
Comparative Investigation of Composition, Antifungal, and Anti-Inflammatory Effects of the Essential Oil from Three Industrial Hemp Varieties from Italian Cultivation

 ,
, 

 , ,
, ,  ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Pharmacognostic Analysis
3.2. HPLC–UV-MS Determination of Terpenophenolic Compounds
3.3. Antityrosinase Activity
3.4. Antimycotic Activities
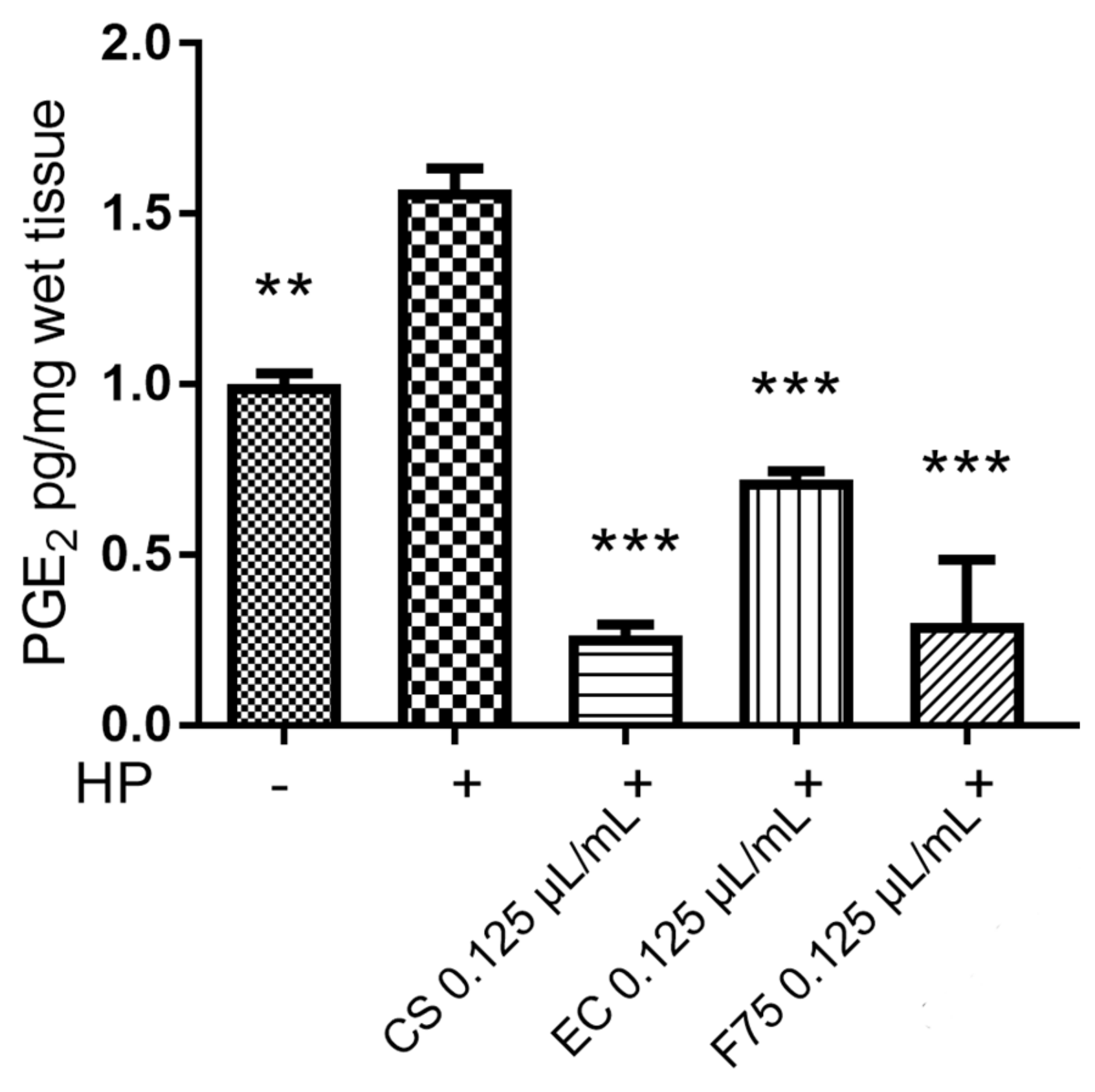
3.5. Ex Vivo Model of Hydrogen-Peroxide-Induced Toxicity in Isolated Mouse Skin Tissue
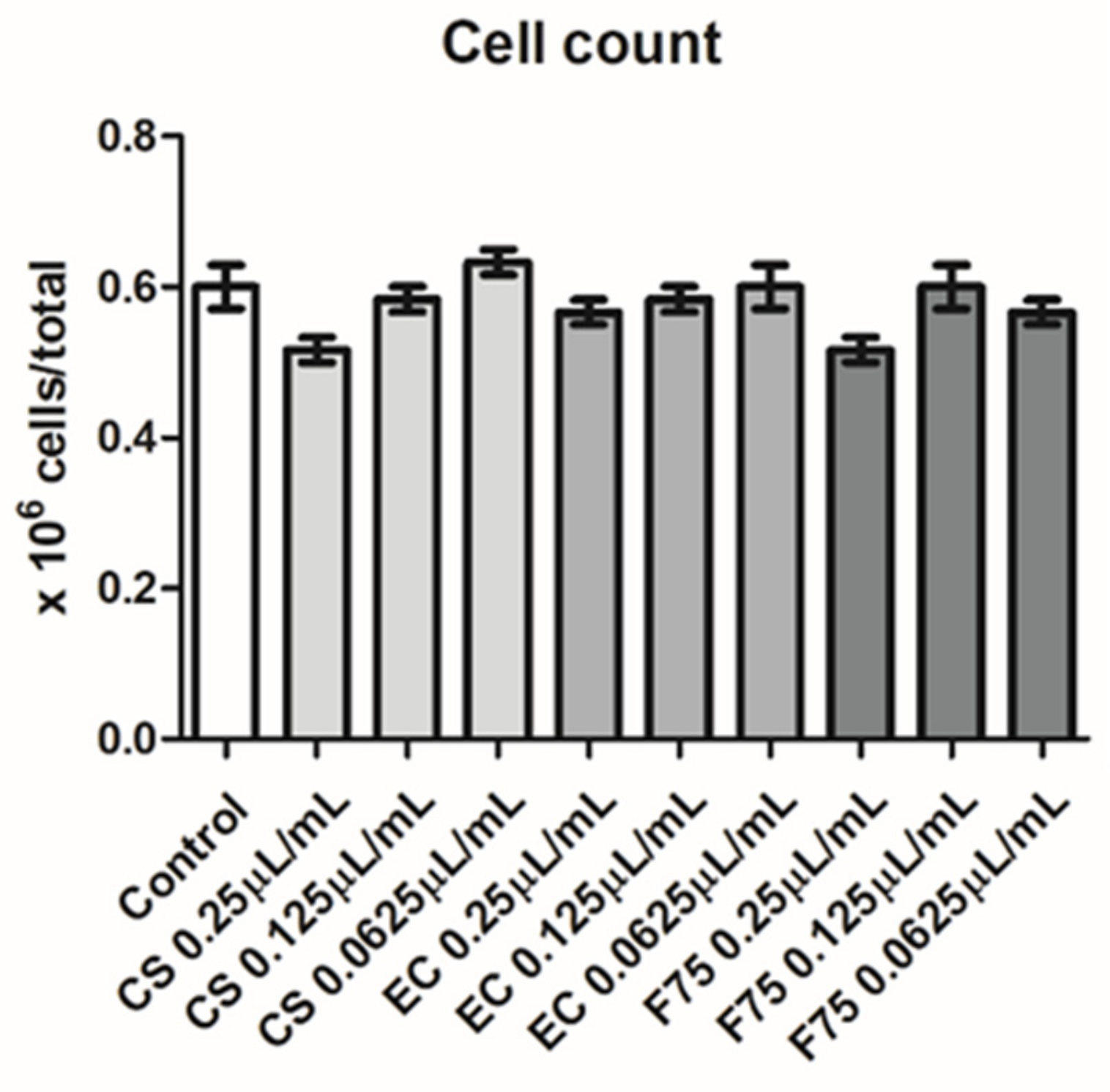
3.6. Cell Culture
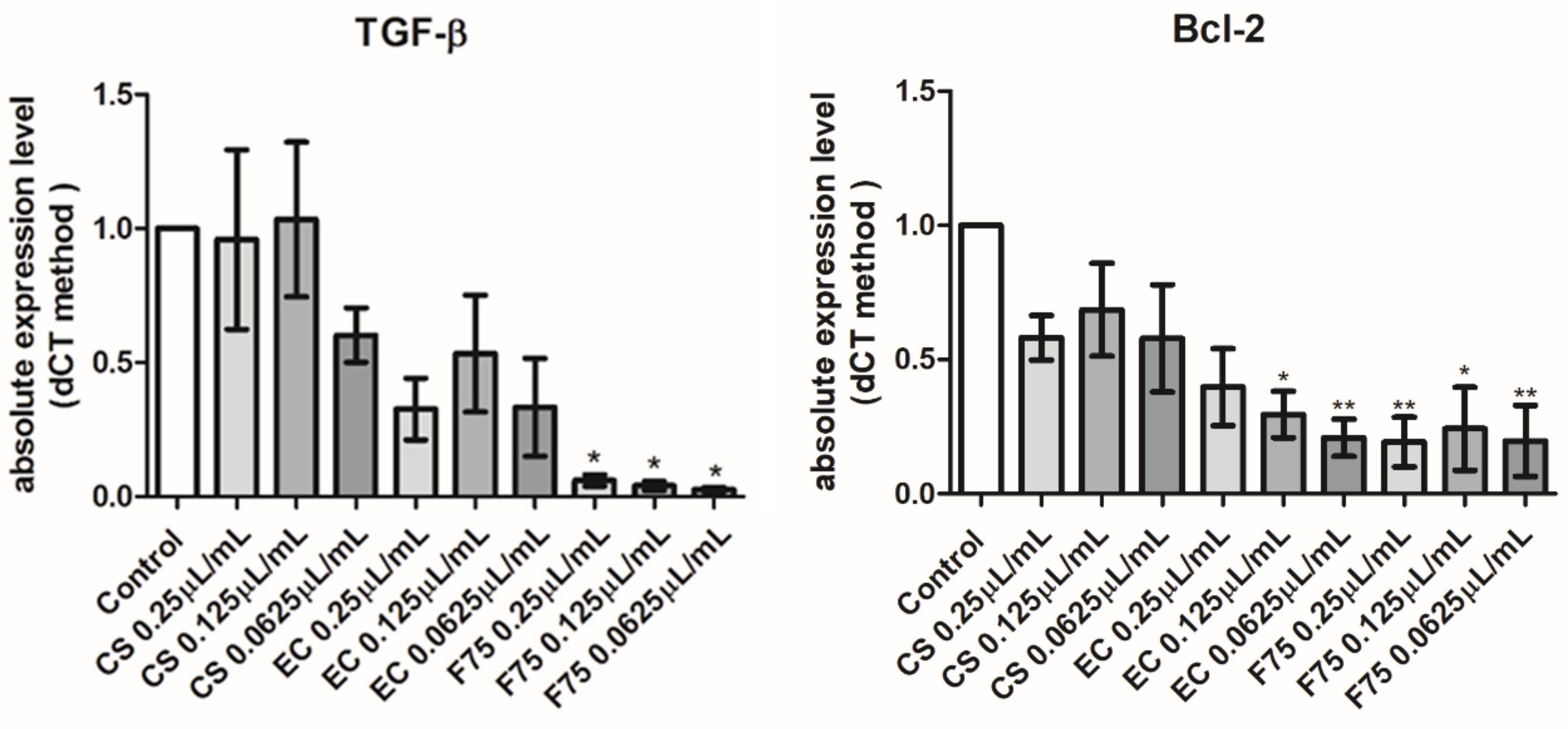
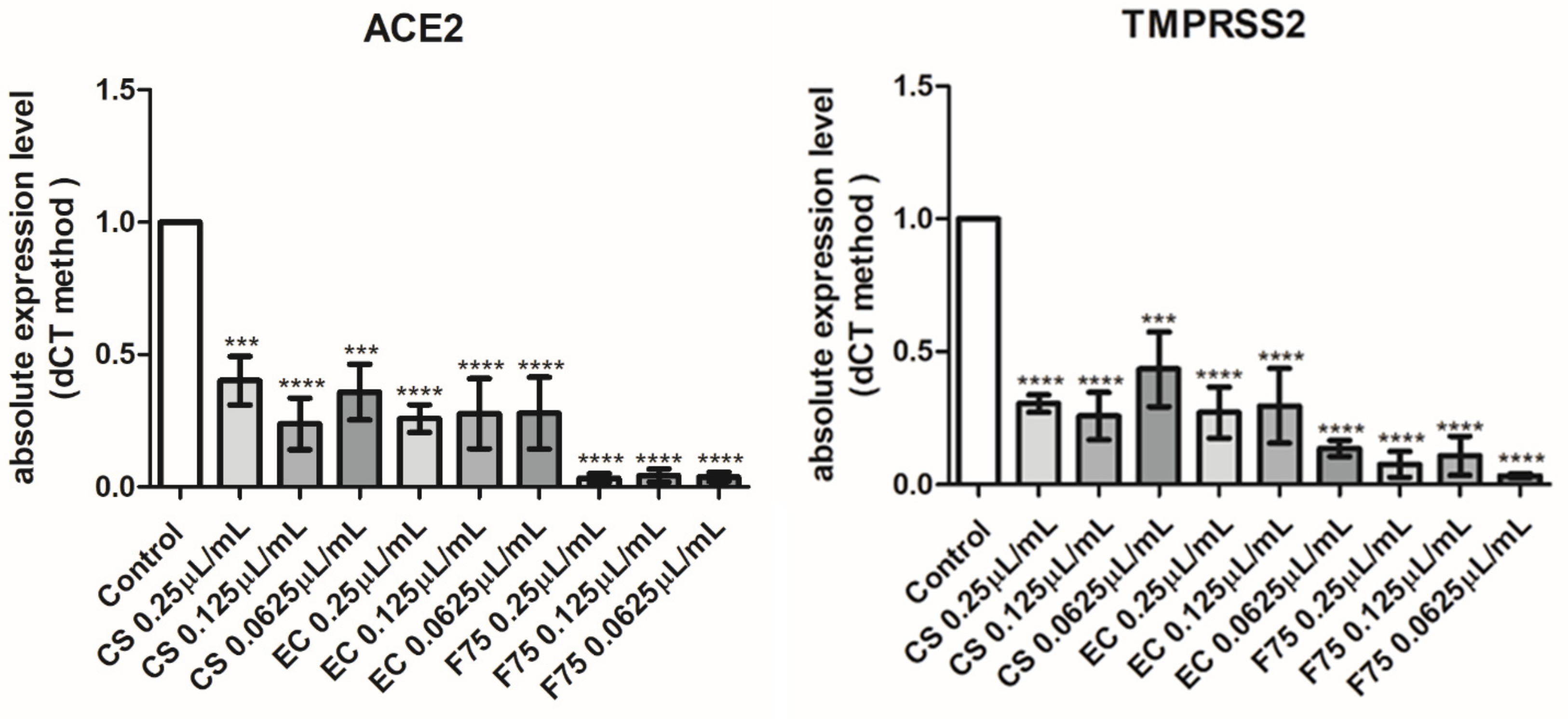
3.7. Real-Time Reverse Transcription Polymerase Chain Reaction (Real-Time RT PCR)
3.8. Bioinformatics
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Montserrat-de la Paz, S.; Marín-Aguilar, F.; García-Gimenez, M.D.; Fernández-Arche, M. Hemp (Cannabis sativa L.) seed oil: Analytical and phytochemical characterization of the unsaponifiable fraction. J. Agric. Food Chem. 2014, 62, 1105–1110. [Google Scholar] [CrossRef]
- Vonapartis, E.; Aubin, M.-P.; Seguin, P.; Mustafa, A.F.; Charron, J.-B. Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Compos. Anal. 2015, 39, 8–12. [Google Scholar] [CrossRef]
- Citti, C.; Linciano, P.; Russo, F.; Luongo, L.; Iannotta, M.; Maione, S.; Laganà, A.; Capriotti, A.L.; Forni, F.; Vandelli, M.A.; et al. A novel phytocannabinoid isolated from Cannabis sativa L. with an in vivo cannabimimetic activity higher than Δ9-tetrahydrocannabinol: Δ9-Tetrahydrocannabiphorol. Sci. Rep. 2019, 9, 20335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiralan, M.; Gül, V.; Metin Kara, S. Fatty acid composition of hemp seed oils from different locations in Turkey. Span. J. Agric. Res. 2010, 8, 385–390. [Google Scholar] [CrossRef]
- Amaducci, S.; Scordia, D.; Liu, F.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S. Key cultivation techniques for hemp in Europe and China. Ind. Crop. Prod. 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Bertoli, A.; Tozzi, S.; Pistelli, L.; Angelini, L.G. Fibre hemp inflorescences: From crop-residues to essential oil production. Ind. Crop. Prod. 2010, 32, 329–337. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Sagredo, O.; Pazos, M.R.; García, C.; Pertwee, R.; Mechoulam, R.; Martínez-Orgado, J. Cannabidiol for neurodegenerative disorders: Important new clinical applications for this phytocannabinoid? Br. J. Clin. Pharmacol. 2013, 75, 323–333. [Google Scholar] [CrossRef]
- Zengin, G.; Menghini, L.; Di Sotto, A.; Mancinelli, R.; Sisto, F.; Carradori, S.; Cesa, S.; Fraschetti, C.; Filippi, A.; Angiolella, L.; et al. Chromatographic Analyses, In Vitro Biological Activities, and Cytotoxicity of Cannabis sativa L. Essential Oil: A Multidisciplinary Study. Molecules 2018, 23, 3266. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, C.; Recinella, L.; Ronci, M.; Menghini, L.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Di Iorio, L.; Carradori, S.; Tirillini, B.; et al. Multiple pharmacognostic characterization on hemp commercial cultivars: Focus on inflorescence water extract activity. Food Chem. Toxicol. 2019, 125, 452–461. [Google Scholar] [CrossRef]
- Orlando, G.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Carradori, S.; Di Simone, S.; Ciferri, M.C.; Zengin, G.; Ak, G.; et al. Water Extract from Inflorescences of Industrial Hemp Futura 75 Variety as a Source of Anti-Inflammatory, Anti-Proliferative and Antimycotic Agents: Results from In Silico, In Vitro and Ex Vivo Studies. Antioxidants 2020, 9. [Google Scholar] [CrossRef]
- Di Giacomo, V.; Recinella, L.; Chiavaroli, A.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Politi, M.; Antolini, M.D.; Acquaviva, A.; et al. Metabolomic Profile and Antioxidant/Anti-Inflammatory Effects of Industrial Hemp Water Extract in Fibroblasts, Keratinocytes and Isolated Mouse Skin Specimens. Antioxidants 2021, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kovalchuk, A.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, Y.; Kovalchuk, I.; Kovalchuk, O. In search of preventive strategies: Novel high-CBD Cannabis sativa extracts modulate ACE2 expression in COVID-19 gateway tissues. Aging 2020, 12, 22425–22444. [Google Scholar] [CrossRef] [PubMed]
- Anil, S.M.; Shalev, N.; Vinayaka, A.C.; Nadarajan, S.; Namdar, D.; Belausov, E.; Shoval, I.; Mani, K.A.; Mechrez, G.; Koltai, H. Cannabis compounds exhibit anti-inflammatory activity in vitro in COVID-19-related inflammation in lung epithelial cells and pro-inflammatory activity in macrophages. Sci Rep. 2021, 11, 1462. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in ahigh-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef]
- Turner, A.J.; Hiscox, J.A.; Hooper, N.M. ACE2: From vasopeptidase to SARS virus receptor. Trends Pharm. Sci. 2004, 25, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Sungnak, W.; Huang, N.; Bécavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. HCA Lung Biological Network. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Isa-Isa, R.; Arenas, R.; Isa, M. Inflammatory tinea capitis: Kerion, dermatophytic granuloma, and mycetoma. Clin. Dermatol. 2010, 28, 133–136. [Google Scholar] [CrossRef]
- Zhang, M.; Jiang, L.; Li, F.; Xu, Y.; Lv, S.; Wang, B. Simultaneous dermatophytosis and keratomycosis caused by Trichophyton interdigitale infection: A case report and literature review. BMC Infect. Dis. 2019, 19, 1–8. [Google Scholar] [CrossRef] [Green Version]
- El-Zawawy, N.A.; Ali, S.S. Pyocyanin as anti-tyrosinase and anti tinea corporis: A novel treatment study. Microb. Pathog. 2016, 100, 213–220. [Google Scholar] [CrossRef]
- Kolbe, L.; Mann, T.; Gerwat, W.; Batzer, J.; Ahlheit, S.; Scherner, C.; Wenck, H.; Stäb, F. 4-n-butylresorcinol, a highly effective tyrosinase inhibitor for the topical treatment of hyperpigmentation. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 19–23. [Google Scholar] [CrossRef]
- Chiavaroli, A.; Di Simone, S.C.; Sinan, K.I.; Ciferri, M.C.; Angeles Flores, G.; Zengin, G.; Etienne, O.K.; Ak, G.; Fawzi Mahomoodally, M.; Jugreet, S.; et al. Pharmacological Properties and Chemical Profiles of Passiflora foetida L. Extracts: Novel Insights for Pharmaceuticals and Nutraceuticals. Processes 2020, 8, 1034. [Google Scholar] [CrossRef]
- Dong, Z.; Zhang, L.; Xu, W.; Zhang, G. EGFR may participate in immune evasion through regulation of B7-H5 expression in non-small cell lung carcinoma. Mol. Med. Rep. 2018, 18, 3769–3779. [Google Scholar] [CrossRef]
- Liu, S.; Ren, J.; Ten Dijke, P. Targeting TGFβ signal transduction for cancer therapy. Transduct. Target. Ther. 2021, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Atalay, S.; Gęgotek, A.; Wroński, A.; Domigues, P.; Skrzydlewska, E. Therapeutic application of cannabidiol on UVA and UVB irradiated rat skin. A Proteomic Study. J. Pharm. Biomed. Anal. 2021, 192, 113656. [Google Scholar] [CrossRef] [PubMed]
- Menghini, L.; Ferrante, C.; Carradori, S.; D’Antonio, M.; Orlando, G.; Cairone, F.; Cesa, S.; Filippi, A.; Fraschetti, C.; Zengin, G.; et al. Chemical and Bioinformatics Analyses of the Anti-Leishmanial and Anti-Oxidant Activities of Hemp Essential Oil. Biomolecules 2021, 11, 272. [Google Scholar] [CrossRef] [PubMed]
- Zheljazkov, V.D.; Sikora, V.; Semerdjieva, I.B.; Kačániová, M.; Astatkie, T.; Dincheva, I. Grinding and Fractionation during Distillation Alter Hemp Essential Oil Profile and Its Antimicrobial Activity. Molecules 2020, 25, 3943. [Google Scholar] [CrossRef] [PubMed]
- Nagy, D.U.; Cianfaglione, K.; Maggi, F.; Sut, S.; Dall’Acqua, S. Chemical Characterization of Leaves, Male and Female Flowers from Spontaneous Cannabis (Cannabis sativa L.) Growing in Hungary. Chem. Biodivers. 2019, 16, e1800562. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Michel, L.; Chaumont, J.P.; Millet-Clerc, J. Use of caryophyllene oxide as an antifungal agent in an in vitro experimental model of onychomycosis. Mycopathologia 1999, 148, 79–82. [Google Scholar] [CrossRef]
- Chatatikun, M.; Supjaroen, P.; Promlat, P.; Chantarangkul, C.; Waranuntakul, S.; Nawarat, J.; Tangpong, J.; Chiabchalard, A. Antioxidant and tyrosinase inhibitor properties of an aqueous extract og Garcinia atrovirdis Griff. Ex. T. Anderson fruit pericarps. Pharm. J. 2020, 12, 71–78. [Google Scholar]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Schuster, N.; Krieglstein, K. Mechanisms of TGF-beta-mediated apoptosis. Cell Tissue Res. 2002, 307, 1–14. [Google Scholar] [CrossRef]
- Senichkin, V.V.; Pervushin, N.V.; Zuev, A.P.; Zhivotovsky, B.; Kopeina, G.S. Target. Bcl-2 Fam. Proteins: WhatWhereWhen? Biochemistry 2020, 85, 1210–1226. [Google Scholar] [CrossRef]
- Zarubin, A.; Stepanov, V.; Markov, A.; Kolesnikov, N.; Marusin, A.; Khitrinskaya, I.; Swarovskaya, M.; Litvinov, S.; Ekomasova, N.; Dzhaubermezov, M.; et al. Structural Variability, Expression Profile, and Pharmacogenetic Properties of TMPRSS2 Gene as a Potential Target for COVID-19 Therapy. Genes 2020, 12, E19. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Jabeen, N.; Amanullah, A.; Baig, A.A.; Aziz, B.; Shabbir, S.; Raza, F.; Uddin, N. Molecular docking between human TMPRSS2 and SARS-CoV-2 spike protein: Conformation and intermolecular interactions. Aims Microbiol. 2020, 6, 350–360. [Google Scholar] [CrossRef]
- Chiavaroli, A.; Recinella, L.; Ferrante, C.; Locatelli, M.; Macchione, N.; Zengin, G.; Leporini, L.; Leone, S.; Martinotti, S.; Brunetti, L.; et al. Crocus sativus, Serenoa repens and Pinus massoniana extracts modulate inflammatory response in isolated rat prostate challenged with LPS. J. Biol. Regul. Homeost. Agents 2017, 31, 531–541.15. [Google Scholar] [PubMed]
- Ferrante, C.; Angelini, P.; Venanzoni, R.; Angeles Flores, G.; Tirillini, B.; Recinella, L.; Chiavaroli., A.; Brunetti, L.; Leone, S.; Di Simone, S.C.; et al. Antimicrobial, Antioxidant, and Antiproliferative Effects of Coronilla minima: An Unexplored Botanical Species. Antibiotics 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Menghini, L.; Ferrante, C.; Leporini, L.; Recinella, L.; Chiavaroli, A.; Leone, S.; Pintore, G.; Vacca, M.; Orlando, G.; Brunetti, L. An Hydroalcoholic Chamomile Extract Modulates Inflammatory and Immune Response in HT29 Cells and Isolated Rat Colon. Phytother. Res. 2016, 30, 1513–1518. [Google Scholar] [CrossRef]
- Angelini, P.; Venanzoni, R.; Angeles Flores, G.; Tirillini, B.; Orlando, G.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Di Simone, S.C.; et al. Evaluation of Antioxidant, Antimicrobial and Tyrosinase Inhibitory Activities of Extracts from Tricholosporum goniospermum, an Edible Wild Mushroom. Antibiotics 2020, 9. [Google Scholar] [CrossRef]
- Ferrante, C.; Chiavaroli, A.; Angelini, P.; Venanzoni, R.; Angeles Flores, G.; Brunetti, L.; Petrucci, M.; Politi, M.; Menghini, L.; Leone, S.; et al. Phenolic Content and Antimicrobial and Anti-Inflammatory Effects of Solidago virga-aurea, Phyllanthus niruri, Epilobium angustifolium, Peumus boldus, and Ononis spinosa Extracts. Antibiotics 2020, 9, 783. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Area % | RI a | RIL b |
|---|---|---|---|
| α-pinene | 11.9 | 934 | 933 |
| camphene | 0.3 | 949 | 953 |
| β-pinene | 3.6 | 977 | 978 |
| myrcene | 6.8 | 994 | 991 |
| α-phellandrene | 0.1 | 1006 | 1007 |
| δ-3-carene | 0.5 | 1011 | 1009 |
| α-terpinene | 0.1 | 1019 | 1018 |
| para-cymene | 0.2 | 1028 | 1024 |
| limonene | 2.2 | 1031 | 1030 |
| eucalyptol | 0.4 | 1040 | 1032 |
| Z-β-ocimene | 0.5 | 1043 | 1035 |
| E-β-ocimene | 1.9 | 1053 | 1046 |
| γ-terpinene | 0.2 | 1062 | 1058 |
| terpinolene | 2.3 | 1091 | 1086 |
| linalyl anthranilate | 0.2 | 1110 | 1104 |
| fenchyl alcohol | 0.1 | 1123 | 1123 |
| terpinen-4-ol | 0.2 | 1187 | 1184 |
| α-ylangene | 0.2 | 1378 | 1371 |
| Z-caryophyllene | 0.3 | 1414 | 1413 |
| E-caryophyllene | 13.5 | 1428 | 1424 |
| α-trans-bergamotene | 0.6 | 1442 | 1432 |
| α-humulene | 5.3 | 1462 | 1454 |
| 9-epi-E-caryophyllene | 0.7 | 1470 | 1464 |
| γ-muurolene | 0.5 | 1485 | 1478 |
| α-amorphene | 0.2 | 1488 | 1482 |
| β-selinene | 1.5 | 1495 | 1492 |
| α-selinene | 1.7 | 1504 | 1501 |
| Z-γ-bisabolene | 1.4 | 1516 | 1511 |
| γ-cadinene | 0.4 | 1523 | 1512 |
| selina-4(15),7(11)-diene | 1.3 | 1545 | 1540 |
| selina-3,7(11)-diene | 2.5 | 1552 | 1546 |
| B-germacrene | 0.3 | 1569 | 1557 |
| caryophyllene oxide | 2.2 | 1599 | 1587 |
| humulene epoxide | 0.7 | 1626 | 1613 |
| α-bisabolol | 0.5 | 1697 | 1688 |
| tetracosane | 6.0 | 2406 | 2400 |
| heptacosane | 23.9 | 2690 | 2700 |
| 12 unknown compounds | 4.8 |
| Compound | Area % | RI a | RIL b |
|---|---|---|---|
| α-pinene | 14.9 | 934 | 933 |
| camphene | 0.3 | 949 | 953 |
| β-pinene | 3.8 | 977 | 978 |
| myrcene | 11.8 | 994 | 991 |
| α-phellandrene | 0.2 | 1006 | 1007 |
| δ-3-carene | 0.5 | 1011 | 1009 |
| α-terpinene | 0.2 | 1018 | 1018 |
| para-cymene | 0.1 | 1028 | 1025 |
| limonene | 1.8 | 1031 | 1030 |
| eucalyptol | 0.2 | 1041 | 1032 |
| E-β-ocimene | 2.9 | 1053 | 1046 |
| γ-terpinene | 0.2 | 1062 | 1058 |
| terpinolene | 5.1 | 1091 | 1086 |
| Z-caryophyllene | 0.5 | 1414 | 1413 |
| α-cis-bergamotene | 0.3 | 1422 | 1416 |
| E-caryophyllene | 19.3 | 1428 | 1424 |
| α-trans-bergamotene | 1.9 | 1442 | 1432 |
| α-humulene | 8.3 | 1462 | 1454 |
| 9-epi-caryophyllene | 1.1 | 1470 | 1464 |
| β-selinene | 1.7 | 1495 | 1492 |
| α-selinene | 1.3 | 1504 | 1501 |
| selina-4(15),7(11)-diene | 0.9 | 1545 | 1540 |
| selina-3,7(11)-diene | 1.5 | 1552 | 1546 |
| caryophyllene oxide | 4.3 | 1599 | 1587 |
| humulene epoxide | 1.1 | 1626 | 1613 |
| allo-aromadendrene epoxide | 0.4 | 1650 | 1644 |
| tetracosane | 8.8 | 2407 | 2400 |
| 13 unknown compounds | 6.6 |
| Compound | Area % | RI a | RIL b |
|---|---|---|---|
| α-pinene | 12.6 | 934 | 933 |
| camphene | 0.3 | 949 | 953 |
| β-pinene | 4.1 | 977 | 978 |
| myrcene | 26.4 | 995 | 991 |
| α-phellandrene | 0.3 | 1006 | 1007 |
| δ-3-carene | 0.3 | 1012 | 1009 |
| α-terpinene | 0.3 | 1019 | 1018 |
| para-cymene | 0.2 | 1028 | 1025 |
| limonene | 4.7 | 1031 | 1030 |
| eucalyptol | 0.5 | 1041 | 1032 |
| Z-β-ocimene | 0.4 | 1043 | 1035 |
| E-β-ocimene | 2.5 | 1053 | 1046 |
| γ-terpinene | 0.3 | 1062 | 1058 |
| terpinolene | 7.0 | 1091 | 1086 |
| linalyl anthranilate | 0.3 | 1110 | 1104 |
| fenchyl-alcohol | 0.2 | 1123 | 1123 |
| Z-caryophyllene | 0.4 | 1414 | 1413 |
| E-caryophyllene | 19.1 | 1428 | 1424 |
| α-trans-bergamotene | 0.2 | 1442 | 1432 |
| α-humulene | 7.2 | 1462 | 1454 |
| 9-epi-E-caryophyllene | 0.6 | 1470 | 1464 |
| β-selinene | 1.3 | 1495 | 1492 |
| α-selinene | 1.1 | 1504 | 1501 |
| Z-γ-bisabolene | 1.1 | 1514 | 1511 |
| selina-4(15),7(11)-diene | 0.3 | 1545 | 1540 |
| selina-3,7(11)-diene | 0.5 | 1552 | 1546 |
| caryophyllene oxide | 3.2 | 1599 | 1587 |
| humulene epoxide | 1.0 | 1626 | 1613 |
| 10 unknown compounds | 3.6 |
| Dermatophytes Species | MIC * (µg/mL) | |||
|---|---|---|---|---|
| Carmagnola Selezionata | Eletta Campana | Futura 75 | Griseofulvin | |
| T. mentagrophytes (CCF 4823) | 0.79(0.625–1.25) | < 0.312 | 0.79(0.625–1.25) | 2.52 (2–4) |
| T. rubrum (CCF 4879) | 3.15(2.5–5) | 6.3(5–10) | 6.3(5–10) | 3.175(2–4) |
| T. rubrum (CCF 4933) | 0.99(0.625–1.25) | 1.57(1.25–2.5) | 0.99(0.625–1.25) | 1.26(1–2) |
| A. crocatum (CCF 5300) | 1.57(1.25–2.5) | 1.57(1.25–2.5) | 0.39(0.625–0.312) | >8 |
| A. quadrifidum (CCF 5792) | 3.15(2.5–5) | 3.15(2.5–5) | 0.39(0.625–0.312) | >8 |
| A. gypseum (CCF 6261) | 1.57(1.25–2.5) | 3.15(2.5–5) | 0.49(0.312–0.625) | 3.174 (2–4) |
| A. curreyi (CCF 5207) | 1.98(1.25–2.5) | 6.3(5–10) | 1.57(1.25–2.5) | >8 |
| A. insingulare (CCF 5417) | 6.3(5–10) | 0.99(0.625–1.25) | 0.79(0.625–1.25) | >8 |
| EO | Tyrosinase inhibition (mg KAE/g oil) |
|---|---|
| EC | 31.73 ± 0.63 a |
| F75 | 29.41 ± 0.61 b |
| CS | 21.31 ± 1.37 c |
| EO | DPPH (mg TE/g EO) | ABTS (mg TE/g EO) | CUPRAC (mg TE/g EO) | FRAP (mg TE/g EO) | Metal Chelating (mg EDTAE/g EO) | Phosphomolybdenum (mmol TE/g EO) |
|---|---|---|---|---|---|---|
| EC | 2.53±0.23 a* | 32.44 ± 0.03 a | 45.51 ± 0.75 a | 19.29 ± 0.34 a | 11.55 ± 0.84 a | 17.95 ± 0.34 b |
| CS | 1.18 ± 0.09 c | 32.15 ± 0.08 b | 29.91 ± 0.95 c | 13.55 ± 0.46 c | 7.19 ± 0.42 b | 17.52 ± 1.12 b |
| F75 | 2.11 ± 0.15 b | 32.47 ± 0.04 a | 35.05 ± 0.85 b | 16.16 ± 0.47 b | 10.84 ± 0.46 a | 18.80 ± 0.47 a |
| EO | GPS Coordinates | Extension (sqm) | Previous Crops (2 Years) | Sowing Scheme (cm Inter/Intra Lines) | Sowing Density |
|---|---|---|---|---|---|
| F75 | 42.363059, 14.093390 | 4000 | Wheat/alfalfa | 20 cm/50 cm | 15 Kg/ha |
| CS | 42.359222, 14.108528 | 14,000 | Wheat/ Wheat | 40 cm/50 cm | 20 Kg/ha |
| EC | 42.343824, 14.102808 | 6000 | Not cultivated | 20 cm/40 cm | 20 Kg/ha |
| Time (min) | Flow (mL/min) | %A | %B |
|---|---|---|---|
| 0 | 0.750 | 32.5 | 67.5 |
| 0.5 | 0.750 | 32.5 | 67.5 |
| 14 | 0.750 | 7 | 93 |
| 22 | 0.750 | 7 | 93 |
| 22.1 | 1.05 | 32.5 | 67.5 |
| 28 | 1.05 | 32.5 | 67.5 |
| 28.1 | 0.750 | 32.5 | 67.5 |
| 30 | 0.750 | 32.5 | 67.5 |
| Standard | m/z | Wavelength (nm) | Retention Time (min) | |
|---|---|---|---|---|
| 1 | CBDA | 357.3 | 230 | 12.4 |
| 2 | CBGA | 343.3–260.1 | 230 | 13.6 |
| 3 | CBG | 317.3–234.1–193.1 | 230 | 15.4 |
| 4 | CBD | 315.2 | 230 | 15.6 |
| 5 | CBN | 311.3–293.25 | 230 | 20.0 |
| 6 | THC-d3 | 318.9; [315.2: EO pool of THC] | 230 | 20.6 |
| 7 | CBC | 315.3–259.13–193.13 | 230 | 21.2 |
| 8 | THCA | 341.3 | 230 | 22.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlando, G.; Adorisio, S.; Delfino, D.; Chiavaroli, A.; Brunetti, L.; Recinella, L.; Leone, S.; D’Antonio, M.; Zengin, G.; Acquaviva, A.; et al. Comparative Investigation of Composition, Antifungal, and Anti-Inflammatory Effects of the Essential Oil from Three Industrial Hemp Varieties from Italian Cultivation. Antibiotics 2021, 10, 334. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030334
Orlando G, Adorisio S, Delfino D, Chiavaroli A, Brunetti L, Recinella L, Leone S, D’Antonio M, Zengin G, Acquaviva A, et al. Comparative Investigation of Composition, Antifungal, and Anti-Inflammatory Effects of the Essential Oil from Three Industrial Hemp Varieties from Italian Cultivation. Antibiotics. 2021; 10(3):334. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030334
Chicago/Turabian StyleOrlando, Giustino, Sabrina Adorisio, Domenico Delfino, Annalisa Chiavaroli, Luigi Brunetti, Lucia Recinella, Sheila Leone, Marianna D’Antonio, Gokhan Zengin, Alessandra Acquaviva, and et al. 2021. "Comparative Investigation of Composition, Antifungal, and Anti-Inflammatory Effects of the Essential Oil from Three Industrial Hemp Varieties from Italian Cultivation" Antibiotics 10, no. 3: 334. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030334