
Prevalence and Characterization of Quinolone-Resistance Determinants in Escherichia coli Isolated from Food-Producing Animals and Animal-Derived Food in the Philippines



Abstract
:1. Introduction
2. Results
2.1. Prevalence of E. coli in Samples
2.2. Antimicrobial Susceptibility
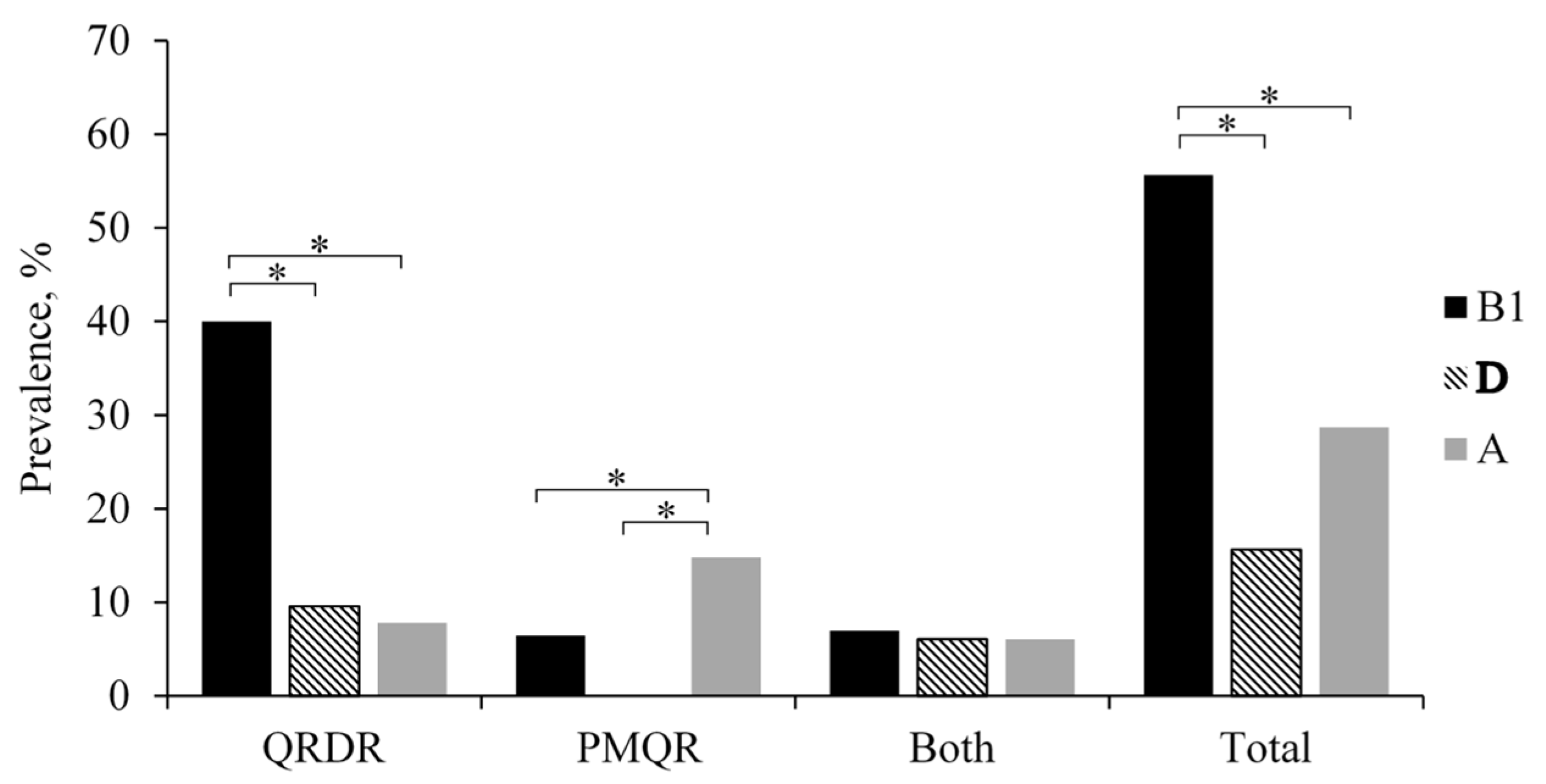
2.3. QRDR and PMQR Determinant Analysis
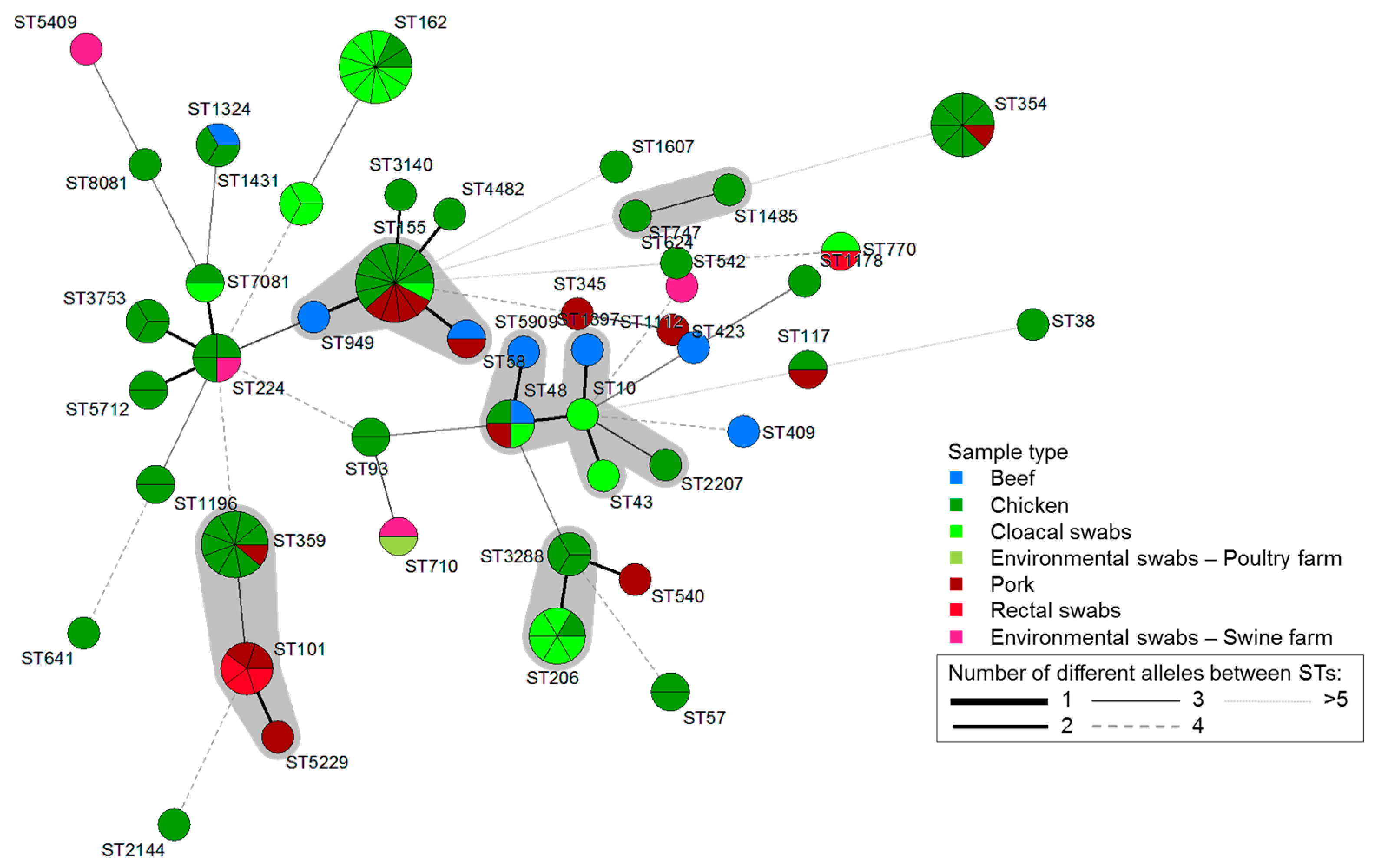
2.4. Multilocus Sequence Typing and Phylogenetic Group Analysis
2.5. Prevalence of Virulence Genes
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Bacterial Enrichment, and Isolation
4.2. Antimicrobial Susceptibility Testing
4.3. Detection of Quinolone-Resistance Determinants
4.4. Multilocus Sequence Typing Analysis
4.5. Detection of Virulence Genes
4.6. Phylogenetic Group Analysis
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- North, J. Challenges to Tackling Antimicrobial Resistance: Economic and Policy Responses; Anderson, M., Cecchini, M., Mossialos, E., Eds.; European Observatory on Health Systems and Policies; Cambridge University Press: Cambridge, UK, 2020; Volume 53. [Google Scholar] [CrossRef]
- Food and Agriculture of the United Nations (FAO). The FAO Action Plan on Antimicrobial Resistance, 2016–2020: Supporting the Food and Agriculture Sectors in Implementing the Global Action Plan on Antimicrobial Resistance to Minimize the Impact of Antimicrobial Resistance; FAO: Rome, Italy, 2015. [Google Scholar]
- Todd, E. Food-borne disease prevention and risk assessment. Int. J. Environ. Res. Public Health 2020, 17, 5129. [Google Scholar] [CrossRef]
- Hao, H.; Sander, P.; Iqbal, Z.; Wang, Y.; Cheng, G.; Yuan, Z. The risk of some veterinary antimicrobial agents on public health associated with antimicrobial resistance and their molecular basis. Front. Microbiol. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.C.; Turnidge, J.; Collignon, P.; Looke, D.; Barton, M.; Gottlieb, T. Control of fluoroquinolone resistance through successful regulation, Australia. Emerg. Infect. Dis. 2012, 18, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.W.; Lee, Y.J. Characterization of plasmid mediated quinolone resistance determinants in ciprofloxacin resistant-Escherichia coli from chicken meat produced by integrated broiler operations in Korea. Int. J. Food Microbiol. 2019, 307, 108274. [Google Scholar] [CrossRef]
- Röderova, M.; Halova, D.; Papousek, I.; Dolejska, M.; Masarikova, M.; Hanulik, V.; Pudova, V.; Broz, P.; Htoutou-Sedlakova, M.; Sauer, P.; et al. Characteristics of Quinolone Resistance in Escherichia coli Isolates from Humans, Animals, and the Environment in the Czech Republic. Front. Microbiol. 2017, 7, 2147. [Google Scholar] [CrossRef] [Green Version]
- Aldred, K.J.; Kerns, R.J.; Osheroff, N. Mechanism of Quinolone Action and Resistance. Biochemistry 2014, 53, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.; Poeta, P.; Hébraud, M.; Capelo, J.L.; Igrejas, G. Mechanisms of quinolone action and resistance: Where do we stand? J. Med. Microbiol. 2017, 66, 551–559. [Google Scholar] [CrossRef]
- Poirel, L.; Cattoir, V.; Nordmann, P. Plasmid-mediated quinolone resistance; interactions between human, animal, and environmental ecologies. Front. Microbiol. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Antimicrob. Resist. Bact. Livest. Companion Anim. 2018, 6, 289–316. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Zeng, Z.; Rao, L.; Chen, X.; He, D.; Lv, L.; Wang, J.; Zeng, L.; Feng, M.; Liu, J.H. The association between occurrence of plasmid-mediated quinolone resistance and ciprofloxacin resistance in Escherichia coli isolates of different origins. Vet. Microbiol. 2014, 170, 89–96. [Google Scholar] [CrossRef]
- Jiao, S.C.; Fami, R.M.L.; Pedernal, V.A.D.; Cabrera, E.C. Prevalence of multiple drug-resistant Escherichia coli from chicken, pig and nile tilapia (Oreochromis nilotica) intestines sold in wet markets in Manila and the conjugative transferability of the resistance. Philipp. Agric. Sci. 2007, 90, 64–70. [Google Scholar]
- Padilla, M.A.; Amatorio, M.Q. High level resistance and multi-resistance to medically important antimicrobials in Escherichia coli Isolated from healthy pigs at slaughter in Laguna, Philippines. Philipp. J. Vet. Med. 2017, 54, 36–45. [Google Scholar]
- Torio, H.E.; Padilla, M.A. Multiple resistance to medically important antimicrobials of commensal Escherichia coli isolated from dressed broiler chickens in Calabarzon, Philippines. Philipp. J. Vet. Med. 2018, 55, 95–106. [Google Scholar]
- Gundran, R.S.; Cardenio, P.A.; Villanueva, M.A.; Sison, F.B.; Benigno, C.C.; Kreausukon, K.; Pichpol, D.; Punyapornwithaya, V. Prevalence and distribution of bla CTX-M, bla SHV, bla TEM genes in extended- spectrum β- Lactamase- producing E. coli isolates from broiler farms in the Philippines. BMC Vet. Res. 2019, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gundran, R.S.; Cardenio, P.A.; Salvador, R.T.; Sison, F.B.; Benigno, C.C.; Kreausukon, K.; Pichpol, D.; Punyapornwithaya, V. Prevalence, Antibiogram, and Resistance Profile of Extended-Spectrum β-Lactamase-Producing Escherichia coli Isolates from Pig Farms in Luzon, Philippines. Microb. Drug Resist. 2020, 26, 160–168. [Google Scholar] [CrossRef]
- Paraoan, C.E.M.; Rivera, W.L.; Vital, P.G. Detection of Class I and II integrons for the assessment of antibiotic and multidrug resistance among Escherichia coli isolates from agricultural irrigation waters in Bulacan, Philippines. J. Environ. Sci. Health Part B 2017, 52, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Vital, P.G.; Caballes, M.B.D.; Rivera, W.L. Antimicrobial resistance in escherichia coli and salmonella spp. Isolates from fresh produce and the impact to food safety. J. Environ. Sci. Health Part B 2017, 52, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Department of Health (DOH). Health Research Brief: Combatting Stubborn Pathogens: Strengthening Surveillance on Antimicrobial Resistance; Department of Health (DOH), Health Policy Development and Planning Bureau: Manila, Philippines, 2017; Volume 3. Available online: https://doh.gov.ph/node/10641 (accessed on 5 February 2019).
- World Health Organization (WHO). Integrated Surveillance of Antimicrobial Resistance in Foodborne Bacteria: Application of a One Health Approach; World Health Organization (WHO): Geneva, Switzerland, 2017. [Google Scholar]
- Elumba, Z.S.; Allera, M.L.M.; Taganas, R.R.R. Occurrence and Antibiotic Sensitivity of Escherichia coli and Salmonella spp. in Retail Chicken Meat at Selected Markets in Valencia City, Bukidnon, Philippines. Asian J. Biol. Life Sci. 2018, 7, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Jiang, X.; Fu, K.; Liu, B.; Xu, D.; Ji, S.; Zhou, L. Detection of Extended-Spectrum β-Lactamase and Plasmid-Mediated Quinolone Resistance Determinants in Escherichia coli Isolates from Retail Meat in China. J. Food Sci. 2015, 80, M1039–M1043. [Google Scholar] [CrossRef]
- Rahman, M.A.; Rahman, A.K.M.A.; Islam, M.A.; Alam, M.M. Antimicrobial Resistance of Escherichia Coli Isolated From Milk, Beef And Chicken Meat in Bangladesh. Bangladesh J. Vet. Med. 2017, 15, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Azanza, M.P.V.; Membrebe, B.N.Q.; Sanchez, R.G.R.; Estilo, E.E.C.; Dollete, U.G.M.; Feliciano, R.J.; Garcia, N.K.A. Foodborne disease outbreaks in the Philippines (2005–2018). Philipp. J. Sci. 2019, 148, 317–336. [Google Scholar]
- Barroga, T.R.M.; Morales, R.G.; Benigno, C.C.; Castro, S.J.M.; Caniban, M.M.; Cabullo, M.F.B.; Agunos, A.; de Balogh, K.; Dorado-Garcia, A. Antimicrobials Used in Backyard and Commercial Poultry and Swine Farms in the Philippines: A Qualitative Pilot Study. Front. Vet. Sci. 2020, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Department of Agriculture (DA) and Department of Health (DOH). Declaring a Ban on the Use of Chloramphenicol in Food Producing Animals: Department of Agriculture (DA) Administrative Order No. 60, Series of 1990, Department of Health (DOH) Administrative Order No. 91, Series of 1990; Department of Agriculture (DA) and Department of Health (DOH): Manila, Philipines, 1990. Available online: https://ww2.fda.gov.ph/attachments/article/155173/AO%2091%20Ban%20(Chloramphenicol%20in%20Animals).pdf (accessed on 10 December 2020).
- Trongjit, S.; Angkittitrakul, S.; Chuanchuen, R. Occurrence and molecular characteristics of antimicrobial resistance of Escherichia coli from broilers, pigs and meat products in Thailand and Cambodia provinces. Microbiol. Immunol. 2016, 60, 575–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Department of Health (DOH). Antimicrobial Resistance Surveillance Program Annual Report—2018; Research Institute for Tropical Medicine, Antimicrobial Resistance Surveillance Reference Laboratory: Manila, Philipines, 2019; Available online: https://arsp.com.ph/arsp-2018-annual-report-data-summary-is-now-available-for-download/ (accessed on 6 September 2019).
- Mølbak, K. Human health consequences of antimicrobial drug-resistant Salmonella and other foodborne pathogens. Clin. Infect. Dis. 2005, 41, 1613–1620. [Google Scholar] [CrossRef]
- VT Nair, D.; Venkitanarayanan, K.; Kollanoor Johny, A. Antibiotic-resistant Salmonella in the food supply and the potential role of antibiotic alternatives for control. Foods 2018, 7, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, K.L.; Davies, R.H.; Threlfall, E.J. Mechanisms of quinolone resistance in Escherichia coli and Salmonella: Recent developments. Int. J. Antimicrob. Agents 2005, 25, 358–373. [Google Scholar] [CrossRef]
- World Health Oorganization (WHO). Critically Important Antimicrobials for Human Medicine; 6th revision; World Health Organization (WHO): Geneva, Switzerland, 2019. [Google Scholar]
- Tamang, M.D.; Nam, H.M.; Chae, M.H.; Kim, S.R.; Gurung, M.; Jang, G.C.; Jung, S.C.; Lim, S.K. Prevalence of plasmid-mediated quinolone resistance determinants among escherichia coli isolated from food animals in Korea. Foodborne Pathog. Dis. 2012, 9, 1057–1063. [Google Scholar] [CrossRef]
- Hang, B.P.T.; Wredle, E.; Börjesson, S.; Sjaunja, K.S.; Dicksved, J.; Duse, A. High level of multidrug-resistant Escherichia coli in young dairy calves in southern Vietnam. Trop. Anim. Health Prod. 2019, 51, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Mochimaru, T.; Morokuma, Y.; Kiyosuke, M.; Fujise, M.; Eto, F.; Eriguchi, Y.; Nagasaki, Y.; Shimono, N.; Kang, D. Clonal spread in Eastern Asia of ciprofloxacin-resistant Escherichia coli serogroup O25 strains, and associated virulence factors. Int. J. Antimicrob. Agents 2010, 35, 444–450. [Google Scholar] [CrossRef]
- Uchida, Y.; Mochimaru, T.; Morokuma, Y.; Kiyosuke, M.; Fujise, M.; Eto, F.; Harada, Y.; Kadowaki, M.; Shimono, N.; Kang, D. Geographic distribution of fluoroquinolone-resistant Escherichia coli strains in Asia. Int. J. Antimicrob. Agents 2010, 35, 387–391. [Google Scholar] [CrossRef]
- Takasu, H.; Suzuki, S.; Reungsang, A.; Viet, P.H. Fluoroquinolone (FQ) contamination does not correlate with occurrence of FQ-resistant bacteria in aquatic environments of vietnam and thailand. Microbes Environ. 2011, 26, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Tyson, G.H.; Li, C.; Hsu, C.H.; Bodeis-Jones, S.; McDermott, P.F. Diverse Fluoroquinolone Resistance Plasmids From Retail Meat E. coli in the United States. Front. Microbiol. 2019, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lascols, C.; Robert, J.; Cattoir, V.; Bébéar, C.; Cavallo, J.D.; Podglajen, I.; Ploy, M.C.; Bonnet, R.; Soussy, C.J.; Cambau, E. Type II topoisomerase mutations in clinical isolates of Enterobacter cloacae and other enterobacterial species harbouring the qnrA gene. Int. J. Antimicrob. Agents 2007, 29, 402–409. [Google Scholar] [CrossRef]
- Strahilevitz, J.; Jacoby, G.A.; Hooper, D.C.; Robicsek, A. Plasmid-mediated quinolone resistance: A multifaceted threat. Clin. Microbiol. Rev. 2009, 22, 664–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacoby, G.A.; Strahilevitz, J.; Hooper, D.C. Plasmid-mediated quinolone resistance—PMQR. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Tian, G.B.; Garcia, J.; Adams-Haduch, J.M.; Evangelista, J.P.; Destura, R.V.; Wang, H.N.; Doi, Y. CTX-M as the predominant extended-spectrum β-lactamases among Enterobacteriaceae in Manila, Philippines. J. Antimicrob. Chemother. 2010, 65, 584–586. [Google Scholar] [CrossRef]
- Kanamori, H.; Navarro, R.B.; Yano, H.; Sombrero, L.T.; Capeding, M.R.Z.; Lupisan, S.P.; Olveda, R.M.; Arai, K.; Kunishima, H.; Hirakata, Y.; et al. Molecular characteristics of extended-spectrum β-lactamases in clinical isolates of Enterobacteriaceae from the Philippines. Acta Trop. 2011, 120, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Umpiérrez, A.; Bado, I.; Oliver, M.; Acquistapace, S.; Etcheverría, A.; Padola, N.L.; Vignoli, R.; Zunino, P. Zoonotic potential and antibiotic resistance of Escherichia coli in neonatal calves in Uruguay. Microbes Environ. 2017, 32, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamani, R.; Flament-Simon, S.C.; García, V.; Mora, A.; Alonso, M.P.; López, C.; García-Meniño, I.; Díaz-Jiménez, D.; Blanco, J.E.; Blanco, M.; et al. Sequence types, clonotypes, serotypes, and virotypes of extended-spectrum β-lactamase-producing Escherichia coli causing bacteraemia in a Spanish Hospital over a 12-Year Period (2000 to 2011). Front. Microbiol. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salim, A.; Babu, P.; Mohan, K.; Moorthy, M.; Raj, M.; Thirumeni, S.K.; Suresh, S.; Madhavan, A.; Nair, B.G.; Chattopadhyay, S.; et al. Draft Genome Sequence of an Escherichia coli Sequence Type 155 Strain Isolated from Sewage in Kerala, India. Microbiol. Resour. Announc. 2019, 8, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Baquero, F.; Tedim, A.P.; Coque, T.M. Antibiotic resistance shaping multi-level population biology of bacteria. Front. Microbiol. 2013, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The role of epidemic resistance plasmids and international high- risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Nazareno, J.; Nakano, R.; Mondoy, M.; Nakano, A.; Bugayong, P. Environmental Presence and Genetic Characteristics of Carbapenemase-Producing Enterobacteriaceae from Hospital Sewage and River Water in the Philippines. Appl. Environ. Microbiol. 2020, 86, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R. Escherichia coli and urinary tract infections: The role of poultry-meat. Clin. Microbiol. Infect. 2016, 22, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindle, P.; Zurfluh, K.; Nüesch-Inderbinen, M.; Von Ah, S.; Sidler, X.; Stephan, R.; Kümmerlen, D. Phenotypic and genotypic characteristics of Escherichia coli with non-susceptibility to quinolones isolated from environmental samples on pig farms. Porc. Health Manag. 2019, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.Y.; Wakeham, D.; Brouwers, H.J.M.; Cobbold, R.N.; Abraham, S.; Mollinger, J.L.; Johnson, J.R.; Chapman, T.A.; Gordon, D.M.; Barrs, V.R.; et al. Human-associated fluoroquinolone-resistant Escherichia coli clonal lineages, including ST354, isolated from canine feces and extraintestinal infections in Australia. Microbes Infect. 2015, 17, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhong, Z.; Luo, Y.; Cox, E.; Devriendt, B. Heat-stable enterotoxins of enterotoxigenic escherichia coli and their impact on host immunity. Toxins 2019, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Dubreuil, J.D. EAST1 toxin: An enigmatic molecule associated with sporadic episodes of diarrhea in humans and animals. J. Microbiol. 2019, 57, 541–549. [Google Scholar] [CrossRef]
- Itoh, Y.; Nagano, I.; Kunishima, M.; Ezaki, T. Laboratory investigation of enteroaggregative Escherichia coli O untypeable:H10 associated with a massive outbreak of gastrointestinal illness. J. Clin. Microbiol. 1997, 35, 2546–2550. [Google Scholar] [CrossRef] [Green Version]
- Savarino, S.J.; Fasano, A.; Watson, J.; Martin, B.M.; Levine, M.M.; Guandalini, S.; Guerry, P. Enteroaggregative Escherichia coli heat-stable enterotoxin 1 represents another subfamily of E. coli heat-stable toxin. Proc. Natl. Acad. Sci. USA. 1993, 90, 3093–3097. [Google Scholar] [CrossRef] [Green Version]
- Sukkua, K.; Manothong, S.; Sukhumungoon, P. Seroprevalence and molecular epidemiology of EAST1 gene-carrying escherichia coli from diarrheal patients and raw meats. J. Infect. Dev. Ctries. 2017, 11, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Ochi, S.; Shah, M.; Odoyo, E.; Bundi, M.; Miringu, G.; Guyo, S.; Wandera, E.; Kathiiko, C.; Kariuki, S.; Karama, M.; et al. An outbreak of diarrhea in Mandera, Kenya, due to Escherichia coli serogroup o-nontypable strain that had a coding gene for enteroaggregative E. coli heat-stable enterotoxin 1. Am. J. Trop. Med. Hyg. 2017, 96, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). M100: Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). M07: Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). M100: Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Utrarachkij, F.; Nakajima, C.; Changkwanyeun, R.; Siripanichgon, K.; Kongsoi, S.; Pornruangwong, S.; Changkaew, K.; Tsunoda, R.; Tamura, Y.; Suthienkul, O.; et al. Quinolone Resistance Determinants of Clinical Salmonella Enteritidis in Thailand. Microb. Drug Resist. 2017, 23, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, J.M.; Velasco, C.; Pascual, A.; García, I.; Martínez-Martínez, L. Correlation of quinolone resistance levels and differences in basal and quinolone-induced expression from three qnrA-containing plasmids. Clin. Microbiol. Infect. 2006, 12, 440–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robicsek, A.; Strahilevitz, J.; Sahm, D.F.; Jacoby, G.A.; Hooper, D.C. qnr prevalence in ceftazidime-resistant Enterobacteriaceae isolates from the United States. Antimicrob. Agents Chemother. 2006, 50, 2872–2874. [Google Scholar] [CrossRef] [Green Version]
- Armas-Freire, P.I.; Trueba, G.; Proaño-Bolaños, C.; Levy, K.; Zhang, L.; Marrs, C.F.; Cevallos, W.; Eisenberg, J.N.S. Unexpected distribution of the fluoroquinolone-resistance gene qnrB in Escherichia coli isolates from different human and poultry origins in Ecuador. Int. Microbiol. 2015, 18, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Minarini, L.A.R.; Poirel, L.; Cattoir, V.; Darini, A.L.C.; Nordmann, P. Plasmid-mediated quinolone resistance determinants among enterobacterial isolates from outpatients in Brazil. J. Antimicrob. Chemother. 2008, 62, 474–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frasson, I.; Cavallaro, A.; Bergo, C.; Richter, S.N.; Palù, G. Prevalence of aac(6′)-Ib-cr plasmid-mediated and chromosome-encoded fluoroquinolone resistance in Enterobacteriaceae in Italy. Gut Pathog. 2011, 3, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, M.; Otomo, Y.; Ahsan, C.R. A novel single-step multiplex polymerase chain reaction assay for the detection of diarrheagenic Escherichia coli. J. Microbiol. Methods 2013, 92, 289–292. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample Source | Sample Type | Number of Samples | 95% Confidence Interval | |
|---|---|---|---|---|
| Examined | Positive (%) | |||
| Supermarket | Beef | 54 | 31 (57.4) | 0.4321–0.7077 |
| Chicken | 47 | 41 (87.2) | 0.7426–0.9517 | |
| Pork | 68 | 44 (64.7) | 0.5217–0.7592 | |
| Open market | Beef | 52 | 26 (50.0) | 0.3581–0.6419 |
| Chicken | 68 | 50 (73.5) | 0.6143–0.8350 | |
| Pork | 48 | 34 (70.8) | 0.5594–0.8305 | |
| Abattoir | Beef | 28 | 15 (53.6) | 0.3387–0.7249 |
| Pork | 23 | 11 (47.8) | 0.2682–0.6941 | |
| Environmental swab | 4 | 4 (100.0) | 0.3976–1.000 | |
| Swine farm | Rectal swab | 43 | 29 (67.4) | 0.5146–0.8092 |
| Environmental swab | 31 | 23 (74.2) | 0.5539–0.8814 | |
| Poultry farm | Cloacal swab | 45 | 24 (53.3) | 0.3787–0.6834 |
| Environmental swab | 10 | 6 (60.0) | 0.2624–0.8784 | |
| Egg | 50 | 1 (2.0) | 0.0005–0.1065 | |
| Buffalo farm | Milk | 30 | 0 (0.0) | 0.0000–0.1157 |
| Total tested | 601 | 339 (56.4) | 0.5234–0.6041 | |
| Antimicrobial Agents | Sample, n (%) | Total (n = 791) | ||||||
|---|---|---|---|---|---|---|---|---|
| Beef (n = 191) | Chicken (n = 224) | Pork (n = 188) | Egg (n = 8) | Cloacal Swabs (n = 52) | Rectal Swabs (n = 55) | Environmental Swabs (n = 73) | ||
| TET | 66 (34.6) | 148 (66.1) | 93 (49.5) | 3 (37.5) | 40 (76.9) | 53 (96.4) | 52 (71.2) | 455 (57.5) |
| AMP | 63 (33.0) | 143 (63.8) | 104 (55.3) | 7 (87.5) | 34 (65.4) | 46 (83.6) | 51 (69.9) | 448 (56.6) |
| SXT | 32 (16.8) | 104 (46.4) | 87 (46.3) | 2 (25.0) | 39 (75.0) | 37 (67.3) | 45 (61.6) | 346 (43.7) |
| CHL | 37 (19.4) | 86 (38.4) | 65 (34.6) | 1 (12.5) | 16 (30.8) | 39 (70.9) | 40 (54.8) | 284 (35.9) |
| STR | 32 (16.8) | 92 (41.1) | 49 (26.1) | 0 | 21 (40.4) | 19 (34.5) | 24 (32.9) | 237 (30.0) |
| NAL | 11 (5.8) | 101 (45.1) | 23 (12.2) | 0 | 43 (82.7) | 13 (23.6) | 9 (12.3) | 200 (25.3) |
| KAN | 9 (4.7) | 64 (28.6) | 7 (3.7) | 0 | 15 (28.8) | 15 (27.3) | 6 (8.2) | 116 (14.7) |
| CIP | 2 (1.0) | 57 (25.4) | 6 (3.2) | 0 | 21 (40.4) | 3 (5.5) | 3 (4.1) | 92 (11.6) |
| CST | 13 (6.8) | 38 (17.0) | 9 (4.8) | 2 (25.0) | 6 (11.5) | 5 (9.1) | 5 (6.8) | 78 (9.9) |
| AmC * | 6 (6.8) a | 12 (10.3) b | 10 (11.4) c | 0 d | 0 e | 4 (10.0) f | 0 g | 32 (8.9) |
| IPM | 3 (1.6) | 43 (19.2) | 5 (2.7) | 0 | 15 (28.8) | 2 (3.6) | 2 (2.7) | 70 (8.8) |
| GEN | 5 (2.6) | 42 (18.8) | 11 (5.9) | 0 | 2 (3.8) | 2 (3.6) | 5 (6.8) | 67 (8.5) |
| FOX | 10 (5.2) | 28 (12.5) | 13 (6.9) | 0 | 9 (17.3) | 3 (5.5) | 3 (4.1) | 66 (8.3) |
| CTX * | 4 (4.5) a | 6 (5.1) b | 2 (2.3) c | 0 d | 6 (60.0) e | 2 (20.0) f | 0 g | 20 (5.5) |
| CAZ | 5 (2.6) | 9 (4.0) | 3 (1.6) | 0 | 3 (5.8) | 1 (1.8) | 3 (4.1) | 24 (3.0) |
| FEP | 3 (1.6) | 6 (2.7) | 1 (0.5) | 0 | 4 (7.7) | 3 (5.5) | 1 (1.4) | 18 (2.3) |
| MEM * | 0 a | 2 (1.7) b | 1 (1.1) c | 0 d | 0 e | 0 f | 0 g | 3 (0.8) |
| No. of Antimicrobials Classes | Number (%) of Quinolone-Resistant Isolates | |||||||
|---|---|---|---|---|---|---|---|---|
| Beef (n = 191) | Chicken (n = 224) | Pork (n = 188) | Egg (n = 8) | Cloacal Swabs (n = 52) | Rectal Swabs (n = 55) | Environmental Swabs (n = 73) | Total (n = 791) | |
| 0 | 89 (46.6) | 20 (8.9) | 51 (27.1) | 0 (0.0) | 2 (3.8) | 1 (1.8) | 6 (8.2) | 169 (21.4) |
| 1–2 | 50 (26.2) | 52 (23.2) | 46 (24.5) | 6 (75.0) | 5 (9.6) | 6 (10.9) | 19 (26.0) | 184 (23.3) |
| 3–4 | 38 (19.9) | 67 (29.9) | 52 (27.7) | 2 (25.0) | 20 (38.5) | 19 (34.5) | 27 (37.0) | 225 (28.4) |
| 5–6 | 13 (6.8) | 60 (26.8) | 37 (19.7) | 0 | 16 (30.8) | 25 (45.5) | 18 (24.7) | 169 (21.4) |
| 7–8 | 1 (0.5) | 24 (10.7) | 2 (1.1) | 0 | 8 (15.4) | 4 (7.3) | 3 (4.1) | 42 (5.3) |
| >9 | 0 | 1 (0.4) | 0 | 0 | 1 (1.9) | 0 | 0 | 2 (0.3) |
| Resistance ≥ 1 | 102 (53.4) | 204 (91.1) | 137 (72.9) | 8 (100.0) | 50 (96.2) | 54 (98.2) | 48 (65.8) | 622 (78.6) |
| MDR ≥ 3 | 52 (27.2) | 152 (67.9) * | 91 (48.4) | 2 (25.0) | 45 (86.5) | 48 (87.3) | 48 (65.8) | 438 (55.4) |
| QRDR Amino-Acid Substitutions a | PMQR | No. of Isolates | MIC (μg/mL) d | ||||
|---|---|---|---|---|---|---|---|
| gyrA | gyrB | parC | parE | Nalidixic Acid | Ciprofloxacin | ||
| Ser83 Leu | -b | -b | -b | -c | 7 | 16 – >128 | 0.125–1 |
| Ser83 Leu | Ser492 Asn | -b | -b | -c | 1 | >128 | 0.5 |
| Ser83 Leu | -b | Ser80 Ile | -b | -c | 1 | >128 | 2 |
| Asp87 Tyr | -b | -b | -b | -c | 1 | >128 | 0.5 |
| Ser83 Leu; Asp87 Asn | - | -b | -b | -c | 1 | >128 | 4 |
| Ser83 Leu; Asp87 Asn | Ser492 Asn | Ser80 Ile; Glu84 Gly | -b | -c | 4 | >128 | >32 |
| Ser83 Leu; Asp87 Asn | -b | Ser80 Ile | -b | -c | 31 | >128 | 4–>32 |
| Ser83 Leu; Asp87 Asn | -b | Ser80 Ile | Ile464 Phe | -c | 2 | >128 | >32 |
| Ser83 Leu; Asp87 Asn | -b | Ser80 Ile | Ser458 Ala | -c | 16 | 128–>128 | 0.5–>32 |
| Ser83 Leu; Asp87 Asn | -b | Ser80 Ile; Glu84 Gly | -b | -c | 2 | >128 | >32 |
| Ser83 Leu | Ser492 Asn | Ser80 Arg | -b | qnrB4 | 1 | >128 | 8 |
| Ser83 Leu | -b | Ser80 Ile | -b | qnrS1 | 1 | >128 | 32 |
| Ser83 Leu | -b | -b | -b | qnrS1 | 1 | >128 | 4 |
| Ser83 Leu | -b | -b | -b | qnrB4 | 12 | >128 | 1–4 |
| Asp87 Try | -b | -b | -b | qnrS1 | 1 | >128 | 2 |
| Asp87 Try | -b | Ser80 Ile | -b | qnrB4 | 1 | 8 | <0.03 |
| Ser83 Leu; Asp87 Asn | S492 n | S80 I; E84 G | -b | qnrB4 | 3 | >128 | >32 |
| -b | S492 n | -b | -b | qnrB4 | 1 | 16 | 0.25 |
| -b | -b | -b | -b | qnrS1 | 27 | 8–>128 | 0.25–2 |
| -b | -b | -b | -b | qnrA1; qnrS1 | 1 | >128 | >32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belotindos, L.; Villanueva, M.; Miguel, J., Jr.; Bwalya, P.; Harada, T.; Kawahara, R.; Nakajima, C.; Mingala, C.; Suzuki, Y. Prevalence and Characterization of Quinolone-Resistance Determinants in Escherichia coli Isolated from Food-Producing Animals and Animal-Derived Food in the Philippines. Antibiotics 2021, 10, 413. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040413
Belotindos L, Villanueva M, Miguel J Jr., Bwalya P, Harada T, Kawahara R, Nakajima C, Mingala C, Suzuki Y. Prevalence and Characterization of Quinolone-Resistance Determinants in Escherichia coli Isolated from Food-Producing Animals and Animal-Derived Food in the Philippines. Antibiotics. 2021; 10(4):413. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040413
Chicago/Turabian StyleBelotindos, Lawrence, Marvin Villanueva, Joel Miguel, Jr., Precious Bwalya, Tetsuya Harada, Ryuji Kawahara, Chie Nakajima, Claro Mingala, and Yasuhiko Suzuki. 2021. "Prevalence and Characterization of Quinolone-Resistance Determinants in Escherichia coli Isolated from Food-Producing Animals and Animal-Derived Food in the Philippines" Antibiotics 10, no. 4: 413. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040413