
Repurposing of the Tamoxifen Metabolites to Combat Infections by Multidrug-Resistant Gram-Negative Bacilli

 and
and
Abstract
:1. Introduction
2. Results
2.1. Tamoxifen Increases Mice Survival and Decreases the Bacterial Burden in Immunosuppressed Mice
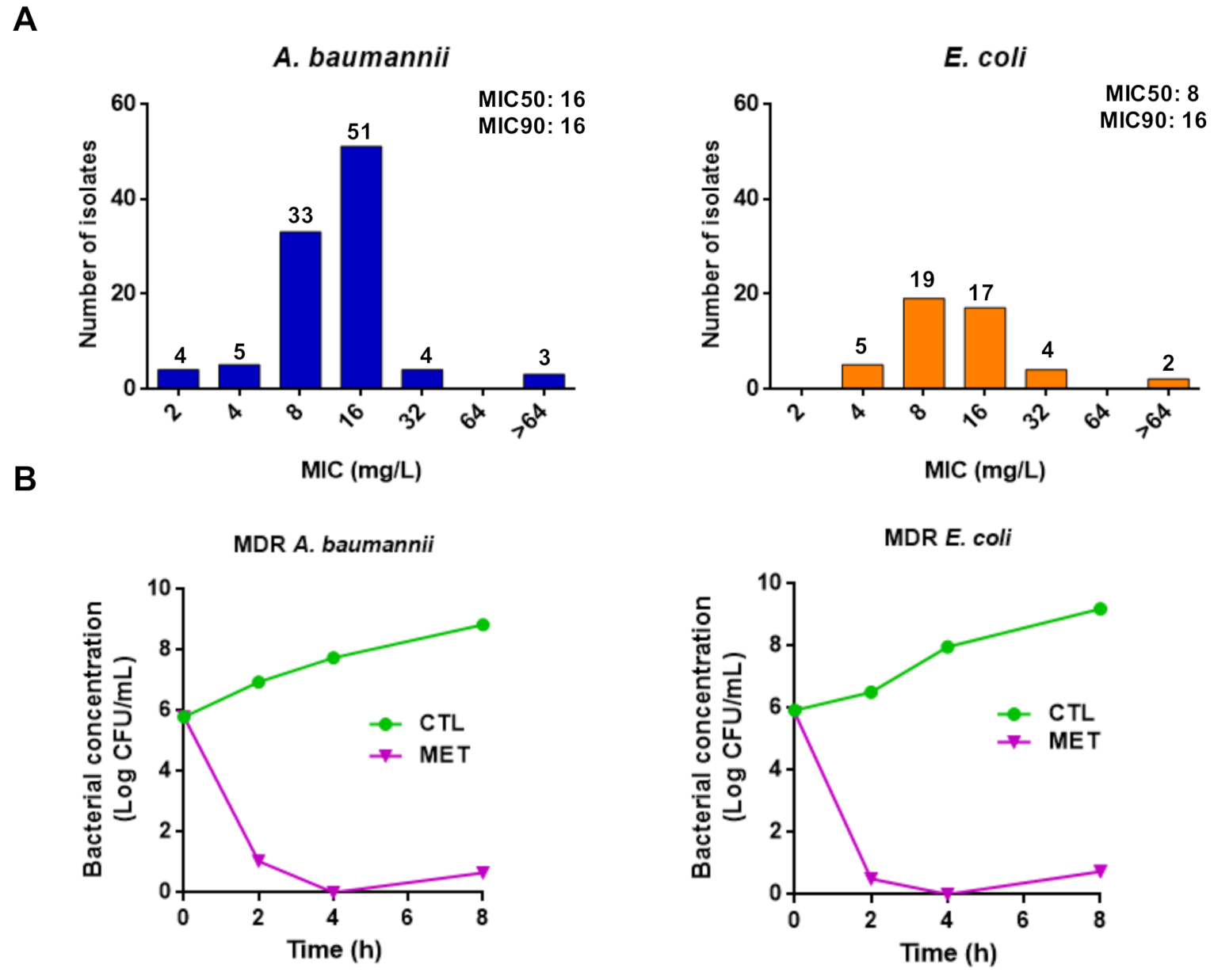
2.2. Antibacterial Activity of Tamoxifen Metabolites
2.3. Effect of Tamoxifen Metabolites on the Bacterial Cell Membrane
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Bacterial Strains
4.3. Animals
4.4. Immunosuppressed Mice
4.5. Therapeutic Effect of Tamoxifen in Immunocompetent Murine Models of Peritoneal Sepsis
4.6. Therapeutic Effect of Tamoxifen in Immunosuppressed Murine Models of Peritoneal Sepsis
4.7. In Vitro Susceptibility Testing and Time-Kill Experiments
4.8. Analysis of Outer Membrane Proteins (OMPs) by SDS–PAGE
4.9. Membrane Permeability Assays
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miró-Canturri, A.; Ayerbe-Algaba, R.; del Toro, R.; Pachón, J.; Smani, Y. Tamoxifen repurposing to combat infections by multidrug-resistant Gram-negative bacilli. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, A.C.; Didone, L.; Jobson, J.; Sofia, M.K.; Krysan, D.; Dunman, P.M. Adenylate kinase release as a high-throughput-screening-compatible reporter of bacterial lysis for identification of antibacterial agents. Antimicrob. Agents Chemother. 2013, 57, 26–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farha, M.A.; Czarny, T.L.; Myers, C.L.; Worrall, L.J.; French, S.; Conrady, D.G.; Wang, Y.; Oldfield, E.; Strynadka, N.C.J.; Brown, E.D. Antagonism screen for inhibitors of bacterial cell wall biogenesis uncovers an inhibitor of undecaprenyl diphosphate synthase. Proc. Natl. Acad. Sci. USA 2015, 112, 11048–11053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corriden, R.; Hollands, A.; Olson, J.; Derieux, J.; Lopez, J.; Chang, J.T.; Gonzalez, D.J.; Nizet, V. Tamoxifen augments the innate immune function of neutrophils through modulation of intracellular ceramide. Nat. Commun. 2015, 6, 8369. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoskins, J.M.; Carey, L.A.; McLeod, H.L. CYP2D6 and tamoxifen: DNA matters in breast cancer. Nat. Rev. Cancer 2009, 9, 576–586. [Google Scholar] [CrossRef]
- Luo, G.; Spellberg, B.; Gebremariam, T.; Bolaris, M.; Lee, H.; Fu, Y.; French, S.W.; Ibrahim, A.S. Diabetic murine models for Acinetobacter baumannii infection. J. Antimicrob. Chemother. 2012, 67, 1439–1445. [Google Scholar] [CrossRef] [Green Version]
- Pletzer, D.; Mansour, S.C.; Wuerth, K.; Rahanjam, N.; Hancock, R.E. New mouse model for chronic infections by Gram-negative bacteria enabling the study of anti-infective efficacy and host-microbe interactions. mBio 2017, 8, e00140-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuluaga, A.F.; Salazar, B.E.; Rodríguez, C.A.; Zapata, A.X.; Agudelo, M.; Vesga, O. Neutropenia induced in outbred mice by a simplified low-dose cyclophosphamide regimen: Characterization and applicability to diverse experimental models of infectious diseases. BMC Infect. Dis. 2006, 6, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, R.E.W.; Brinkman, F.S.L. Function of Pseudomonas porin in uptake and efflux. Ann. Rev. Microbiol. 2002, 56, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Mesaros, N.; Nordmann, P.; Plésiat, P.; Roussel-Delvallez, M.; Eldere, J.V.; Glupczynski, Y.; Van Laethem, Y.; Jacobs, F.; Lebecque, P.; Malfroot, A.; et al. Pseudomonas aeruginosa: Resistance and therapeutic options at the turn of the new millennium. Clin. Microbiol. Infect. 2007, 13, 560–578. [Google Scholar] [CrossRef] [Green Version]
- Montoya, M.C.; Krysan, D.J. Repurposing estrogen receptor antagonists for the treatment of infectious disease. mBio 2018, 9, e02272-18. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.C.; Liao, Y.C.; Huang, J.M.; Lin, C.H.; Chen, Y.Y.; Dou, H.Y.; Hsiung, C.A. Pros and cons of the tuberculosis drugome approach-an empirical analysis. PLoS ONE 2014, 9, e100829. [Google Scholar] [CrossRef]
- Weinstock, A.; Gallego-Delgado, J.; Gomes, C.; Sherman, J.; Nikain, C.; Gonzalez, S.; Fisher, E.; Rodriguez, A. Tamoxifen activity against Plasmodium in vitro and in mice. Malar. J. 2019, 18, 378. [Google Scholar] [CrossRef] [PubMed]
- Butts, A.; Koselny, K.; Chabrier-Rosello, Y.; Semighini, C.P.; Brown, J.C.; Wang, X.; Annadurai, S.; DiDone, L.; Tabroff, J.; Childers, W.E., Jr.; et al. Estrogen receptor antagonists are anti-cryptococcal agents that directly bind EF hand proteins and synergize with fluconazole in vivo. mBio 2014, 5, e00765-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolan, K.; Montgomery, S.; Buchheit, B.; DiDone, L.; Wellington, M.; Krysan, D.J. Antifungal activity of tamoxifen: In vitro and in vivo activities and mechanistic characterization. Antimicrob. Agents Chemother. 2009, 53, 3337–3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butts, A.; Martin, J.A.; DiDone, L.; Bradley, E.K.; Mutz, M.; Krysan, J. Structure-activity relationships for the antifungal activity of selective estrogen receptor antagonists related to tamoxifen. PLoS ONE 2015, 10, e0125927. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.A.; Spencer, C.T.; O’Reilly, M.C.; Browb, K.A.; Lavieri, R.R.; Cho, C.H.; Jung, D.I.; Larock, R.C.; Brown, H.A.; Lindsley, C.W. Discovery of desketoraloxifene analogues as inhibitors of mammalian, Pseudomonas aeruginosa, and nape phospholipase d enzymes. ACS Chem. Biol. 2015, 10, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Baumann, P.; Doudoroff, M.; Stanier, M.R. A 377 study of the Moraxella grouP. II. Oxidative negative species (genus Acinetobacter). J. Bacteriol. 1968, 95, 1520–1541. [Google Scholar] [CrossRef] [Green Version]
- Holloway, I.W. Genetic recombination in Pseudomonas aeruginosa. J. Gen. Microbial. 1955, 13, 572–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escherichia coli (Migula) Castellani and Chalmers (ATCC® 25922™). In 1946 FDA strain Seattle [DSM 1103, NCIB 12210]; American Type Culture Collection: Manassas, VA, USA, 2021.
- Parra Millán, R.; Jiménez Mejías, M.E.; Sánchez Encinales, V.; Ayerbe Algaba, R.; Gutiérrez Valencia, A.; Pachón Ibáñez, M.E.; Díaz, C.; Pérez Del Palacio, J.; López Cortés, L.F.; Pachón, J.; et al. Efficacy of lysophosphatidylcholine in combination with antimicrobial agents against Acinetobacter baumannii in experimental murine peritoneal sepsis and pneumonia models. Antimicrob. Agents Chemother. 2016, 60, 4464–4470. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Cuenca, F.; Tomás-Carmona, M.; Caballero-Moyano, F.; Bou, G.; Martínez-Martínez, L.; Vila, J.; Pachón, J.; Cisneros, J.M.; Rodríguez-Baño, J.; Pascual, A. In vitro activity of 18 antimicrobial agents against clinical isolates of Acinetobacter spp.: Multicenter national study GEIH-REIPI-Ab 2010. Enferm. Infecc. Microbiol. Clin. 2012, 31, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Yanat, B.; Machuca, J.; Yahia, R.D.; Touati, A.; Pascual, Á.; Rodríguez-Martínez, J.M. First report of the plasmid-mediated colistin resistance gene mcr-1 in a clinical Escherichia coli isolate in Algeria. Int. J. Antimicrob. Agents 2016, 48, 760–761. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Villodres, A.; Alvarez Marín, R.; Durán Lobato, M.; Pachón Ibáñez, M.E.; Aznar, J.; Pachón Díaz, J.; Lepe, J.A.; Smani, Y. Overexpression of outer membrane protein A by Escherichia coli as a risk factor for mortality in bloodstream infection. In Proceedings of the 29th European Congress of Clinical Microbiology and Infectious Diseases, Amsterdam, The Netherlands, 13–16 April 2019. [Google Scholar]
- Vila-Farrés, X.; Parra-Millán, R.; Sánchez-Encinales, V.; Varese, M.; Ayerbe-Algaba, R.; Bayó, N.; Guardiola, S.; Pachón-Ibáñez, M.E.; Kotev, M.; García, J.; et al. Combating virulence of Gram-negative bacilli by OmpA inhibition. Sci. Rep. 2017, 7, 14683. [Google Scholar] [CrossRef] [Green Version]
- Jackson Website. Available online: https://www.jax.org/research-and-faculty/resources/cre-repository/tamoxifen# (accessed on 3 February 2017).
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing (Twenty-Seventh Informational Supplement M100-S27; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Smani, Y.; Docobo-Pérez, F.; López-Rojas, R.; Domínguez-Herrera, J.; Ibáñez-Martínez, J.; Pachón, J. Platelet-activating factor receptor initiates contact of Acinetobacter baumannii expressing phosphorylcholine with host cells. J. Biol. Chem. 2012, 287, 26901–26910. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Strain | Treatment | n | Bacterial Load (log CFU/g or mL ± SEM) | Survival (%) | ||
|---|---|---|---|---|---|---|
| Spleen | Lung | Blood | ||||
| A. baumannii ATCC 17978 | CTL | 6 | 9.51 ± 0.17 | 9.77 ± 0.17 | 6.14 ± 0.94 | 0 |
| CPM | 6 | 10.06 ± 0.24 | 9.91 ± 0.25 | 8.32 ± 0.25 a | 0 | |
| TAM | 6 | 2.87 ± 1.21 | 2.61 ± 1.07 | 0.61 ± 0.61 | 100 | |
| CPM + TAM | 6 | 3.14 ± 1.25c | 3.46 ± 1.31c | 2.33 ± 1.00 c | 66.7 | |
| P. aeruginosa PAO1 | CTL | 5 | 8.91 ± 0.15 | 9.24 ± 0.17 | 6.71 ± 0.27 | 0 |
| CPM | 6 | 9.94 ± 0.06 a | 9.36 ± 0.13 | 7.09 ± 0.28 | 0 | |
| TAM | 6 | 5.33 ± 1.08 | 4.14 ± 1.50 | 1.26 ± 1.26 | 66.7 | |
| CPM + TAM | 6 | 8.71 ± 0.94 b | 8.77 ± 0.83 b | 4.04 ± 0.82 b,c | 16.7 | |
| E. coli ATCC 25922 | CTL | 6 | 8.71 ± 0.05 | 8.88 ± 0.16 | 8.18 ± 0.37 | 0 |
| CPM | 6 | 10.55 ± 0.13 a | 9.92 ± 0.19 a | 6.92 ± 0.38a | 0 | |
| TAM | 6 | 5.01 ± 1.20 | 4.72 ± 1.08 | 3.87 ± 0.99 | 83.3 | |
| CPM + TAM | 6 | 5.93 ± 1.22 c | 5.92 ± 1.00 c | 3.95 ± 1.22 c | 66.7 | |
| Strain | TAM (mg/L) | DTAM (mg/L) | ENDX (mg/L) | HTAM (mg/L) | MET (mg/L) |
|---|---|---|---|---|---|
| A. baumannii ATCC 17978 | >256 | 8 | 16 | 16 | 8 |
| A. baumannii Ab186 | >256 | 16 | 16 | 32 | 4 |
| P. aeruginosa PAO1 | >256 | ≥64 | ≥64 | ≥64 | ≥64 |
| E. coli ATCC 25922 | >256 | 8 | 16 | 32 | 8 |
| E. coli EcMCR+ | >256 | ≥64 | 64 | ≥64 | 32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miró-Canturri, A.; Ayerbe-Algaba, R.; Vila-Domínguez, A.; Jiménez-Mejías, M.E.; Pachón, J.; Smani, Y. Repurposing of the Tamoxifen Metabolites to Combat Infections by Multidrug-Resistant Gram-Negative Bacilli. Antibiotics 2021, 10, 336. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030336
Miró-Canturri A, Ayerbe-Algaba R, Vila-Domínguez A, Jiménez-Mejías ME, Pachón J, Smani Y. Repurposing of the Tamoxifen Metabolites to Combat Infections by Multidrug-Resistant Gram-Negative Bacilli. Antibiotics. 2021; 10(3):336. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030336
Chicago/Turabian StyleMiró-Canturri, Andrea, Rafael Ayerbe-Algaba, Andrea Vila-Domínguez, Manuel E. Jiménez-Mejías, Jerónimo Pachón, and Younes Smani. 2021. "Repurposing of the Tamoxifen Metabolites to Combat Infections by Multidrug-Resistant Gram-Negative Bacilli" Antibiotics 10, no. 3: 336. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030336