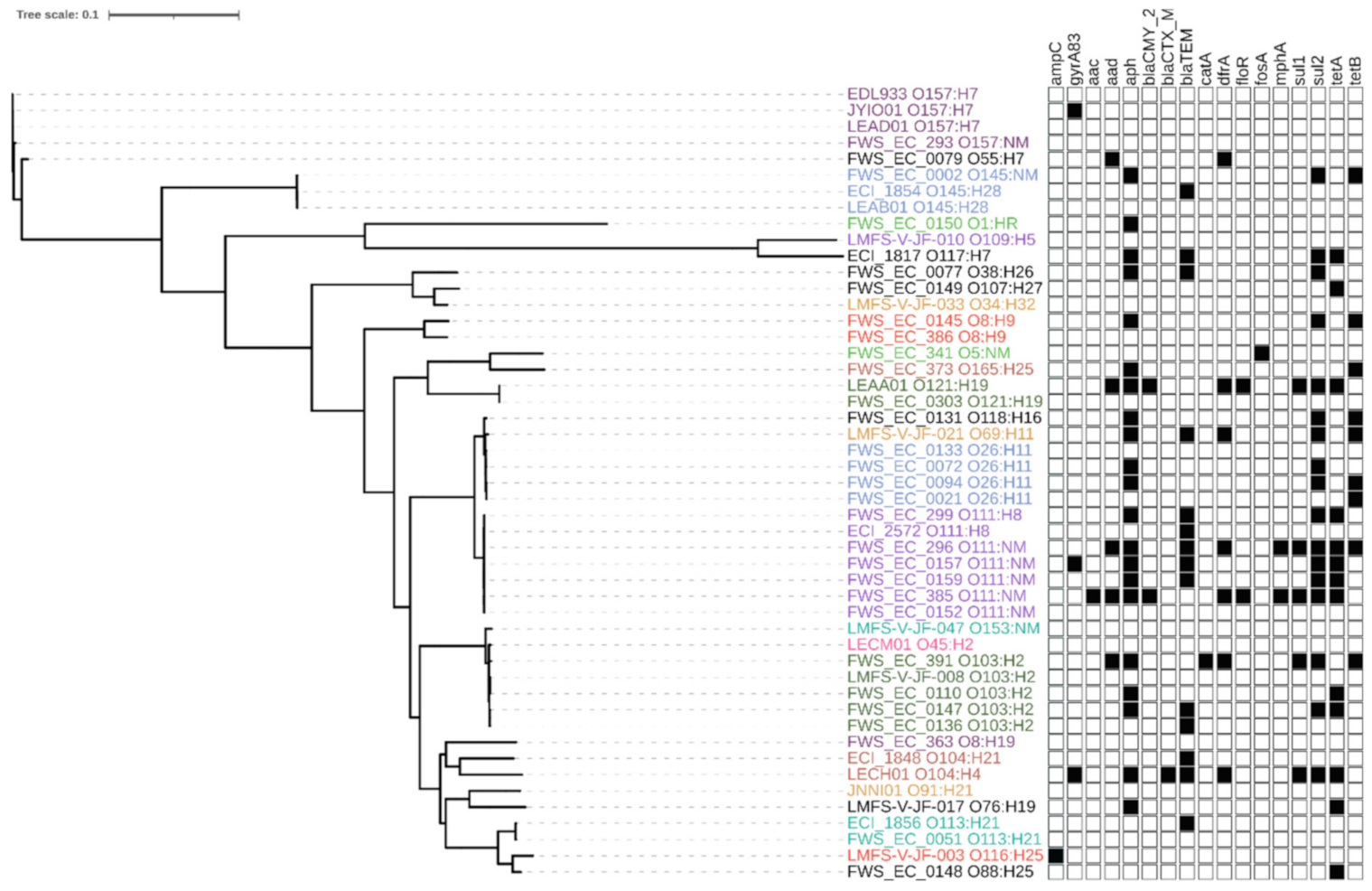
2.2. Genomic Analysis of the STEC Isolates
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Antibiotic Susceptibility Testing
4.3. Genomic Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riley, L.W.; Remis, R.S.; Helgerson, S.D.; McGee, H.B.; Wells, J.G.; Davis, B.R.; Hebert, R.J.; Olcott, E.S.; Johnson, L.M.; Hargrett, N.T.; et al. hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med. 1983, 308, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar] [CrossRef]
- Nathanson, S.; Kwon, T.; Elmaleh, M.; Charbit, M.; Launay, E.A.; Harambat, J.; Brun, M.; Ranchin, B.; Bandin, F.; Cloarec, S.; et al. Acute neurological involvement in diarrhea-associated hemolytic uremic syndrome. Clin. J. Am. Soc. Nephrol. 2010, 5, 1218–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melton-Celsa, A.; Mohawk, K.; Teel, L.; O’Brien, A. Pathogenesis of Shiga-Toxin Producing Escherichia coli; Springer: Berlin/Heidelberg, Germany, 2011; Volume 357, pp. 67–103. [Google Scholar]
- Pires, S.M.; Majowicz, S.; Gill, A.; Devleesschauwer, B. Global and regional source attribution of Shiga toxin-producing Escherichia coli infections using analysis of outbreak surveillance data. Epidemiol. Infect. 2019, 147, e236. [Google Scholar] [CrossRef] [Green Version]
- Goldwater, P.N.; Bettelheim, K.A. Treatment of enterohemorrhagic Escherichia coli (EHEC) infection and hemolytic uremic syndrome (HUS). BMC Med. 2012, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; McDaniel, A.D.; Wolf, L.E.; Keusch, G.T.; Waldor, M.K.; Acheson, D.W.K. Quinolone antibiotics induce shiga toxin–encoding bacteriophages, toxin production, and death in mice. J. Infect. Dis. 2000, 181, 664–670. [Google Scholar] [CrossRef]
- Agger, M.; Scheutz, F.; Villumsen, S.; Mølbak, K.; Petersen, A.M. Antibiotic treatment of verocytotoxin-producing Escherichia coli (VTEC) infection: A systematic review and a proposal. J. Antimicrob. Chemother. 2015, 70, 2440–2446. [Google Scholar] [CrossRef] [Green Version]
- Menne, J.; Nitschke, M.; Stingele, R.; Abu-Tair, M.; Beneke, J.; Bramstedt, J.; Bremer, J.P.; Brunkhorst, R.; Busch, V.; Dengler, R.; et al. Validation of treatment strategies for enterohaemorrhagic Escherichia coli O104:H4 induced haemolytic uraemic syndrome: Case-control study. BMJ 2012, 345, e4565. [Google Scholar] [CrossRef] [Green Version]
- Wieler, L.H.; Bauerfeind, R. STEC as a veterinary problem. Diagnostics and prophylaxis in animals. Methods Mol. Med. 2003, 73, 75–89. [Google Scholar] [PubMed]
- Premanandh, J.; Samara, B.S.; Mazen, A.N. Race against antimicrobial resistance requires coordinated action—An overview. Front. Microbiol. 2016, 6, 1536. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance—The need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Antimicrobial Resistance Global Report on Surveillance 2014. Available online: http://www.who.int/drugresistance/documents/surveillancereport/en (accessed on 18 January 2021).
- Kim, H.H.; Samadpour, M.; Grimm, L.; Clausen, C.R.; Besser, T.E.; Baylor, M.; Kobayashi, J.M.; Neill, M.A.; Schoenknecht, F.D.; Tarr, P.I. Characteristics of antibiotic-resistant Escherichia coli O157:H7 in Washington State, 1984–1991. J. Infect. Dis. 1994, 170, 1606–1609. [Google Scholar] [CrossRef]
- Zhao, S.; White, D.G.; Ge, B.; Ayers, S.; Friedman, S.; English, L.; Wagner, D.; Gaines, S.; Meng, J. Identification and characterization of integron-mediated antibiotic resistance among Shiga toxin-producing Escherichia coli isolates. Appl. Environ. Microbiol. 2001, 67, 1558–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Schroeder, C.M.; Meng, J.; White, D.G.; McDermott, P.F.; Wagner, D.D.; Yang, H.; Simjee, S.; Debroy, C.; Walker, R.D.; et al. Identification of antimicrobial resistance and class 1 integrons in Shiga toxin-producing Escherichia coli recovered from humans and food animals. J. Antimicrob. Chemother. 2005, 56, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Murinda, S.E.; Ebner, P.D.; Nguyen, L.T.; Mathew, A.G.; Oliver, S.P. Antimicrobial resistance and class 1 integrons in pathogenic Escherichia coli from dairy farms. Foodborne Pathog. Dis. 2005, 2, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Junker, E.; Schroeter, A.; Helmuth, R.; Guth, B.E.C.; Beutin, L. Phenotypic and genotypic characterization of antimicrobial resistance in Escherichia coli O111 isolates. J. Antimicrob. Chemother. 2006, 57, 1210–1214. [Google Scholar] [CrossRef] [Green Version]
- Karmali, M.A.; Gannon, V.; Sargeant, J.M. Verocytotoxin-producing Escherichia coli (VTEC). Vet. Microbiol. 2010, 140, 360–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Stokes, H.W.; Hall, R.M. A novel family of potentially mobile DNA elements encoding site-specific gene-integration functions: Integrons. Mol. Microbiol. 1989, 3, 1669–1683. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shintani, M.; Sanchez, Z.K.; Kimbara, K. Genomics of microbial plasmids: Classification and identification based on replication and transfer systems and host taxonomy. Front. Microbiol. 2015, 6, 242. [Google Scholar] [CrossRef] [PubMed]
- Losada, L.; Debroy, C.; Radune, D.; Kim, M.; Sanka, R.; Brinkac, L.M.; Kariyawasam, S.; Shelton, D.R.; Fratamico, P.M.; Kapur, V.; et al. Whole genome sequencing of diverse Shiga toxin-producing and non-producing Escherichia coli strains reveals a variety of virulence and novel antibiotic resistance plasmids. Plasmid 2016, 83, 8–11. [Google Scholar] [CrossRef]
- Köser, C.U.; Ellington, M.J.; Peacock, S.J. Whole-genome sequencing to control antimicrobial resistance. Trends Genet. 2014, 30, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Tyson, G.H.; McDermott, P.F.; Li, C.; Chen, Y.; Tadesse, D.A.; Mukherjee, S.; Bodeis-Jones, S.; Kabera, C.; Gaines, S.A.; Loneragan, G.H.; et al. WGS accurately predicts antimicrobial resistance in Escherichia coli. J. Antimicrob. Chemother. 2015, 70, 2763–2769. [Google Scholar] [CrossRef] [Green Version]
- Nadya, S.; Delaquis, P.; Chen, J.; Allen, K.; Johnson, R.P.; Ziebell, K.; Laing, C.; Gannon, V.; Bach, S.; Topp, E. Phenotypic and genotypic characteristics of Shiga toxin-producing Escherichia coli isolated from surface waters and sediments in a Canadian urban-agricultural landscape. Front. Cell. Infect. Microbiol. 2016, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.S. Prevalence and pathogenicity of Shiga toxin-producing Escherichia coli in beef cattle and their products. J. Anim. Sci. 2007, 85, E63–E72. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Hu, B.; Xu, Y.; Sun, H.; Zhao, A.; Ba, P.; Fu, S.; Fan, R.; Jin, Y.; Wang, H.; et al. Molecular and phylogenetic characterization of non-O157 Shiga toxin-producing Escherichia coli strains in China. Front. Cell. Infect. Microbiol. 2016, 6, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Fratamico, P.; Debroy, C.; Bumbaugh, A.C.; Allen, J.W. DNA Sequencing and identification of serogroup-specific genes in the Escherichia coli O118 O antigen gene cluster and demonstration of antigenic diversity but only minor variation in DNA sequence of the O antigen clusters of E. coli O118 and O151. Foodborne Pathog. Dis. 2008, 5, 449–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Day, M.J.; Mafura, M.T.; Nunez-Garcia, J.; Fenner, J.J.; Sharma, M.; Van Essen-Zandbergen, A.; Rodríguez, I.; Dierikx, C.; Kadlec, K.; et al. Comparative analysis of ESBL-positive Escherichia coli isolates from animals and humans from the UK, The Netherlands and Germany. PLoS ONE 2013, 8, e75392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Nolan, L.K. Pathogenomics of the Virulence Plasmids of Escherichia coli. Microbiol. Mol. Biol. Rev. 2009, 73, 750–774. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Partridge, S.R.; Yang, X.; Hou, J.; Deng, Y.; Yao, Q.; Zeng, Z.; Chen, Z.; Liu, J. Complete nucleotide sequence of pHN7A8, an F33:A-:B- type epidemic plasmid carrying blaCTX-M-65, fosA3 and rmtB from China. J. Antimicrob. Chemother. 2013, 68, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Bielak, E.M.; Fortini, D.; Hansen, L.H.; Hasman, H.; Debroy, C.; Nolan, L.K.; Carattoli, A. Expansion of the IncX plasmid family for improved identification and typing of novel plasmids in drug-resistant Enterobacteriaceae. Plasmid 2012, 68, 43–50. [Google Scholar] [CrossRef]
- Allen, K.J.; Laing, C.R.; Cancarevic, A.; Zhang, Y.; Mesak, L.R.; Xu, H.; Paccagnella, A.; Gannon, V.P.J.; Hoang, L. Characteristics of clinical Shiga toxin-producing Escherichia coli Isolated from British Columbia. BioMed Res. Int. 2013, 2013, 1–11. [Google Scholar] [CrossRef]
- Health Canada. Categorization of Antimicrobial Drugs Based on Importance in Human Medicine. Available online: https://www.canada.ca/en/health-canada/services/drugs-health-products/veterinary-drugs/antimicrobial-resistance/categorization-antimicrobial-drugs-based-importance-human-medicine.html (accessed on 18 January 2021).
- Maal-Bared, R.; Bartlett, K.H.; Bowie, W.R.; Hall, E.R. Phenotypic antibiotic resistance of Escherichia coli and E. coli O157 isolated from water, sediment and biofilms in an agricultural watershed in British Columbia. Sci. Total. Environ. 2013, 443, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhao, S.; Doyle, M.P.; Joseph, S.W. Antibiotic resistance of Escherichia coli O157:H7 and O157:NM isolated from animals, food, and humans. J. Food Prot. 1998, 61, 1511–1514. [Google Scholar] [CrossRef]
- Cabal, A.; Gómez-Barrero, S.; Porrero, C.; Bárcena, C.; López, G.; Cantón, R.; Gortazar, C.; Domínguez, L.; Alvarez, J. Assessment of virulence factors characteristic of human Escherichia coli pathotypes and antimicrobial resistance in O157:H7 and non-O157:H7 isolates from livestock in Spain. Appl. Environ. Microbiol. 2013, 79, 4170–4172. [Google Scholar] [CrossRef] [Green Version]
- Krüger, A.; Lucchesi, P.M.A.; Sanso, A.M.; Etcheverría, A.I.; Bustamante, A.V.; Burgán, J.; Fernández, L.; Fernandez, D.; Leotta, G.; Friedrich, A.W.; et al. Genetic characterization of Shiga toxin-producing Escherichia coli O26:H11 strains isolated from animal, food, and clinical samples. Front. Cell. Infect. Microbiol. 2015, 5, 74. [Google Scholar] [CrossRef] [Green Version]
- Amézquita-López, B.A.; Quiñones, B.; Soto-Beltrán, M.; Lee, B.G.; Yambao, J.C.; Lugo-Melchor, O.Y.; Chaidez, C. Antimicrobial resistance profiles of Shiga toxin-producing Escherichia coli O157 and Non-O157 recovered from domestic farm animals in rural communities in Northwestern Mexico. Antimicrob. Resist. Infect. Control. 2016, 5, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, M.; Doumith, M.; Jenkins, C.; Dallman, T.J.; Hopkins, K.L.; Elson, R.; Godbole, G.; Woodford, N. Antimicrobial resistance in Shiga toxin-producing Escherichia coli serogroups O157 and O26 isolated from human cases of diarrhoeal disease in England, 2015. J. Antimicrob. Chemother. 2017, 72, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, A.; Blanco, J.E.; Blanco, M.; Alonso, M.P.; Dhabi, G.; Echeita, A.; González, E.A.; Bernárdez, M.I.; Blanco, J. Antimicrobial resistance of Shiga toxin (verotoxin)-producing Escherichia coli O157:H7 and non-O157 strains isolated from humans, cattle, sheep and food in Spain. Res. Microbiol. 2005, 156, 793–806. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). National Antimicrobial Resistance Monitoring System for Enteric Bacteria. Available online: https://www.cdc.gov/narms/index.html (accessed on 18 January 2021).
- British Columbia Center of Disease Control (BCCDC). Antimicrobial Resistance Trends in the Province of British Columbia. 2014. Available online: http://www.bccdc.ca/resource-gallery/Documents/AMR%202014%20Report-3NOV2015_FINAL.pdf (accessed on 18 January 2021).
- Redgrave, L.S.; Sutton, S.B.; Webber, M.A.; Piddock, L.J. Fluoroquinolone resistance: Mechanisms, impact on bacteria, and role in evolutionary success. Trends Microbiol. 2014, 22, 438–445. [Google Scholar] [CrossRef]
- Fasugba, O.; Gardner, A.; Mitchell, B.G.; Mnatzaganian, G. Ciprofloxacin resistance in community- and hospital-acquired Escherichia coli urinary tract infections: A systematic review and meta-analysis of observational studies. BMC Infect. Dis. 2015, 15, 545. [Google Scholar] [CrossRef] [Green Version]
- Public Health Agency of Canada (PHAC). Human Antimicrobial Use Report. 2014. Available online: https://www.canada.ca/en/public-health/services/publications/drugs-health-products/human-antimicrobial-use-report-2014.html (accessed on 1 February 2021).
- Public Health Agency of Canada (PHAC). Antimicrobial Resistance Surveillance System (CARSS) Report. 2016. Available online: https://www.canada.ca/en/public-health/services/publications/drugs-health-products/canadian-antimicrobial-resistance-surveillance-system-report-2016.html (accessed on 1 February 2021).
- Morrison, D.B.; Saksida, S. Trends in antimicrobial use in Marine Harvest Canada farmed salmon production in British Columbia (2003–2011). Can. Vet. J. 2013, 54, 1160–1163. [Google Scholar] [PubMed]
- Gomi, R.; Matsuda, T.; Matsumura, Y.; Yamamoto, M.; Tanaka, M.; Ichiyama, S.; Yoneda, M. Whole-genome analysis of antimicrobial-resistant and extraintestinal pathogenic Escherichia coli in river water. Appl. Environ. Microbiol. 2017, 83, e02703-16. [Google Scholar] [CrossRef] [Green Version]
- Hamelin, K.; Bruant, G.; El-Shaarawi, A.; Hill, S.; Edge, T.A.; Fairbrother, J.; Harel, J.; Maynard, C.; Masson, L.; Brousseau, R. Occurrence of virulence and antimicrobial resistance genes in Escherichia coli isolates from different aquatic ecosystems within the St. Clair River and Detroit River Areas. Appl. Environ. Microbiol. 2006, 73, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stange, C.; Sidhu, J.; Tiehm, A.; Toze, S. Antibiotic resistance and virulence genes in coliform water isolates. Int. J. Hyg. Environ. Health 2016, 219, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Von Wintersdorff, C.J.H.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [Green Version]
- van Hoek, A.H.; Mevius, D.; Guerra, B.; Mullany, P.; Roberts, A.P.; Aarts, H.J. Acquired antibiotic resistance genes: An overview. Front. Microbiol. 2011, 2, 1–203. [Google Scholar]
- Poole, K. Efflux-mediated antimicrobial resistance. In Antibiotic Discovery and Development; Dougherty, T.J., Pucci, M.J., Eds.; Springer: Boston, MA, USA, 2012; pp. 349–395. [Google Scholar]
- Jacoby, G.A. AmpC β-lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [Green Version]
- Blickwede, M.; Schwarz, S. Molecular analysis of florfenicol-resistant Escherichia coli isolates from pigs. J. Antimicrob. Chemother. 2004, 53, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Meunier, D.; Jouy, E.; Lazizzera, C.; Doublet, B.; Kobisch, M.; Cloeckaert, A.; Madec, J.-Y. Plasmid-borne florfenicol and ceftiofur resistance encoded by the floR and blaCMY-2 genes in Escherichia coli isolates from diseased cattle in France. J. Med. Microbiol. 2010, 59, 467–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Hansen, K.H.; Nielsen, C.A.; Fritsche, T.R.; Guardabassi, L. High diversity of plasmids harbouring blaCMY-2 among clinical Escherichia coli isolates from humans and companion animals in the upper Midwestern USA. J. Antimicrob. Chemother. 2014, 69, 1492–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falardeau, J.; Johnson, R.P.; Pagotto, F.; Wang, S. Occurrence, characterization, and potential predictors of verotoxigenic Escherichia coli, Listeria monocytogenes, and Salmonella in surface water used for produce irrigation in the Lower Mainland of British Columbia, Canada. PLoS ONE 2017, 12, e0185437. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Tech. Bull. Regist. Med. Technol. 1966, 36, 49. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing: Twenty-First Informational Supplement; CLSI Document M100-S21; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011; ISBN 1-56238-742-1. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 5th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; ISBN 978-1-68440-009-6. [Google Scholar]
- Ahmed, M.O.; Clegg, P.D.; Williams, N.J.; Baptiste, K.E.; Bennett, M. Antimicrobial resistance in equine faecal Escherichia coli isolates from North West England. Ann. Clin. Microbiol. Antimicrob. 2010, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Fong, K.; Lu, Y.T.L.; Brenner, T.; Falardeau, J.; Wang, S. CRISPR and prophage diversity across Salmonella & verotoxin-producing Escherichia coli. BMC Genom. 2021. [Google Scholar]
- Leinonen, R.; Sugawara, H.; Shumway, M.; International Nucleotide Sequence Database Collaboration. The sequence read archive. Nucleic Acids Res. 2011, 39, D19–D21. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.; Clark, K.; Gevorgyan, R.; Gorelenkov, V.; Gribov, E.; Karsch-Mizrachi, I.; Kimelman, M.; Pruitt, K.D.; Resenchuk, S.; Tatusova, T.; et al. BioProject and BioSample databases at NCBI: Facilitating capture and organization of metadata. Nucleic Acids Res. 2012, 40, D57–D63. [Google Scholar] [CrossRef] [PubMed]
- Kislyuk, A.O.; Katz, L.S.; Agrawal, S.; Hagen, M.S.; Conley, A.B.; Jayaraman, P.; Nelakuditi, V.; Humphrey, J.C.; Sammons, S.A.; Govil, D.; et al. A computational genomics pipeline for prokaryotic sequencing projects. Bioinformatics 2010, 26, 1819–1826. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, J.; Noll, I.; Feig, M.; Weiss, B.; Claus, H.; Werner, G.; Eckmanns, T.; Hermes, J.; Abu Sin, M. Decline in the proportion of methicillin resistance among Staphylococcus aureus isolates from non-invasive samples and in outpatient settings, and changes in the co-resistance profiles: An analysis of data collected within the Antimicrobial Resistance Surveillance Network, Germany 2010 to 2015. BMC Infect. Dis. 2017, 17, 169. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Isolate Number and Serotype | Antibiotic Resistance Phenotype 1 | Source |
|---|---|---|
| Pansusceptible | ||
| 347-O174:H21 | None | Water |
| 005-O103:H2 | None | Water |
| 007-O103:H2 | None | Water |
| 008-O103:H2 | None | Water |
| 010-O109:H5 | None | Water |
| 033-O34:H32 | None | Water |
| 036-O34:H32 | None | Water |
| 039-O22:H8 | None | Water |
| 043-O153:NM | None | Water |
| 047-O153:NM | None | Water |
| Intermediate resistant | ||
| 292-O177:NM | TETI FFCI | Water |
| 293-O157:NM | TIOI GENI BCNI STRI TETI FFCI | Water |
| 294-O26:H11 | TETI FFCI | Water |
| 300-O26:H11 | FFCI | Water |
| 301-O26:H11 | FFCI | Water |
| 337-O8:H19 | TETI FFCI | Water |
| 338-O168:H8 | FFCR AMPI TIOI GENI BCNI STRI TETI | Water |
| 340-O116:H25 | AMCR AMPR AMKI TIOI FOXI CROI GENI BCNI STRI TETI | Water |
| 341-O5:NM | AMCI AMPI TETI FFCI | Water |
| 343-O98:NM | TIOI GENI STRI TETI FFCI | Water |
| 344-O5:NM | FFCR AMCI AMPI TETI | Water |
| 345-O22:H8 | FFCR AMCI AMPI TETI | Water |
| 360-OR:H21 | FFCI | Water |
| 362-O5:NM | AMCI FFCI | Water |
| 363-O8:H19 | TETI FFCI | Water |
| 365-O128:H2 | AMPI TETI FFCI | Water |
| 369-O98:NM | STRI CHLI TETI FFCI | Water |
| 370-O98:NM | CHLI TETI FFCI | Water |
| 371-O157:H7 | FFCI | Water |
| 375-O157:H7 | FFCI | Water |
| 377-O103:H2 | FFCI | Water |
| 378-O103:H25 | AMPI TIOI FFCI | Water |
| 379-O26:H11 | FFCI | Water |
| 380-O165:NM | FFCR AMPI TETI | Water |
| 381-O174:H8 | STRR AMPI TIOI GENI BCNI | Water |
| 383-O163:H19 | BCNI STRI TETI FFCI | Water |
| 384-O128:H2 | AMPI TETI | Water |
| 386-O8:H9 | AMPI FFCI | Water |
| 387-O103:H25 | FFCR AMPI | Water |
| 389-O103:H2 | AMPI FFCI | Sediment |
| 390-O103:H2 | AMPI FFCI | Sediment |
| 003-O116:H25 | AMXR, AMPR | Water |
| 012-O116:H25 | AMXR, AMPR | Water |
| 017-O76:H19 | STRR, TETR | Water |
| 021-O69:H11 | AMPR, STRR, SXTR, TETR | Water |
| 025-O69:H11 | AMPR, STRR, SXTR, TETI | Water |
| 029-O69:H11 | AMPR, STRR, SXTR, TETI | Water |
| Multidrug resistant | ||
| 296-O111:NM | AMPR STRR SXTR TETR AMCI FFCI | Water |
| 298-O111:NM | AMPR BCNR STRR TETR FFCR AMCI TIOI | Water |
| 299-O111:H8 | AMPR TIOR BCNR STRR TETR AMCI FFCI | Water |
| 356-O69:H11 | AMPR STRR SXTR TETR AMCI FFCI | Water |
| 373-O165:H25 | BCNR STRR TETR FFCR AMPI TIOI | Water |
| 374-O165:NM | BCNR STRR TETR FFCR AMPI | Water |
| 385-O111:NM | AMCR AMPR TIOR FOXR STRR SXTR CHLR TETR FFCR CROI GENI BCNI | Water |
| 391-O103:H2 | STRR SXTR CHLR TETR AMPI FFCI | Sediment |
| Prevalence (%) in All Isolates | Water/Sediment Isolates | Clinical Isolates | p Value Fisher’s Extract | Bonferroni Corrected p Value | |||
|---|---|---|---|---|---|---|---|
| Gene | (+) | (−) | (+) | (−) | |||
| ampC | 1.80 | 3 | 52 | 0 | 112 | 0.034 | 0.585 |
| gyrA83 | 1.80 | 0 | 55 | 3 | 109 | 0.552 | 1.000 |
| aac | 0.60 | 1 | 54 | 0 | 112 | 0.329 | 1.000 |
| aadA | 32.99 | 3 | 52 | 2 | 110 | 0.333 | 1.000 |
| aph | 16.17 | 12 | 43 | 15 | 97 | 0.184 | 1.000 |
| blaCMY-2 | 1.80 | 1 | 54 | 2 | 110 | 1.000 | 1.000 |
| blaCTX-M | 0.60 | 0 | 55 | 1 | 111 | 1.000 | 1.000 |
| blaTEM | 12.57 | 7 | 48 | 14 | 98 | 1.000 | 1.000 |
| catA | 0.60 | 1 | 54 | 0 | 112 | 0.329 | 1.000 |
| dfrA | 5.99 | 7 | 48 | 3 | 109 | 0.015 | 0.263 |
| floR | 1.20 | 1 | 54 | 1 | 111 | 0.552 | 1.000 |
| fosA | 2.40 | 3 | 52 | 1 | 111 | 0.105 | 1.000 |
| mphA | 1.20 | 2 | 53 | 0 | 112 | 0.107 | 1.000 |
| sul1 | 2.99 | 3 | 52 | 2 | 110 | 0.333 | 1.000 |
| sul2 | 13.17 | 9 | 46 | 13 | 99 | 0.466 | 1.000 |
| tetA | 8.98 | 5 | 50 | 10 | 102 | 1.000 | 1.000 |
| tetB | 7.78 | 8 | 47 | 5 | 107 | 0.031 | 0.531 |
| Isolate | Antibiotic Class/Group and Resistance Profile | ARDs Detected in the Sequences and Putative Role in Resistance 1 | Plasmid Replicon Sequences 2,3 |
|---|---|---|---|
| 296-O111:NM | Aminoglycosides STRR Folate pathway inhibitors SXTR Penicillins, penicillin +β-lactam inhibitors AMPR AMCI Phenicols FFCI Tetracyclines TETR NT 4 | aadA2, aph(3″)-Ib, aph(6)-Id drfA8, dfrA12, sul1, sul2 blaTEM-1B - tetA, tetB mphA | ColE1 (2), IncB/O/K/Z, IncFII(pHN7A8), IncX1 |
| 298-O111:NM | Aminoglycosides BCNR STRR Cephalosporins TIOI Penicillins, penicillin +β-lactam inhibitors AMPR AMCI Phenicols FFCR Tetracyclines TETR NT 4 | aph(3′)-Ia, aph(3″)-Ib, aph(6)-Id blaTEM-1B blaTEM-1B - tetA sul2 | Col156, IncB/O/K/Z, IncFII(pRSB107), IncQ1 |
| 299-O111:H8 | Aminoglycosides BCNR STRR Cephalosprins TIOR Penicillins, penicillin +β-lactam inhibitors AMPR AMCI Phenicols FFCI Tetracyclines TETR NT 4 | aph(3′)-Ia, s aph(3″)-Ib, aph(6)-Id blaTEM-1B blaTEM-1B - tetA sul2 | Col156, IncB/O/K/Z, IncFII(pRSB107), IncQ1 |
| 356-O69:H11 | Aminoglycosides STRR Folate pathway inhibitors SXTR Penicillins penicillin +β-lactam inhibitors AMPR AMCI Phenicols FFCI Tetracyclines TETR | aph(3″)-Ib, aph(6)-Id dfrA8, sul2 blaTEM-1B - tetB | IncB/O/K/Z, IncFII(pHN7A8), IncFIB(AP001918) |
| 373-O165:H25 | Aminoglycosides BCNR STRR Cephalosporins TIOI Penicillins AMPI Phenicols FFCR Tetracyclines TETR | aph(3′)-1a, aph(3″)-Ib, aph(6)-Id - - - tetB | IncFII, IncFIB(AP001918) |
| 374-O165:NM | Aminoglycosides BCNR STRR Phenicols FFCR Penicillins AMPI Tetracyclines TETR | aph(3′)-Ia, aph(3″)-Ib,aph(6)-Id - - tetB | IncFII, IncFIB(AP001918), |
| 385-O111:NM | Aminoglycosides STRR BCNI GENI Cephalosporins TIOR CROI Cephamycins FOXR Penicillins, penicillin + β-lactam inhibitors AMPR AMCR Folate pathway inhibitors SXTR Phenicols CHLR FFCR Tetracyclines TETR NT | aac(3)-VIa, aadA1, aadA2, aph(3″)-Ib, aph(6)-Id blaCMY-2 blaCMY-2 dfrA12, sul1, sul2 floR tetA mphA | ColEI, IncB/O/K/Z, IncFII(pRSB107), IncY |
| 391-O103:H2 | Aminoglycosides STRR Folate pathway inhibitors SXTR Phenicols CHLR FFCI Penicillin AMPI Tetracyclines TETR | aadA1, aph(3″)-Ib, aph(6)-Id dfrA1, sul1, sul2 catA1 - tetB | ColEI, IncB/O/K/Z |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Chen, J.; Fong, K.; Nadya, S.; Allen, K.; Laing, C.; Ziebell, K.; Topp, E.; Carroll, L.M.; Wiedmann, M.; et al. Antibiotic Resistance in Shiga Toxigenic Escherichia coli Isolates from Surface Waters and Sediments in a Mixed Use Urban Agricultural Landscape. Antibiotics 2021, 10, 237. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030237
Ma Y, Chen J, Fong K, Nadya S, Allen K, Laing C, Ziebell K, Topp E, Carroll LM, Wiedmann M, et al. Antibiotic Resistance in Shiga Toxigenic Escherichia coli Isolates from Surface Waters and Sediments in a Mixed Use Urban Agricultural Landscape. Antibiotics. 2021; 10(3):237. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030237
Chicago/Turabian StyleMa, Yvonne, Jessica Chen, Karen Fong, Stephanie Nadya, Kevin Allen, Chad Laing, Kim Ziebell, Ed Topp, Laura M. Carroll, Martin Wiedmann, and et al. 2021. "Antibiotic Resistance in Shiga Toxigenic Escherichia coli Isolates from Surface Waters and Sediments in a Mixed Use Urban Agricultural Landscape" Antibiotics 10, no. 3: 237. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030237