
Antibiotic-Resistant Enterobacteriaceae in Wastewater of Abattoirs

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Isolates
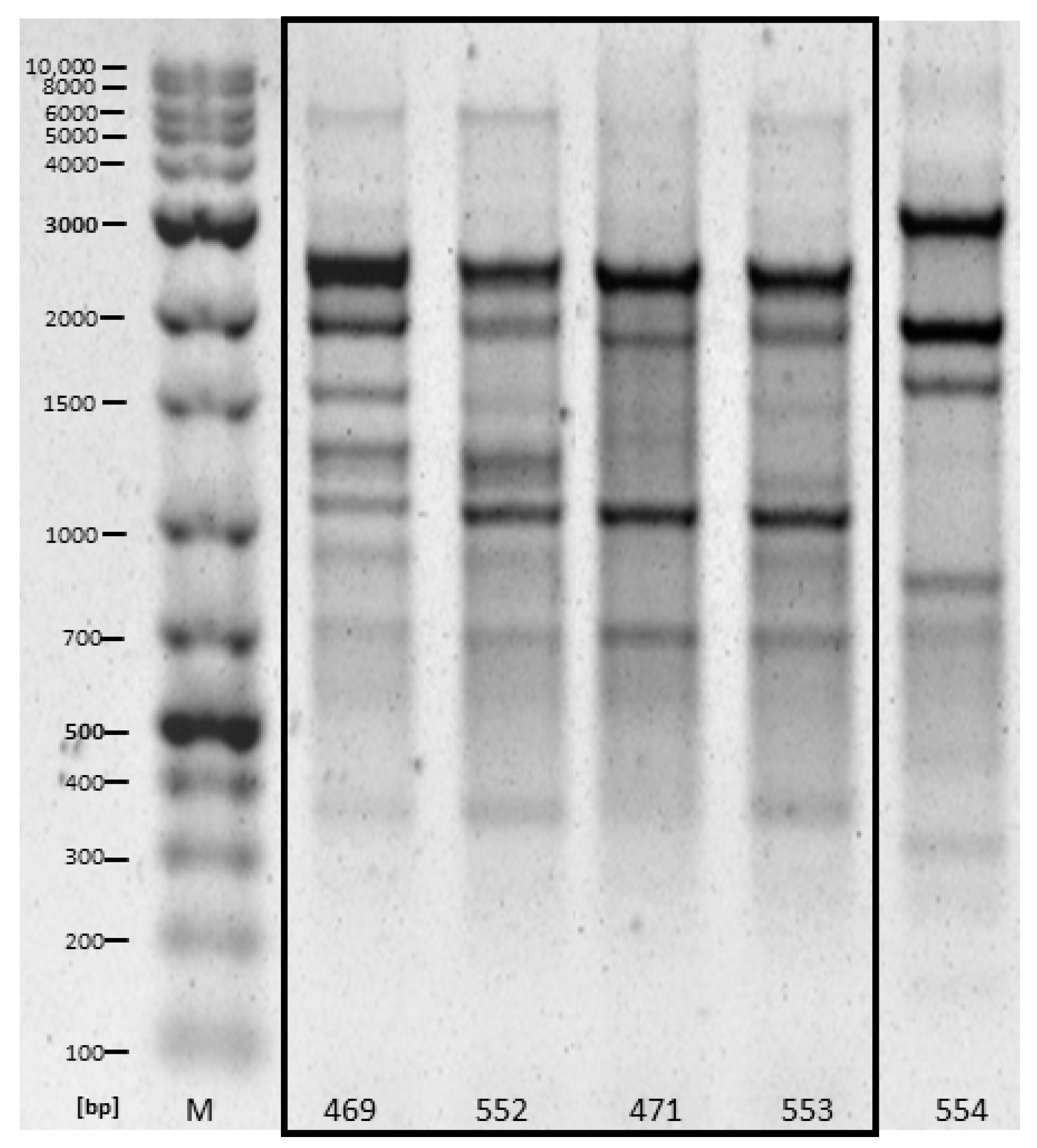
2.2. Similarity Analysis
2.3. Antimicrobial Susceptibility Testing (AST)
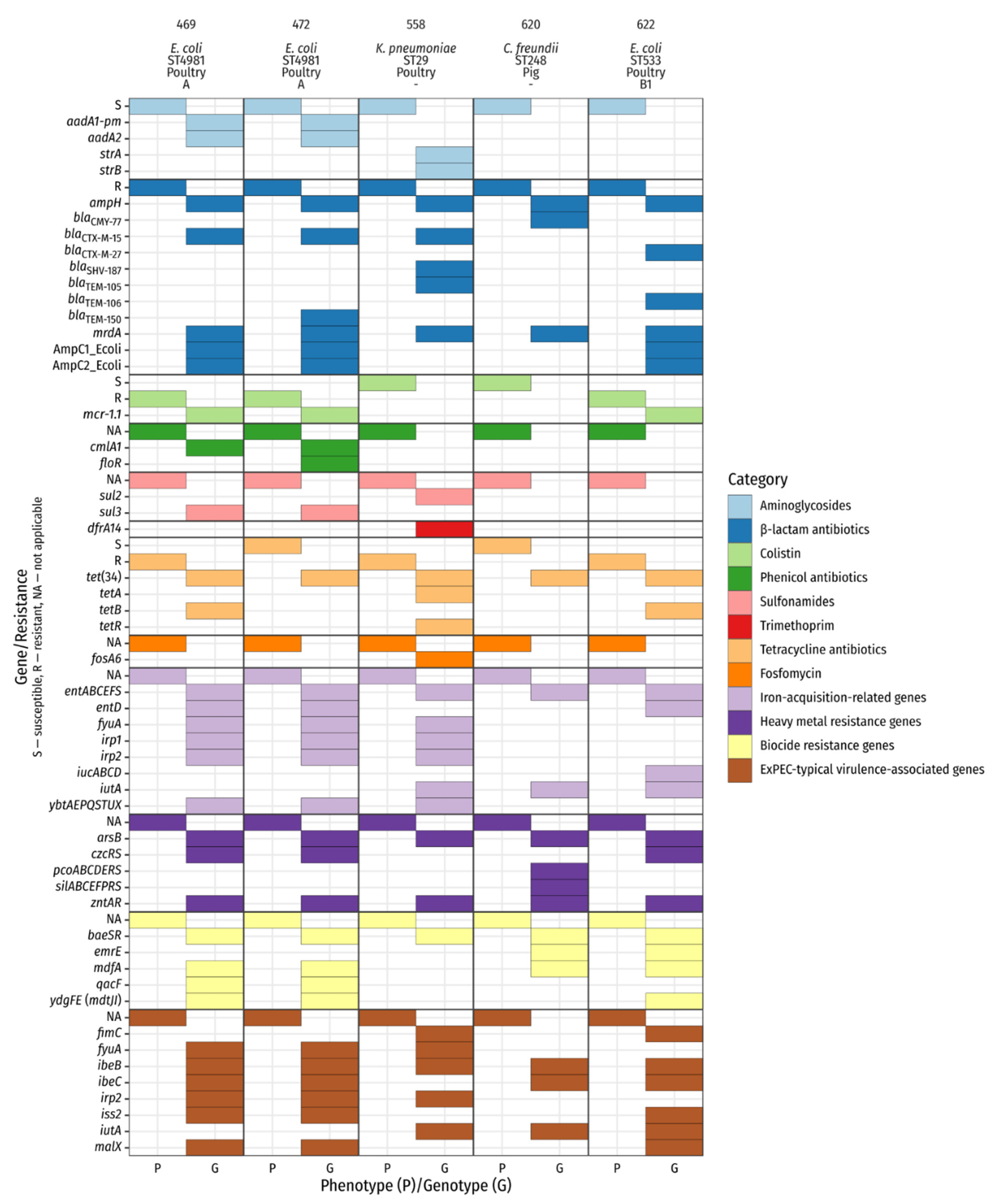
2.4. Whole-Genome Sequencing
3. Discussion
3.1. Resistance Data
3.2. Sequencing Data
3.3. Wastewater Treatment
4. Materials and Methods
4.1. Sampling Locations and Sample Collection
4.2. Isolation of Bacteria and Identification
4.3. Antimicrobial Susceptibility Testing (AST)
4.4. Similarity Analysis
4.5. Hypermucoviscosity
4.6. Sequence Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Antimicrobial Resistance Global Report on Surveillance; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Verburg, I.; Garcia-Cobos, S.; Leal, L.H.; Waar, K.; Friedrich, A.W.; Schmitt, H. Abundance and Antimicrobial Resistance of Three Bacterial Species Along a Complete Wastewater Pathway. Microorganisms 2019, 7, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The Who Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Knothe, H.; Shah, P.; Krcmery, V.; Antal, M.; Mitsuhashi, S. Transferable Resistance to Cefotaxime, Cefoxitin, Cefamandole and Cefuroxime in Clinical Isolates of Klebsiella pneumoniae and Serratia marcescens. Infection 1983, 11, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. Epidemiology of Β-Lactamase-Producing Pathogens. Clin. Microbiol. Rev. 2020, 33, e00047-19. [Google Scholar] [CrossRef]
- Woolhouse, M.; Ward, M.; van Bunnik, B.; Farrar, J. Antimicrobial Resistance in Humans, Livestock and the Wider Environment. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140083. [Google Scholar] [CrossRef]
- European Medicines Agency—European Surveillance of Veterinary Antimicrobial Consumption. “Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2018”. EMA-ESVAC, 2020. Available online: https://www.ema.europa.eu/en/documents/report/sales-veterinary-antimicrobial-agents-31-european-countries-2018-trends-2010-2018-tenth-esvac-report_en.pdf (accessed on 1 February 2021).
- van Rennings, L.; von Münchhausen, C.; Hartmann, M.; Ottilie, H.; Honscha, W.; Käsbohrer, A.; Kreienbrock, L. [Antibiotic Usage and Antibiotic Sales in Germany in 2011—The Situation of Drug Usage in Veterinary Medicine]. Berl. Munch. Tierarztl. Wochenschr. 2014, 127, 366–374. (In German) [Google Scholar]
- German Federal Ministry of Food and Agriculture. “Pressemitteilung Nr. 157/2019.” German Federal Ministry of Food and Agriculture. 2019. Available online: https://www.bmel.de/SharedDocs/Pressemitteilungen/DE/2019/157-reserve-antibiotika.html (accessed on 1 February 2021).
- Huebner, N.O.; Dittmann, K.; Henck, V.; Wegner, C.; Kramer, A. Epidemiology of Multidrug Resistant Bacterial Organisms and Clostridium Difficile in German Hospitals in 2014: Results from a Nationwide One-Day Point Prevalence of 329 German Hospitals. BMC Infect. Dis. 2016, 16, 467. [Google Scholar] [CrossRef] [Green Version]
- Blaak, H.; van Hoek, A.H.A.M.; Hamidjaja, R.A.; van der Plaats, R.Q.J.; Kerkhof-de Heer, L.; Husman, A.M.D.; Schets, F.M. Distribution, Numbers, and Diversity of Esbl-Producing E. coli in the Poultry Farm Environment. PLoS ONE 2015, 10, e0135402. [Google Scholar]
- Ghodousi, A.; Bonura, C.; Di Noto, A.M.; Mammina, C. Extended-Spectrum Beta-Lactamase, Ampc-Producing, and Fluoroquinolone-Resistant Escherichia coli in Retail Broiler Chicken Meat, Italy. Foodborne Pathog. Dis. 2015, 12, 619–625. [Google Scholar] [CrossRef] [Green Version]
- Savin, M.; Bierbaum, G.; Hammerl, J.A.; Heinemann, C.; Parcina, M.; Sib, E.; Voigt, A.; Kreyenschmidt, J. Antibiotic-Resistant Bacteria and Antimicrobial Residues in Wastewater and Process Water from German Pig Slaughterhouses and Their Receiving Municipal Wastewater Treatment Plants. Sci. Total Environ. 2020, 727, 138788. [Google Scholar] [CrossRef] [PubMed]
- Savin, M.; Bierbaum, G.; Blau, K.; Parcina, M.; Sib, E.; Smalla, K.; Schmithausen, R.; Heinemann, C.; Hammerl, J.A.; Kreyenschmidt, J. Colistin-Resistant Enterobacteriaceae Isolated from Process Waters and Wastewater from German Poultry and Pig Slaughterhouses. Front. Microbiol. 2020, 11, 575391. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wang, Y.; Shi, X.; Wang, S.; Ren, H.; Shen, Z.; Wang, Y.; Lin, J.; Wang, S. Rapid Rise of the Esbl and Mcr-1 Genes in Escherichia coli of Chicken Origin in China, 2008–2014. Emerg. Microbes Infect. 2018, 7, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzopardi, E.A.; Boyce, D.E.; Thomas, D.W.; Dickson, W.A. Colistin in Burn Intensive Care: Back to the Future? Burns 2013, 39, 7–15. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization—Advisory Group on Integrated Surveillance of Antimicrobial Resistance. Critically Important Antimicrobials for Human Medicine, 5th ed.; WHO—AGISAR: Geneva, Switzerland, 2017; Volume 5. [Google Scholar]
- Statistisches Bundesamt. “Viehbestand und Tierische Erzeugung 2017, Fachserie 3 Reihe 4.” Destatis. 2018. Available online: https://www.destatis.de/DE/Themen/Branchen-Unternehmen/Landwirtschaft-Forstwirtschaft-Fischerei/Tiere-Tierische-Erzeugung/Publikationen/Downloads-Tiere-und-tierische-Erzeugung/viehbestand-tierische-erzeugung-2030400177005.html (accessed on 1 February 2021).
- Mittal, G.S. Characterization of the Effluent Wastewater from Abattoirs for Land Application. Food Rev. Int. 2004, 20, 229–256. [Google Scholar] [CrossRef]
- Savin, M.; Bierbaum, G.; Hammerl, J.A.; Heinemann, C.; Parcina, M.; Sib, E.; Voigt, A.; Kreyenschmidt, J. Eskape Bacteria and Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Isolated from Wastewater and Process Water from German Poultry Slaughterhouses. Appl. Environ. Microb. 2020, 86, e02748-19. [Google Scholar] [CrossRef] [Green Version]
- Savin, M.; Bierbaum, G.; Kreyenschmidt, J.; Schmithausen, R.M.; Sib, E.; Schmoger, S.; Käsbohrer, A.; Hammerl, J.A. Clinically Relevant Escherichia coli Isolates from Process Waters and Wastewater of Poultry and Pig Slaughterhouses in Germany. Microorganisms 2021, 9, 698. [Google Scholar] [CrossRef]
- Schaekel, F.; May, T.; Seiler, J.; Hartmann, M.; Kreienbrock, L. Antibiotic Drug Usage in Pigs in Germany—Are the Class Profiles Changing? PLoS ONE 2017, 12, e0182661. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, T.H.; Aslam, B.; Arshad, M.I.; Alvi, R.F.; Muzammil, S.; Yasmeen, N.; Aslam, M.A.; Khurshid, M.; Rasool, M.H.; Baloch, Z. Emergence of Bla (Ndm-1) Harboring Klebsiella pneumoniae St29 and St11 in Veterinary Settings and Waste of Pakistan. Infect. Drug Resist. 2020, 13, 3033–3043. [Google Scholar] [CrossRef]
- Eibach, D.; Dekker, D.; Boahen, K.G.; Akenten, C.W.; Sarpong, N.; Campos, C.B.; Berneking, L.; Aepfelbacher, M.; Krumkamp, R.; Owusu-Dabo, E.; et al. Extended-Spectrum Beta-Lactamase-Producing Escherichia coli and Klebsiella pneumoniae in Local and Imported Poultry Meat in Ghana. Vet. Microbiol. 2018, 217, 7–12. [Google Scholar] [CrossRef]
- Kock, R.; Daniels-Haardt, I.; Becker, K.; Mellmann, A.; Friedrich, A.W.; Mevius, D.; Schwarz, S.; Jurke, A. Carbapenem-Resistant Enterobacteriaceae in Wildlife, Food-Producing, and Companion Animals: A Systematic Review. Clin. Microbiol. Infect. 2018, 24, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Anyanwu, M.U.; Jaja, I.F.; Nwobi, O.C. Occurrence and Characteristics of Mobile Colistin Resistance (Mcr) Gene-Containing Isolates from the Environment: A Review. Int. J. Environ. Res. Public Health 2020, 17, 1028. [Google Scholar] [CrossRef] [Green Version]
- Catry, B.; Cavaleri, M.; Baptiste, K.; Grave, K.; Grein, K.; Holm, A.; Jukes, H.; Liebana, E.; Lopez Navas, A.; Mackay, D.; et al. Use of Colistin-Containing Products within the European Union and European Economic Area (Eu/Eea): Development of Resistance in Animals and Possible Impact on Human and Animal Health. Int. J. Antimicrob. Agents 2015, 46, 297–306. [Google Scholar] [CrossRef]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Letellier, A. Colistin in Pig Production: Chemistry, Mechanism of Antibacterial Action, Microbial Resistance Emergence, and One Health Perspectives. Front. Microbiol. 2016, 7, 1789. [Google Scholar] [CrossRef] [PubMed]
- Apostolakos, I.; Piccirillo, A. A Review on the Current Situation and Challenges of Colistin Resistance in Poultry Production. Avian Pathol. 2018, 47, 546–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irrgang, A.; Roschanski, N.; Tenhagen, B.A.; Grobbel, M.; Skladnikiewicz-Ziemer, T.; Thomas, K.; Roesler, U.; Kasbohrer, A. Prevalence of Mcr-1 in E. coli from Livestock and Food in Germany, 2010–2015. PLoS ONE 2016, 11, e0159863. [Google Scholar] [CrossRef]
- Gekenidis, M.-T.; Qi, W.; Hummerjohann, J.; Zbinden, R.; Walsh, F.; Drissner, D. Antibiotic-Resistant Indicator Bacteria in Irrigation Water: High Prevalence of Extended-Spectrum Beta-Lactamase (Esbl)-Producing Escherichia coli. PLoS ONE 2018, 13, e0207857. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, J.D.; Haenni, M.; Metayer, V.; Madec, J.Y.; Ferreira, H.M.N. Epidemic Spread of Inci1/Pst113 Plasmid Carrying the Extended-Spectrum Beta-Lactamase (Esbl) Blactx-M-8 Gene in Escherichia coli of Brazilian Cattle. Vet. Microbiol. 2020, 243, 108629. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-C.; Yeh, K.-S. Characteristics of Extended-Spectrum Β-Lactamase–Producing Escherichia coli Isolated from Fecal Samples of Piglets with Diarrhea in Central and Southern Taiwan in 2015. BMC Vet. Res. 2017, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli Phylo-Typing Method Revisited: Improvement of Specificity and Detection of New Phylo-Groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Bonnedahl, J.; Drobni, M.; Gauthier-Clerc, M.; Hernandez, J.; Granholm, S.; Kayser, Y.; Melhus, Å.; Kahlmeter, G.; Waldenström, J.; Johansson, A.; et al. Dissemination of Escherichia coli with Ctx-M Type Esbl between Humans and Yellow-Legged Gulls in the South of France. PLoS ONE 2009, 4, e5958. [Google Scholar] [CrossRef]
- Huber, H.; Zweifel, C.; Wittenbrink, M.M.; Stephan, R. Esbl-Producing Uropathogenic Escherichia coli Isolated from Dogs and Cats in Switzerland. Vet. Microbiol. 2013, 162, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Cormier, A.; Zhang, P.L.C.; Chalmers, G.; Weese, J.S.; Deckert, A.; Mulvey, M.; McAllister, T.; Boerlin, P. Diversity of Ctx-M-Positive Escherichia coli Recovered from Animals in Canada. Vet. Microbiol. 2019, 231, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.; Sellera, F.P.; Cunha, M.P.V.; Lopes, R.; Cerdeira, L.; Lincopan, N. Emergence of Ctx-M-27-Producing Escherichia coli of St131 and Clade C1-M27 in an Impacted Ecosystem with International Maritime Traffic in South America. J. Antimicrob. Chemother. 2020, 75, 1647–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, H.; Doijad, S.; Falgenhauer, L.; Fritzenwanker, M.; Imirzalioglu, C.; Chakraborty, T. Bla(Ctx-M-27)-Encoding Escherichia coli Sequence Type 131 Lineage C1-M27 Clone in Clinical Isolates, Germany. Emerg. Infect. Dis. 2017, 23, 1754–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccirilli, A.; Pompilio, A.; Rossi, L.; Segatore, B.; Amicosante, G.; Rosatelli, G.; Perilli, M.; Di Bonaventura, G. Identification of Ctx-M-15 and Ctx-M-27 in Antibiotic-Resistant Gram-Negative Bacteria Isolated from Three Rivers Running in Central Italy. Microb. Drug Resist. 2019, 25, 1041–1049. [Google Scholar] [CrossRef]
- Dohmen, W.; Van Gompel, L.; Schmitt, H.; Liakopoulos, A.; Heres, L.; Urlings, B.A.; Mevius, D.; Bonten, M.J.M.; Heederik, D.J.J. Esbl Carriage in Pig Slaughterhouse Workers Is Associated with Occupational Exposure. Epidemiol. Infect. 2017, 145, 2003–2010. [Google Scholar] [CrossRef] [Green Version]
- Shon, A.S.; Bajwa, R.P.S.; Russo, T.A. Hypervirulent (Hypermucoviscous) Klebsiella pneumoniae: A New and Dangerous Breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; Marr, C.M. Hypervirulent Klebsiella pneumoniae. Clin. Microbiol. Rev. 2019, 32, e00001-19. [Google Scholar] [CrossRef] [Green Version]
- Moura, Q.; Esposito, F.; Fernandes, M.R.; Espinoza-Muñoz, M.; Souza, T.A.; Santos, S.R.; Cerdeira, L.; Cassettari, V.; Lincopan, N. Genome Sequence Analysis of a Hypermucoviscous/Hypervirulent and Mdr Ctx-M-15/K19/St29 Klebsiella pneumoniae Isolated from Human Infection. Pathog. Dis. 2017, 75, ftx121. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Zhang, K.; Chen, W.; Chen, J.; Zheng, J.; Liu, C.; Cheng, L.; Zhou, W.; Shen, H.; Cao, X. Epidemiological Characteristics of Carbapenem-Resistant Enterobacteriaceae Collected from 17 Hospitals in Nanjing District of China. Antimicrob. Resist. Infect. Control 2020, 9, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. Commission Implementing Decision (Eu) 2016/902 of 30 May 2016 Establishing Best Available Techniques (Bat) Conclusions, under Directive 2010/75/Eu of the European Parliament and of the Council, for Common Waste Water and Waste Gas Treatment/Management Systems in the Chemical Sector. In C(2016)3127; European Commission: Brussels, Belgium, 2016; Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32016D0902 (accessed on 1 February 2021).
- Parnanen, K.M.M.; Narciso-da-Rocha, C.; Kneis, D.; Berendonk, T.U.; Cacace, D.; Do, T.T.; Elpers, C.; Fatta-Kassinos, D.; Henriques, I.; Jaeger, T.; et al. Antibiotic Resistance in European Wastewater Treatment Plants Mirrors the Pattern of Clinical Antibiotic Resistance Prevalence. Sci. Adv. 2019, 5, eaau9124. [Google Scholar] [CrossRef] [Green Version]
- Flach, C.F.; Genheden, M.; Fick, J.; Larsson, D.G.J. A Comprehensive Screening of Escherichia coli Isolates from Scandinavia’s Largest Sewage Treatment Plant Indicates No Selection for Antibiotic Resistance. Environ. Sci. Technol. 2018, 52, 11419–11428. [Google Scholar] [CrossRef] [PubMed]
- Karkman, A.; Do, T.T.; Walsh, F.; Virta, M.P.J. Antibiotic-Resistance Genes in Waste Water. Trends Microbiol. 2018, 26, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manaia, C.M.; Rocha, J.; Scaccia, N.; Marano, R.; Radu, E.; Biancullo, F.; Cerqueira, F.; Fortunato, G.; Iakovides, I.C.; Zammit, I.; et al. Antibiotic Resistance in Wastewater Treatment Plants: Tackling the Black Box. Environ. Int. 2018, 115, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Ravelo, C.; Magariños, B.; López-Romalde, S.; Toranzo, A.E.; Romalde, J.L. Molecular Fingerprinting of Fish-Pathogenic Lactococcus garvieae Strains by Random Amplified Polymorphic DNA Analysis. J. Clin. Microbiol. 2003, 41, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Rocco, L.; Valentino, I.V.; Scapigliati, G.; Stingo, V. Rapd-Pcr Analysis for Molecular Characterization and Genotoxic Studies of a New Marine Fish Cell Line Derived from Dicentrarchus Labrax. Cytotechnology 2014, 66, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Baym, M.; Kryazhimskiy, S.; Lieberman, T.D.; Chung, H.; Desai, M.M.; Kishony, R. Inexpensive Multiplexed Library Preparation for Megabase-Sized Genomes. PLoS ONE 2015, 10, e0128036. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. Spades: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Proc, G.P.D. The Sequence Alignment/Map Format and Samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.D.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. Checkm: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: Bigsdb Software, the Pubmlst.Org Website and Their Applications [Version 1; Peer Review: 2 Approved]. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.D.; Jin, Q.; Chen, L.H.; Yang, J. Vfdb 2019: A Comparative Pathogenomic Platform with an Interactive Web Interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids Using Plasmidfinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G.J. Bacmet: Antibacterial Biocide and Metal Resistance Genes Database. Nucleic Acids Res. 2014, 42, D737–D743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. Arg-Annot, a New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Slaughterhouse | Sampling Date | Selective Media | ||||
|---|---|---|---|---|---|---|
| ESBL | COL | MRSA | Carb | OXA-48 | ||
| A (swine) | November 2020 | + | + | - | - | - |
| December 2020 | + | - | - | - | - | |
| B (swine) | November 2020 | + | + | - | - | - |
| December 2020 | + | - | - | - | - | |
| C (swine) | November 2020 | + | + | - | - | - |
| December 2020 | + | - | - | - | - | |
| D (poultry) | November 2020 | + | + | - | - | - |
| December 2020 | + | + | - | - | - | |
| E (swine) | November 2020 | + | + | - | - | - |
| December 2020 | + | + | - | - | - | |
| F (poultry) | November 2020 | + | + | - | - | - |
| December 2020 | + | - | - | - | - | |
| G (poultry) | November 2020 | + | + | - | - | - |
| December 2020 | N/A | N/A | N/A | N/A | N/A | |
| Slaughterhouse | Sampling Date | Selective Media | |
|---|---|---|---|
| ESBL | COL | ||
| A (swine) | November 2020 | E. coli (2), K. pneumoniae (2) | R. ornithinolytica (2) |
| December 2020 | E. coli (2) | ||
| B (swine) | November 2020 | E. coli (5) | |
| December 2020 | E. coli (1) | ||
| C (swine) | November 2020 | E. coli (2) | |
| December 2020 | E. coli (2) | ||
| D (poultry) | November 2020 | E. coli (4), K. pneumoniae (1) | E. coli (2), K. pneumoniae (1) |
| December 2020 | E. coli (1), K. pneumoniae (1) | ||
| E (swine) | November 2020 | E. coli (3), E. cloacae (1) | |
| December 2020 * | E. coli (4), K. pneumoniae (2) | ||
| F (poultry) | November 2020 | E. coli (2), K. pneumoniae (2) | E. coli (2) |
| December 2020 | E. coli (2) | E. coli (2) | |
| G (poultry) | November 2020 | E. coli (2), K. pneumoniae (1), E. cloacae (1) | E. coli (2) |
| December 2020 | N/A | N/A | |
| Designation | Slaughterhouse | Sampling Month | Medium | Duplicate | Species | ESBL Production | MDR | Piperacillin | Cefuroxime | Cefotaxime | Ceftazidime | Meropenem | Imipenem | Gentamicin | Ciprofloxacin | Colistin | Tetracycline |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 467 | A | N | C | 1 | R. ornithinolytica | - | - | S | S | S | S | S | S | S | S | R | S |
| 468 | A | N | C | 2 | R. ornithinolytica | - | - | S | S | S | S | S | S | S | S | R | S |
| 469 * | D | N | C | 1 | E. coli | + | + | R | R | R | R | S | S | S | R | R | R |
| 470 | D | N | C | 2 | K. pneumoniae | + | + | R | R | R | R | S | S | S | R | R | S |
| 471 | D | N | C | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | R | R |
| 472 * | F | N | C | 1 | E. coli | + | + | R | R | R | R | S | S | S | R | R | S |
| 473 | F | N | C | 2 | E. coli | - | + | R | S | S | S | S | S | S | R | R | S |
| 474 | G | N | C | 1 | E. coli | + | + | R | R | S | R | S | S | S | R | R | S |
| 475 | G | N | C | 2 | E. coli | + | + | R | R | S | R | S | S | S | S | R | S |
| 545 | A | N | E | 1 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 546 | A | N | E | 2 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 547 | B | N | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | S | S | R |
| 548 | B | N | E | 2 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 549 | B | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | S | R |
| 550 | C | N | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | R | R | R |
| 551 | C | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | R | S | S | S |
| 552 | D | N | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | R | R | R |
| 553 | D | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | R | R |
| 554 | D | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | R | S |
| 555 | E | N | E | 1 | E. cloacae complex | + | + | R | R | R | R | S | S | R | R | S | S |
| 556 | E | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | S | R |
| 557 | F | N | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | R | S | S |
| 558 * | F | N | E | 1 | K. pneumoniae | + | + | R | R | R | R | S | S | S | R | S | R |
| 559 | F | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | S | R |
| 560 | F | N | E | 2 | K. pneumoniae | + | + | R | R | R | R | S | S | S | R | S | R |
| 561 | G | N | E | 1 | E. coli | + | + | R | R | S | R | S | S | S | S | R | S |
| 562 | G | N | E | 1 | K. pneumoniae | + | + | R | R | R | R | S | S | S | R | S | S |
| 563 | G | N | E | 2 | E. cloacae complex | + | + | R | R | R | R | S | S | S | R | S | R |
| 564 | G | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | S | S |
| 595 | E | D | E | 2 | E. coli | + | + | R | R | R | R | S | S | R | S | S | S |
| 618 | F | D | C | 1 | E. coli | + | + | R | R | R | R | S | S | S | S | R | R |
| 620 * | E | D | C | 1 | C. freundii | + | - | R | R | R | R | S | S | S | S | S | S |
| 622 * | F | D | C | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | R | R |
| 625 | A | D | E | 1 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 627 | F | D | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | S | S | R |
| 628 | F | D | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | S | R |
| 629 | E | D | E | 1 | K. pneumoniae | + | + | R | R | R | R | S | S | R | S | R | R |
| 630 | E | D | E | 1 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 631 | E | D | E | 2 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 632 | E | D | E | 2 | K. pneumoniae | + | + | R | R | R | R | S | S | S | S | S | R |
| 633 | A | D | E | 2 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 634 | A | N | E | 2 | K. pneumoniae | + | + | R | R | R | R | S | S | S | S | R | S |
| 635 | B | D | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | R | S | S |
| 636 | B | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | S | S | R |
| 637 | B | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | S | R |
| 638 | C | D | E | 1 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 639 | C | D | E | 2 | E. coli | + | - | R | R | R | R | S | S | S | S | S | S |
| 640 | D | D | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | S | S | R |
| 641 | D | D | E | 1 | K. pneumoniae | + | + | R | R | R | R | S | S | S | R | R | S |
| 642 | D | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | S | S | R |
| 643 | E | D | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | R | S | R |
| 648 | D | N | E | 1 | K. pneumoniae | + | + | R | R | R | R | S | S | R | R | S | S |
| 649 | E | N | E | 1 | E. coli | + | + | R | R | R | R | S | S | S | S | S | R |
| 650 | E | N | E | 2 | E. coli | + | + | R | R | R | R | S | S | S | R | S | R |
| 651 | A | N | E | 2 | K. pneumoniae | + | + | R | R | R | R | S | S | S | R | R | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Homeier-Bachmann, T.; Heiden, S.E.; Lübcke, P.K.; Bachmann, L.; Bohnert, J.A.; Zimmermann, D.; Schaufler, K. Antibiotic-Resistant Enterobacteriaceae in Wastewater of Abattoirs. Antibiotics 2021, 10, 568. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050568
Homeier-Bachmann T, Heiden SE, Lübcke PK, Bachmann L, Bohnert JA, Zimmermann D, Schaufler K. Antibiotic-Resistant Enterobacteriaceae in Wastewater of Abattoirs. Antibiotics. 2021; 10(5):568. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050568
Chicago/Turabian StyleHomeier-Bachmann, Timo, Stefan E. Heiden, Phillip K. Lübcke, Lisa Bachmann, Jürgen A. Bohnert, Dirk Zimmermann, and Katharina Schaufler. 2021. "Antibiotic-Resistant Enterobacteriaceae in Wastewater of Abattoirs" Antibiotics 10, no. 5: 568. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050568