
Quantity and Quality of Aquaculture Enrichments Influence Disease Epidemics and Provide Ecological Alternatives to Antibiotics

 , , and
, , and
Abstract
:
1. Introduction
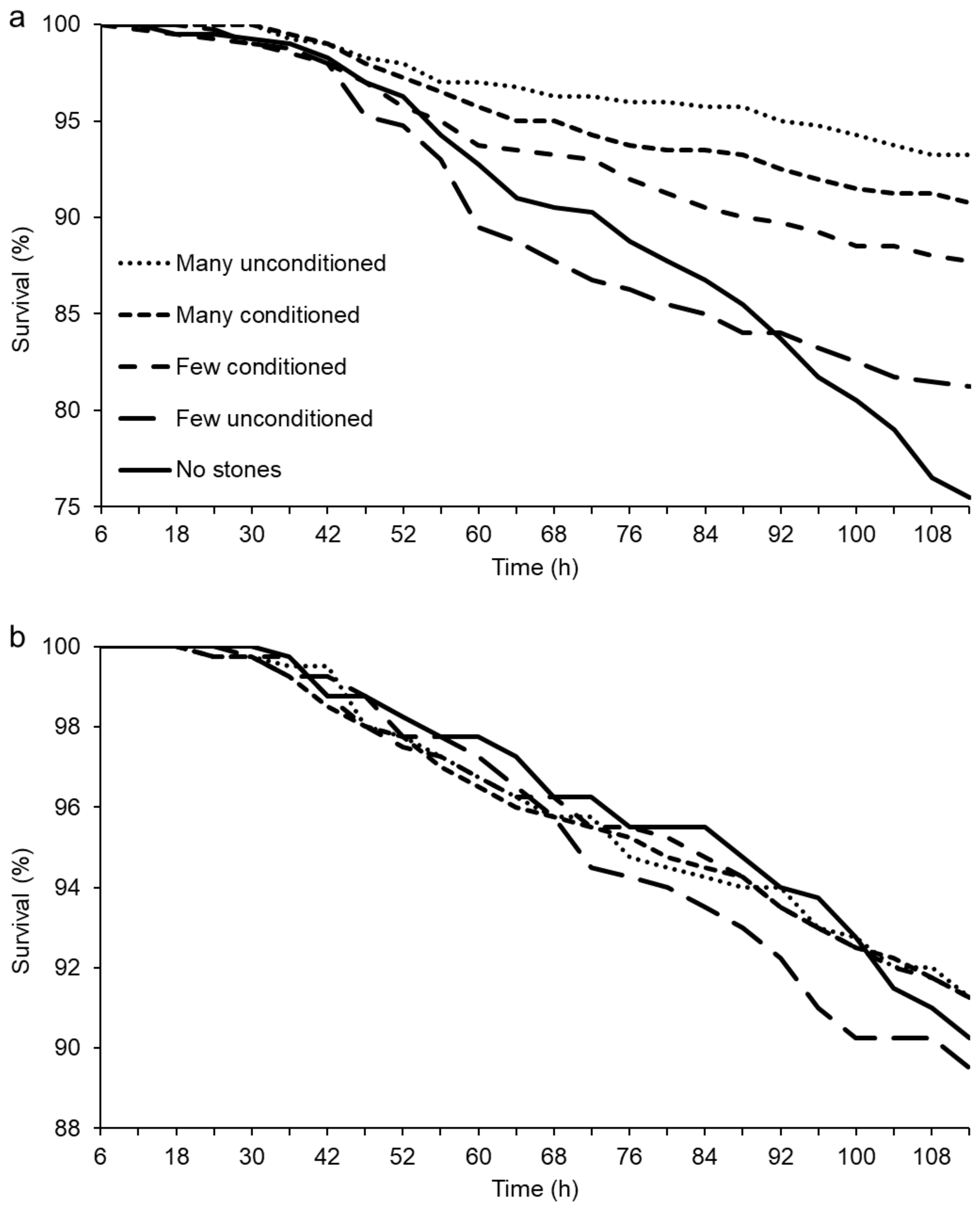
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savary, S.; Ficke, A.; Aubertot, J.N.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Strange, R.N.; Scott, P.R. Plant disease: A threat to global food security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef] [PubMed]
- Stentiford, G.D.; Sritunyalucksana, K.; Flegel, T.W.; Williams, B.A.P.; Withyachumnarnkul, B.; Itsathitphaisarn, O.; Bass, D. New Paradigms to Help Solve the Global Aquaculture Disease Crisis. PLoS Pathog. 2017, 13, e1006160. [Google Scholar] [CrossRef]
- Bondad-Reantaso, M.G.; Subasinghe, R.P.; Arthur, J.R.; Ogawa, K.; Chinabut, S.; Adlard, R.; Tan, Z.; Shariff, M. Disease and health management in Asian aquaculture. Vet. Parasitol. 2005, 132, 249–272. [Google Scholar] [CrossRef] [PubMed]
- Karvonen, A.; Rintamäki, P.; Jokela, J.; Valtonen, E.T. Increasing water temperature and disease risks in aquatic systems: Climate change increases the risk of some, but not all, diseases. Int. J. Parasitol. 2010, 40, 1483–1488. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Porter, J.W.; Ford, S.E. Are diseases increasing in the ocean? Annu. Rev. Ecol. Evol. Syst. 2004, 35, 31–54. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.; Shinn, A.P.; Grenfell, S.E.; Bron, J.E.; Burnell, G.; Cook, E.J.; Crumlish, M.; Culloty, S.; Davidson, K.L.; Ellis, R.P.; et al. Review of climate change impacts on marine aquaculture in the UK and Ireland. Aquat. Conserv. Mar. Freshw. Ecosyst. 2012, 22, 389–421. [Google Scholar] [CrossRef]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Martinez, J.L. Environmental pollution by antibiotics and by antibiotic resistance determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef]
- Archie, E.A.; Luikart, G.; Ezenwa, V.O. Infecting epidemiology with genetics: A new frontier in disease ecology. Trends Ecol. Evol. 2009, 24, 21–30. [Google Scholar] [CrossRef]
- Parratt, S.R.; Numminen, E.; Laine, A.-L. Infectious disease dynamics in heterogeneous landscapes. Ann. Rev. Ecol. Evol. Syst. 2016, 47, 283–306. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Glass, G.E.; Keesing, F. Spatial epidemiology: An emerging (or re-emerging) discipline. Trends Ecol. Evol. 2005, 20, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K. Landscape epidemiology of vector-borne diseases. Annu. Rev. Entomol. 2010, 55, 461–483. [Google Scholar] [CrossRef] [Green Version]
- Laine, A.L.; Burdon, J.J.; Dodds, P.N.; Thrall, P.H. Spatial variation in disease resistance: From molecules to metapopulations. J. Ecol. 2010, 99, 96–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penczykowski, R.M.; Parratt, S.R.; Barres, B.; Sallinen, S.K.; Laine, A.L. Manipulating host resistance structure reveals impact of pathogen dispersal and environmental heterogeneity on epidemics. Ecology 2018, 99, 2853–2863. [Google Scholar] [CrossRef] [Green Version]
- McCallum, H.; Barlow, N.; Hone, J. How should pathogen transmission be modelled? Trends Ecol. Evol. 2001, 16, 295–300. [Google Scholar] [CrossRef]
- Civitello, D.J.; Cohen, J.; Fatima, H.; Halstead, N.T.; Liriano, J.; McMahon, T.A.; Ortega, C.N.; Sauer, E.L.; Sehgal, T.; Young, S.; et al. Biodiversity inhibits parasites: Broad evidence for the dilution effect. Proc. Natl. Acad. Sci. USA 2015, 112, 8667–8671. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.T.J.; Thieltges, D.W. Diversity, decoys and the dilution effect: How ecological communities affect disease risk. J. Exp. Biol. 2010, 213, 961–970. [Google Scholar] [CrossRef] [Green Version]
- Louhi, K.-R.; Sundberg, L.-R.; Jokela, J.; Karvonen, A. Interactions among bacterial strains and fluke genotypes shape virulence of co-infection. Proc. R. Soc. B 2015, 282, 20152097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karvonen, A.; Jokela, J.; Laine, A.-L. Importance of sequence and timing in parasite coinfections. Trends Parasitol. 2015, 35, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Tollenaere, C.; Susi, H.; Laine, A.-L. Evolutionary and epidemiological implications of multiple infection in plants. Trends Plant. Sci. 2016, 21, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; De Roode, J.C.; Fenton, A. Why infectious disease research needs community ecology. Science 2015, 349, 1259504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddicoat, C.; Weinstein, P.; Bissett, A.; Genie, N.J.C.; Mills, J.G.; Waycott, M.; Breed, M.F. Can bacterial indicators of a grassy woodland restoration inform ecosystem assessment and microbiota-mediated human health? Environ. Int. 2019, 129, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Roalkvam, I.; Dronen, K.; Dahle, H.; Wergeland, H.I. Microbial communities in a flow-through fish farm for lumpfish (Cyclopterus lumpus L.) during healthy rearing conditions. Front. Microbiol. 2019, 10, 1594. [Google Scholar] [CrossRef] [Green Version]
- Conlon, M.A.; Bird, A.R. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2015, 7, 17–44. [Google Scholar] [CrossRef]
- Harris, E.V.; de Roode, J.C.; Gerardo, N.M. Diet-microbiome-disease: Investigating diet’s influence on infectious disease resistance through alteration of the gut microbiome. PLoS Pathog. 2019, 15, e1007891. [Google Scholar] [CrossRef]
- Villarino, N.F.; LeCleir, G.R.; Denny, J.E.; Dearth, S.P.; Harding, C.L.; Sloan, S.S.; Gribble, J.L.; Campagna, S.R.; Wilhelm, S.W.; Schmidt, N.W. Composition of the gut microbiota modulates the severity of malaria. Proc. Natl. Acad. Sci. USA 2016, 113, 2235–2240. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.N.; Yang, Y.Z.; Cao, L.; Yin, H.Q.; Xu, M.Y.; Wang, Z.J.; Liu, Y.Y.; Wang, X.; Deng, Y. Habitat environments impacted the gut microbiome of long-distance migratory swan geese but central species conserved. Sci. Rep. 2018, 8, 13314. [Google Scholar] [CrossRef]
- De Assis, A.B.; Barreto, C.C.; Navas, C.A. Skin microbiota in frogs from the Brazilian Atlantic Forest: Species, forest type, and potential against pathogens. PLoS ONE 2017, 12, e0179628. [Google Scholar] [CrossRef] [Green Version]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A.M. Environmental and physiological factors shape the gut microbiota of Atlantic salmon parr (Salmo salar L.). Aquaculture 2017, 467, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Jani, A.J.; Briggs, C.J. Host and aquatic environment shape the amphibian skin microbiome but effects on downstream resistance to the pathogen Batrachochytrium dendrobatidis are variable. Front. Microbiol. 2018, 9, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasemägi, A.; Visse, M.; Kisand, V. Effect of environmental factors and an emerging parasitic disease on gut microbiome of wild salmonid fish. MSphere 2017, 2, e00418-17. [Google Scholar] [CrossRef] [Green Version]
- Xavier, R.; Mazzei, R.; Perez-Losada, M.; Rosado, D.; Santos, J.L.; Verissimo, A.; Soares, M.C. A risky business? Habitat and social behavior impact skin and gut microbiomes in Caribbean cleaning gobies. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karvonen, A.; Aalto-Araneda, M.; Virtala, A.M.; Kortet, R.; Koski, P.; Hyvärinen, P. Enriched rearing environment and wild genetic background can enhance survival and disease resistance of salmonid fishes during parasite epidemics. J. Appl. Ecol. 2016, 53, 213–221. [Google Scholar] [CrossRef]
- Räihä, V.; Sundberg, L.-R.; Ashrafi, R.; Hyvärinen, P.; Karvonen, A. Rearing background and exposure environment together explain higher survival of aquaculture fish during a bacterial outbreak. J. Appl. Ecol. 2019, 56, 1741–1750. [Google Scholar] [CrossRef]
- Suomalainen, L.-R.; Kunttu, H.; Valtonen, E.T.; Hirvelä-Koski, V.; Tiirola, M. Molecular diversity and growth features of Flavobacterium columnare strains isolated in Finland. Dis. Aquat. Org. 2006, 70, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Declercq, A.M.; Haesebrouck, F.; Van den Broeck, W.; Bossier, P.; Decostere, A. Columnaris disease in fish: A review with emphasis on bacterium-host interactions. Vet. Res. 2013, 44, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulkkinen, K.; Suomalainen, L.-R.; Read, A.F.; Ebert, D.; Rintamäki, P.; Valtonen, E.T. Intensive fish farming and the evolution of pathogen virulence: The case of columnaris disease in Finland. Proc. R. Soc. B 2010, 277, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef]
- Bottery, M.J.; Pitchford, J.W.; Friman, V.P. Ecology and evolution of antimicrobial resistance in bacterial communities. ISME J. 2020, 1–10. [Google Scholar] [CrossRef]
- Sundberg, L.-R.; Karvonen, A. Minor environmental concentrations of antibiotics can modify bacterial virulence in co-infection with a non-targeted parasite. Biol. Lett. 2018, 14, 20180663. [Google Scholar] [CrossRef] [Green Version]
- Rodewald, P.; Hyvärinen, P.; Hirvonen, H. Wild origin and enriched environment promote foraging rate and learning to forage on natural prey of captive reared Atlantic salmon parr. Ecol. Freshw. Fish 2011, 20, 569–579. [Google Scholar] [CrossRef]
- Näslund, J.; Rosengren, M.; Del Villar, D.; Gansel, L.; Norrgard, J.R.; Persson, L.; Winkowski, J.J.; Kvingedal, E. Hatchery tank enrichment affects cortisol levels and shelter-seeking in Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 2013, 70, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Strand, D.A.; Utne-Palm, A.C.; Jakobsen, P.J.; Braithwaite, V.A.; Jensen, K.H.; Salvanes, A.G.V. Enrichment promotes learning in fish. Mar. Ecol. Prog. Ser. 2010, 412, 273–282. [Google Scholar] [CrossRef]
- Salvanes, A.G.V.; Moberg, O.; Ebbesson, L.O.E.; Nilsen, T.O.; Jensen, K.H.; Braithwaite, V.A. Environmental enrichment promotes neural plasticity and cognitive ability in fish. Proc. R. Soc. B 2013, 280, 20131331. [Google Scholar] [CrossRef] [PubMed]
- Braithwaite, V.A.; Salvanes, A.G.V. Environmental variability in the early rearing environment generates behaviourally flexible cod: Implications for rehabilitating wild populations. Proc. R. Soc. B 2005, 272, 1107–1113. [Google Scholar] [CrossRef] [Green Version]
- Salvanes, A.G.V.; Braithwaite, V.A. Exposure to variable spatial information in the early rearing environment generates asymmetries in social interactions in cod (Gadus morhua). Behav. Ecol. Sociobiol. 2005, 59, 250–257. [Google Scholar] [CrossRef]
- Hyvärinen, P.; Rodewald, P. Enriched rearing improves survival of hatchery-reared Atlantic salmon smolts during migration in the River Tornionjoki. Can. J. Fish Aquat. Sci. 2013, 70, 1386–1395. [Google Scholar] [CrossRef]
- Makino, H.; Masuda, R.; Tanaka, M. Environmental stimuli improve learning capability in striped knifejaw juveniles: The stage-specific effect of environmental enrichment and the comparison between wild and hatchery-reared fish. Fish Sci. 2015, 81, 1035–1042. [Google Scholar] [CrossRef] [Green Version]
- Suomalainen, L.-R.; Tiirola, M.A.; Valtonen, E.T. Effect of Pseudomonas sp MT5 baths on Flavobacterium columnare infection of rainbow trout and on microbial diversity on fish skin and gills. Dis. Aquat. Org. 2005, 63, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Tiirola, M.; Valtonen, E.T.; Rintamaki-Kinnunen, P.; Kulomaa, M.S. Diagnosis of flavobacteriosis by direct amplification of rRNA genes. Dis. Aquat. Org. 2002, 51, 93–100. [Google Scholar] [CrossRef]
- Chowdhury, M.B.R.; Wakabayashi, H. Effects of competitive bacteria on the survival and infectivity of Flexibacter columnaris. Fish Pathol. 1989, 24, 9–15. [Google Scholar] [CrossRef]
- Kunttu, H.M.T.; Valtonen, E.T.; Jokinen, E.I.; Suomalainen, L.-R. Saprophytism of a fish pathogen as a transmission strategy. Epidemics 2009, 1, 96–100. [Google Scholar] [CrossRef]
- Magnadottir, B. Innate immunity of fish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef]
- Tort, L. Stress and immune modulation in fish. Dev. Comparative Immunol. 2011, 35, 1366–1375. [Google Scholar] [CrossRef]
- Kortet, R.; Taskinen, J.; Sinisalo, T.; Jokinen, I. Breeding-related seasonal changes in immunocompetence, health state and condition of the cyprinid fish, Rutilus rutilus, L. Biol. J. Linn. Soc. 2003, 78, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Bartolomucci, A. Social stress, immune functions and disease in rodents. Front. Neuroendocr. 2007, 28, 28–49. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Holt, R.D. How should environmental stress affect the population dynamics of disease? Ecol. Lett. 2003, 6, 654–664. [Google Scholar] [CrossRef] [Green Version]
- Leicht, K.; Jokela, J.; Seppälä, O. An experimental heat wave changes immune defense and life history traits in a freshwater snail. Ecol. Evol. 2013, 3, 4861–4871. [Google Scholar] [CrossRef]
- Jokela, J.; Taskinen, J.; Mutikainen, P.; Kopp, K. Virulence of parasites in hosts under environmental stress: Experiments with anoxia and starvation. Oikos 2005, 108, 156–164. [Google Scholar] [CrossRef]
- Klemme, I.; Hendrikx, L.; Ashrafi, R.; Sundberg, L.-R.; Räihä, V.; Piironen, J.; Hyvärinen, P.; Karvonen, A. Opposing health effects of hybridization for conservation. Conserv. Sci. Pract. 2021, e379. [Google Scholar] [CrossRef]
- Sundberg, L.-R.; Ketola, T.; Laanto, E.; Kinnula, H.; Bamford, J.K.H.; Penttinen, R.; Mappes, J. Intensive aquaculture selects for increased virulence and interference competition in bacteria. Proc. R. Soc. B 2016, 283, 20153069. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.L.; Fryer, J.L.; Rohovec, J.S. Comparison of six media for the cultivation of Flexibacter columnaris. Fish Pathol. 1988, 23, 91–94. [Google Scholar] [CrossRef] [Green Version]
- Decostere, A.; Haesebrouck, F.; Devriese, L.A. Shieh medium supplemented with tobramycin for selective isolation of Flavobacterium columnare (Flexibacter columnaris) from diseased fish. J. Clin. Microbiol. 1997, 35, 322–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Source | χ2 | Degrees of Freedom | p |
|---|---|---|---|
| Quantity of stones | 18.043 | 1 | <0.001 |
| Quality of stones | 4.333 | 1 | 0.037 |
| Quantity×Quality | 4.933 | 1 | 0.026 |
| Fish length | 0.067 | 1 | 0.796 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karvonen, A.; Räihä, V.; Klemme, I.; Ashrafi, R.; Hyvärinen, P.; Sundberg, L.-R. Quantity and Quality of Aquaculture Enrichments Influence Disease Epidemics and Provide Ecological Alternatives to Antibiotics. Antibiotics 2021, 10, 335. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030335
Karvonen A, Räihä V, Klemme I, Ashrafi R, Hyvärinen P, Sundberg L-R. Quantity and Quality of Aquaculture Enrichments Influence Disease Epidemics and Provide Ecological Alternatives to Antibiotics. Antibiotics. 2021; 10(3):335. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030335
Chicago/Turabian StyleKarvonen, Anssi, Ville Räihä, Ines Klemme, Roghaieh Ashrafi, Pekka Hyvärinen, and Lotta-Riina Sundberg. 2021. "Quantity and Quality of Aquaculture Enrichments Influence Disease Epidemics and Provide Ecological Alternatives to Antibiotics" Antibiotics 10, no. 3: 335. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030335