
Antifungal and Anti-Inflammatory Potential of Bupleurum rigidum subsp. paniculatum (Brot.) H.Wolff Essential Oil

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Essential Oil Composition
2.2. Antifungal Activity
2.2.1. The Essential Oil Showed a Broad-Spectrum Fungicidal Effect
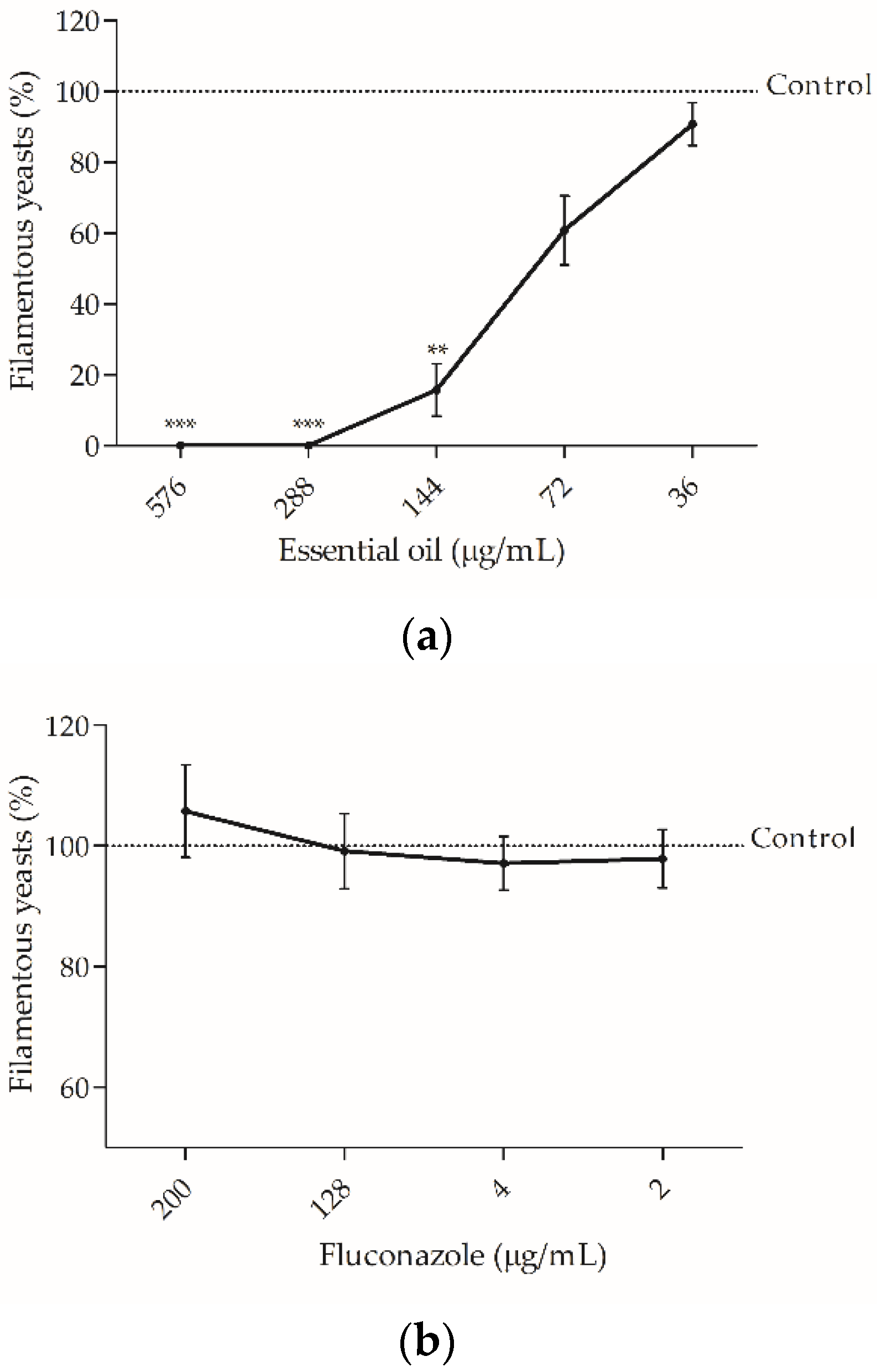
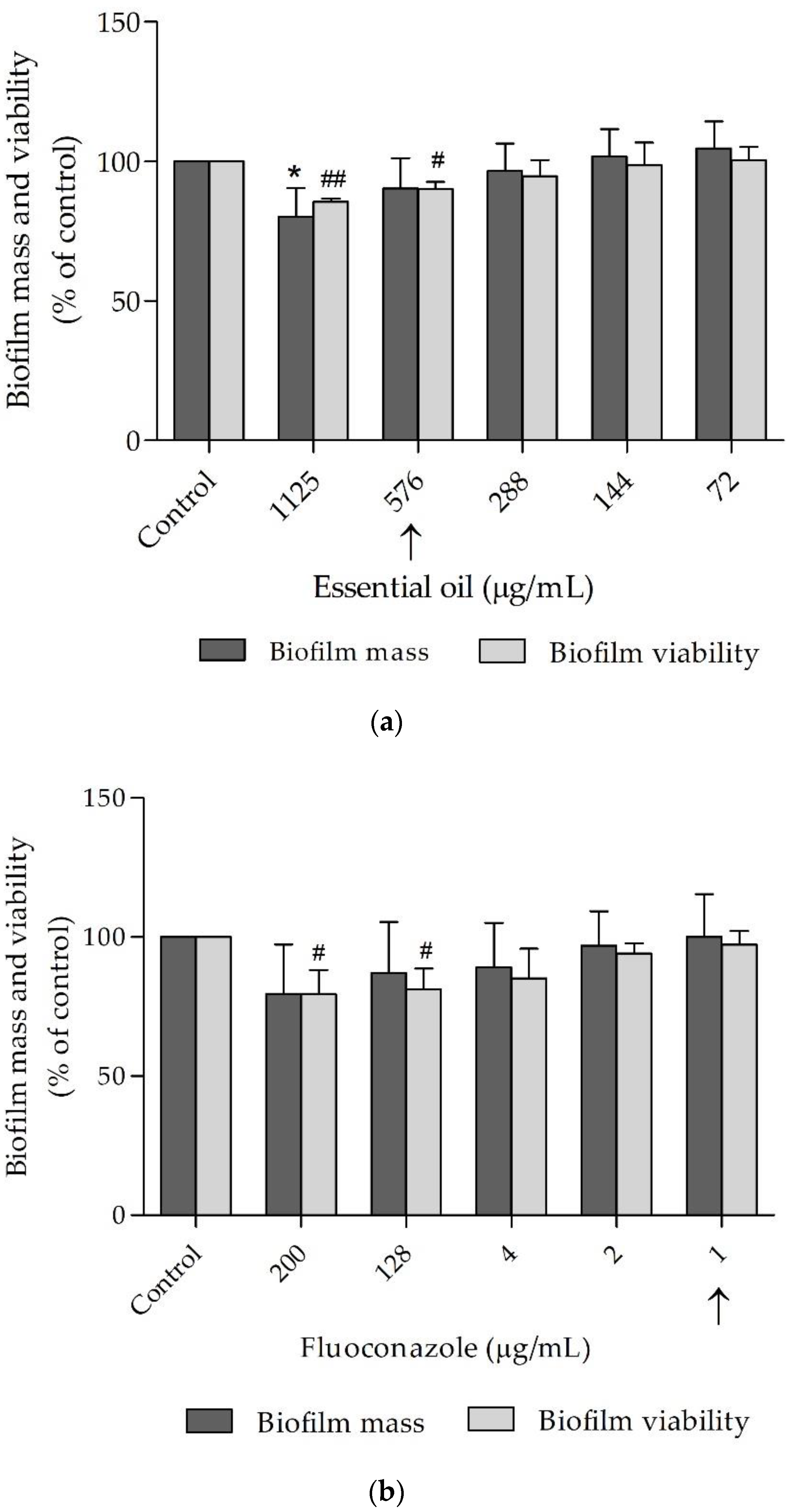
2.2.2. The Essential Oil was Effective against Candida albicans Virulent Factors
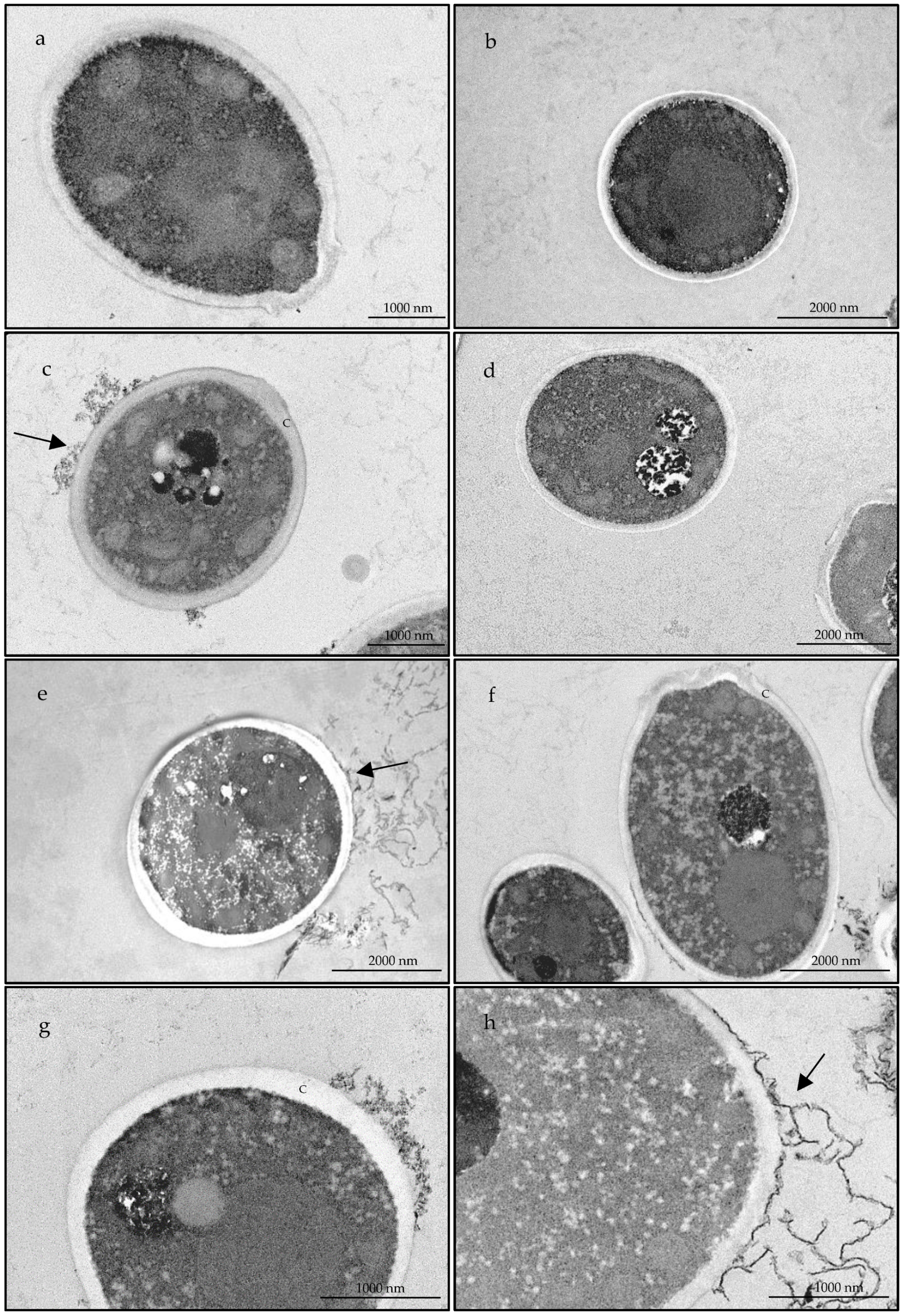
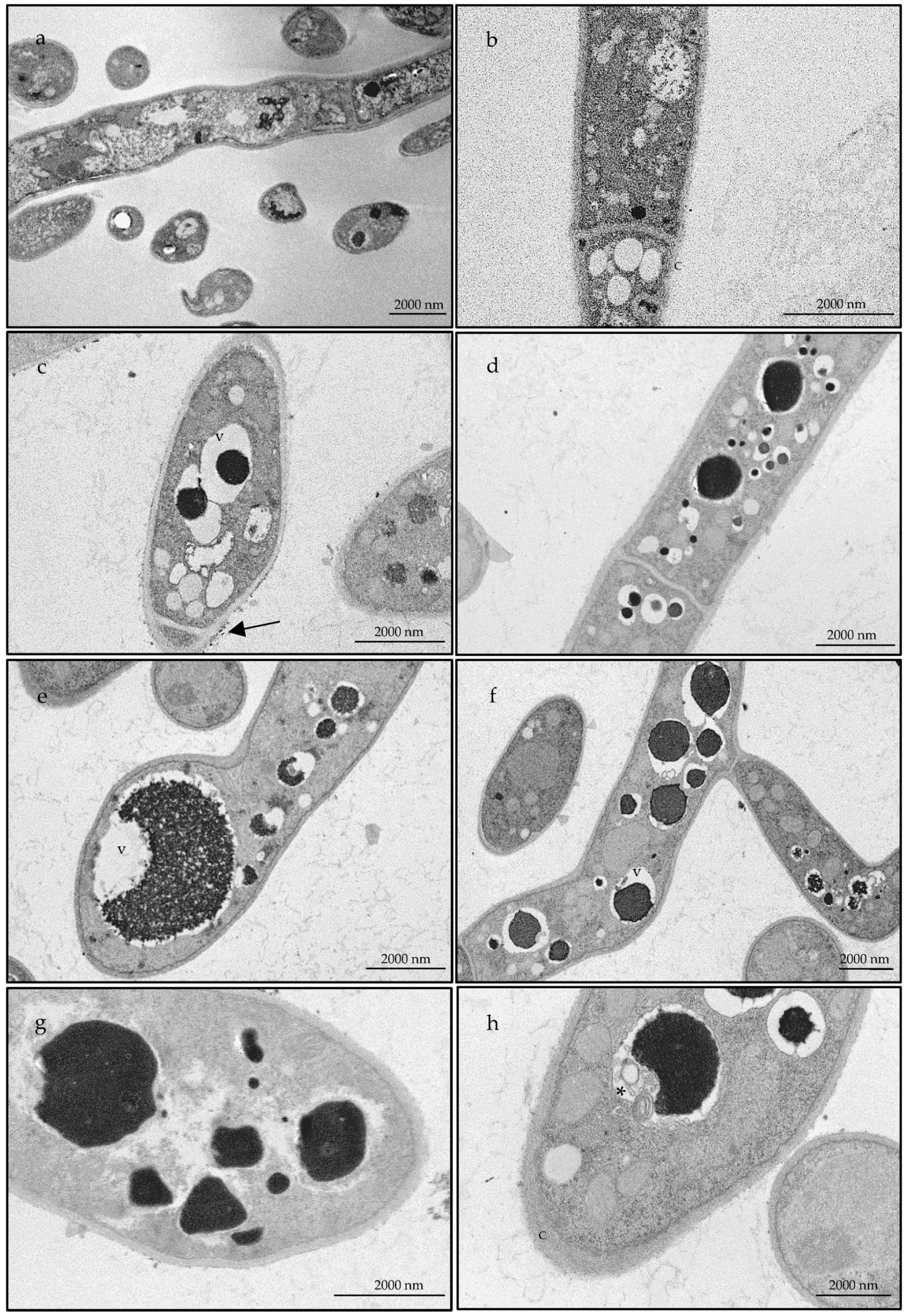
2.2.3. The Essential Oil Modified the Ultrastructure of Candida albicans and Trichophyton rubrum
2.3. Anti-Inflammatory Activity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Essential Oil Isolation and Chemical Analysis
5.3. Antifungal Activity
5.3.1. Fungal Strains
5.3.2. Fungal Growth
5.3.3. Fungal Ultrastructure
5.3.4. Candida albicans Germ Tube Formation
5.3.5. Candida albicans Biofilm Disruption
5.4. Anti-Inflammatory Activity
5.4.1. Cell Culture
5.4.2. Assessment of Nitric Oxide (NO) Production
5.4.3. Assessment of Cell Viability
5.5. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.; Denning, D. Global and multi-national prevalence of fungal diseases—Estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Pierce, C.; Lopez-Ribot, J.L. Candidiasis drug discovery and development: New approaches targeting virulence for discovering and identifying new drugs. Expert Opin. Drug Discov. 2013, 8, 1117–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makanjuola, O.; Bongomin, F.; Fayemiwo, S. An Update on the roles of non-albicans Candida species in vulvovaginitis. J. Fungi 2018, 4, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadar, M.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Shahali, Y.; Dhama, K. Candida albicans—Biology, molecular characterization, pathogenicity, and advances in diagnosis and control—An update. Microb. Pathog. 2018, 117, 128–138. [Google Scholar] [CrossRef]
- Romo, J.A.; Pierce, C.G.; Chaturvedi, A.K.; Lazzell, A.L.; McHardy, S.F.; Saville, S.P.; Lopez-Ribot, J.L. Development of anti-virulence approaches for candidiasis via a novel series of small-molecule inhibitors of Candida albicans filamentation. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Rodrigues, C.; Araújo, D.; Rodrigues, M.; Henriques, M. Candida species biofilms’ antifungal resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Kim, Y.-G.; Gupta, V.K.; Manoharan, R.K.; Lee, J. Suppression of fluconazole resistant Candida albicans biofilm formation and filamentation by methylindole derivatives. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Sugui, J.A.; Kwon-Chung, K.J.; Juvvadi, P.R.; Latge, J.-P.; Steinbach, W.J. Aspergillus fumigatus and related species. Cold Spring Harb. Perspect. Med. 2015, 5, a019786. [Google Scholar] [CrossRef] [Green Version]
- Fuentefria, A.M.; Pippi, B.; Dalla Lana, D.F.; Donato, K.K.; de Andrade, S.F. Antifungals discovery: An insight into new strategies to combat antifungal resistance. Lett. Appl. Microbiol. 2018, 66, 2–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Rossi, N.M.; Bitencourt, T.A.; Peres, N.T.A.; Lang, E.A.S.; Gomes, E.V.; Quaresemin, N.R.; Martins, M.P.; Lopes, L.; Rossi, A. Dermatophyte resistance to antifungal drugs: Mechanisms and prospectus. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Colombo, A.L.; de Almeida Júnior, J.N.; Guinea, J. Emerging multidrug-resistant Candida species. Curr. Opin. Infect. Dis. 2017, 30, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, M.; Khosravi, A.; Mortaz, E.; Nikaein, D.; Athari, S.S. Role of pathogen-associated molecular patterns (PAMPS) in immune responses to fungal infections. Eur. J. Pharm. 2017, 808, 8–13. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, R.J.S.; Xolalpa, N.C.; Castro, A.J.A.; González, C.P.; Ramos, J.P.; Gutiérrez, S.P. Terpenes from natural products with potential anti-inflammatory activity. In Terpenes and Terpenoids; IntechOpen: London, UK, 2018; pp. 59–85. [Google Scholar]
- Sun, S.-C. The non-canonical NF-κB pathway in immunity and inflammation. Nat. Rev. Immunol. 2017, 17, 545–558. [Google Scholar] [CrossRef]
- Kumamoto, C.A. Inflammation and gastrointestinal Candida colonization. Curr. Opin. Microbiol. 2011, 14, 386–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharm. 2020, 180, 114147. [Google Scholar] [CrossRef] [PubMed]
- Alves-Silva, J.M.; Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Cardoso, S.M.; Salgueiro, L. New claims for wild carrot (Daucus carota subsp. carota) essential oil. Evid. Based Complement. Altern. Med. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Valente, J.; Zuzarte, M.; Resende, R.; Gonçalves, M.J.; Cavaleiro, C.; Pereira, C.F.; Cruz, M.T.; Salgueiro, L. Daucus carota subsp. gummifer essential oil as a natural source of antifungal and anti-inflammatory drugs. Ind. Crop. Prod. 2015, 65, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Zou, Y.; Chen, X. Traditional use, pharmacology, toxicology, and quality control of species in genus Bupleurum L. Chin. Herb. Med. 2013, 5, 245–255. [Google Scholar] [CrossRef]
- Prieto, J.M.; Ogunsina, M.O.; Novak, A.; Joshi, A.; Kokai, J.; Rocha, I.D.C.; de Santayana, M.P. Comparative study of the in vitro bioactivities of Bupleurum rigidum and B. fruticescens. Nat. Prod. Commun. 2012, 7. [Google Scholar] [CrossRef]
- Ashour, M.L.; El-Readi, M.; Youns, M.; Mulyaningsih, S.; Sporer, F.; Efferth, T.; Wink, M. Chemical composition and biological activity of the essential oil obtained from Bupleurum marginatum (Apiaceae). J. Pharm. Pharm. 2009, 61, 1079–1087. [Google Scholar] [CrossRef]
- Lorente, I.; Ocete, M.A.; Zarzuelo, A.; Cabo, M.M.; Jimenez, J. Bioactivity of the essential oil of Bupleurum fruticosum. J. Nat. Prod. 1989, 52, 267–272. [Google Scholar] [CrossRef]
- Martin, S.; Padilla, E.; Ocete, M.; Galvez, J.; Jiménez, J.; Zarzuelo, A. Anti-inflammatory activity of the essential oil of Bupleurum Fruticescens. Planta Med. 1993, 59, 533–536. [Google Scholar] [CrossRef]
- Guinea, M.; Parellada, J.; Lacaille-Dubois, M.; Wagner, H. Biologically active triterpene saponins from Bupleurum fruticosum. Planta Med. 1994, 60, 163–167. [Google Scholar] [CrossRef]
- Yen, M.-H.; Weng, T.-C.; Liu, S.-Y.; Chai, C.-Y.; Lin, C.-C. The hepatoprotective effect of Bupleurum kaoi, an endemic plant to Taiwan, against dimethylnitrosamine-induced hepatic fibrosis in rats. Biol. Pharm. Bull. 2005, 28, 442–448. [Google Scholar] [CrossRef] [Green Version]
- Shi, B.; Liu, W.; Wei, S.-P.; Wu, W.-J. Chemical composition, antibacterial and antioxidant activity of the essential oil of Bupleurum longiradiatum. Nat. Prod. Commun. 2010, 5, 1139–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafaghat, A. Antioxidant, antimicrobial activities and fatty acid components of leaf and seed of Bupleurum lancifolium hornem. J. Med. Plants Res. 2011, 5, 3758–3762. [Google Scholar]
- Maxia, A.; Frau, M.A.; Marongiu, B.; Piras, A.; Porcedda, S.; Falconieri, D.; Gonçalves, M.J.; Cavaleiro, C.; Salgueiro, L. Biological assays and chemical composition of volatile oils of Bupleurum fruticosum L. (Apiaceae). Rec. Nat. Prod. 2011, 5, 92–99. [Google Scholar]
- Fernández-Ocaña, A.M.; Gómez-Rodríguez, M.V.; Velasco-Negueruela, A.; Camacho-Simarro, A.M.; Fernández-López, C.; Altarejos, J. In vivo antifungal activity of the essential oil of Bupleurum gibraltarium against Plasmopara halstedii in sunflower. J. Agric. Food Chem. 2004, 52, 6414–6417. [Google Scholar] [CrossRef]
- Lartaud, P.; Pegeon, A.; Pelletier, P. Cosmetic Use of Essential Oil of Bupleurum fruticosum 2011; WO/2012/080992; L’OREAL: Clichy, France, 2012. [Google Scholar]
- Contreras, S.S.; Lanza, A.M.D.; Pajares, M.B.; Esteban, C.B.; Castillo, L.V.; Martínez, M.J.A.; Benito, P.B.; Matellano, L.F. Bioactive components of Bupleurum rigidum L. subsp. rigidum. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2002; pp. 659–696. [Google Scholar]
- Mukaremera, L.; Lee, K.K.; Mora-Montes, H.M.; Gow, N.A.R. Candida albicans yeast, pseudohyphal, and hyphal morphogenesis differentially affects immune recognition. Front. Immunol. 2017, 8, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, N.N.R.; Alviano, C.S.; Blank, A.F.; Romanos, M.T.V.; Fonseca, B.B.; Rozental, S.; Rodrigues, I.A.; Alviano, D.S. Synergism effect of the essential oil from Ocimum basilicum var. Maria Bonita and its major components with fluconazole and its influence on ergosterol biosynthesis. Evid. Based Complement. Altern. Med. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romagnoli, C.; Baldisserotto, A.; Vicentini, C.; Mares, D.; Andreotti, E.; Vertuani, S.; Manfredini, S. Antidermatophytic action of resorcinol derivatives: Ultrastructural evidence of the activity of phenylethyl resorcinol against Microsporum Gypseum. Molecule 2016, 21, 1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palá-Paúl, J.; Pérez-Alonso, M.J.; Velasco-Negueruela, A. Volatile constituents isolated from the essential oil of Bupleurum rigidum ssp. paniculatum (Brot.) H. Wolff. J. Essent. Oil Res. 1999, 11, 456–458. [Google Scholar] [CrossRef]
- Correia, P.; Canhoto, J.M.; Zuzarte, M.; Salgueiro, L. Bupleurum spp. in central Portugal: In vitro propagation and secretory ducts. Acta Hortic. 2015, 1083, 527–534. [Google Scholar] [CrossRef]
- Yousefzadi, M.; Sonboli, A.; Ebrahimi, S.N.; Hashemi, S.H. Antimicrobial activity of essential oil and major constituents of Salvia chloroleuca. Z. Nat. C 2008, 63, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Tampieri, M.P.; Galuppi, R.; Macchioni, F.; Carelle, M.S.; Falcioni, L.; Cioni, P.L.; Morelli, I. The inhibition of Candida albicans by selected essential oils and their major components. Mycopathologia 2005, 159, 339–345. [Google Scholar] [CrossRef]
- Sonboli, A.; Babakhani, B.; Mehrabian, A.R. Antimicrobial activity of six constituents of essential oil from Salvia. Z. Nat. C. 2006, 61, 160–164. [Google Scholar]
- Rajput, S.B.; Karuppayil, S.M. Small molecules inhibit growth, viability and ergosterol biosynthesis in Candida albicans. Springerplus 2013, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Pinto, E.; Hrimpeng, K.; Lopes, G.; Vaz, S.; Gonçalves, M.J.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of Ferulago capillaris essential oil against Candida, Cryptococcus, Aspergillus and dermatophyte species. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Sugano, M.; Majid, A.; Fujii, Y. Antifungal effects of volatile compounds from black zira (Bunium persicum) and other spices and herbs. J. Chem. Ecol. 2007, 33, 2123–2132. [Google Scholar] [CrossRef]
- Leite, A.M.; de Oliveira Lima, E.; de Souza, E.L.; de Fátima Formiga Melo Diniz, M.; Trajano, V.N.; de Medeiros, I.A. Inhibitory effect of beta-pinene, alpha-pinene and eugenol on the growth of potential infectious endocarditis causing Gram-positive bacteria. Rev. Bras. Ciências Farm. 2007, 43, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antifungal activity of the components of Melaleuca alternifolia (tea tree) oil. J. Appl. Microbiol. 2003, 95, 853–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipowicz, N.; Kamiński, M.; Kurlenda, J.; Asztemborska, M.; Ochocka, J.R. Antibacterial and antifungal activity of juniper berry oil and its selected components. Phyther. Res. 2003, 17, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Halbandge, S.D.; Mortale, S.P.; Jadhav, A.K. Differential Sensitivities of various growth modes of Candida albicans to sixteen molecules of plant origin. J. Pharm. Phytochem. 2017, 6, 306–318. [Google Scholar]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Mohan Karuppayil, S. Terpenoids of plant origin inhibit morphogenesis, adhesion, and biofilm formation by Candida albicans. Biofouling 2013, 29, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Serra, E.; Hidalgo-Bastida, L.; Verran, J.; Williams, D.; Malic, S. Antifungal activity of commercial essential oils and biocides against Candida albicans. Pathogens 2018, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, C.; Prados-Rosales, R.; Silva, B.M.A.; Nakouzi-Naranjo, A.; Zuzarte, M.; Chatterjee, S.; Stark, R.E.; Casadevall, A.; Gonçalves, T. Activation of melanin synthesis in Alternaria infectoria by antifungal drugs. Antimicrob. Agents Chemother. 2016, 60, 1646–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, M.L.; Jimenez, J.; Ocete, M.A.; Zarzuelo, A.; Cabo, M.M. Comparative study of different essential oils of Bupleurum gibraltaricum Lamarck. Pharmazie 1989, 44, 284–287. [Google Scholar]
- Ocete, M.A.; Risco, S.; Zarzuelo, A.; Jimenez, J. Pharmacological activity of the essential oil of Bupleurum gibraltaricum: Anti-inflammatory activity and effects on isolated rat uteri. J. Ethnopharmacol. 1989, 25, 305–313. [Google Scholar] [CrossRef]
- Coté, H.; Boucher, M.-A.; Pichette, A.; Legault, J. Anti-inflammatory, antioxidant, antibiotic, and cytotoxic activities of Tanacetum vulgare L. essential oil and its constituents. Medicines 2017, 4, 34. [Google Scholar] [CrossRef]
- Rufino, A.T.; Ribeiro, M.; Sousa, C.; Judas, F.; Salgueiro, L.; Cavaleiro, C.; Mendes, A.F. Evaluation of the anti-inflammatory, anti-catabolic and pro-anabolic effects of E-caryophyllene, myrcene and limonene in a cell model of osteoarthritis. Eur. J. Pharm. 2015, 750, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Rufino, A.T.; Ribeiro, M.; Judas, F.; Salgueiro, L.; Lopes, M.C.; Cavaleiro, C.; Mendes, A.F. Anti-inflammatory and chondroprotective activity of (+)-α-pinene: Structural and enantiomeric selectivity. J. Nat. Prod. 2014, 77, 264–269. [Google Scholar] [CrossRef]
- Ehrnhöfer-Ressler, M.M.; Fricke, K.; Pignitter, M.; Walker, J.M.; Walker, J.; Rychlik, M.; Somoza, V. Identification of 1,8-cineole, borneol, camphor, and thujone as anti-inflammatory compounds in a Salvia officinalis L. infusion using human gingival fibroblasts. J. Agric. Food Chem. 2013, 61, 3451–3459. [Google Scholar] [CrossRef] [PubMed]
- de Lima, V.T.; Vieira, M.C.; Kassuya, C.A.L.; Cardoso, C.A.L.; Alves, J.M.; Foglio, M.A.; de Carvalho, J.E.; Formagio, A.S.N. Chemical composition and free radical-scavenging, anticancer and anti-inflammatory activities of the essential oil from Ocimum kilimandscharicum. Phytomedicine 2014, 21, 1298–1302. [Google Scholar] [CrossRef]
- Ramalho, T.; Pacheco de Oliveira, M.; Lima, A.; Bezerra-Santos, C.; Piuvezam, M. Gamma-terpinene modulates acute inflammatory response in mice. Planta Med. 2015, 81, 1248–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council of Europe. European Pharmacopoeia, 9th ed.; Council of Europe: Strasbourg, France, 2016. [Google Scholar]
- Marichal, P.; Gorrens, J.; Cutsem, J.; Bossche, H. Vanden Culture media for the study of the effects of azole derivatives on germ tube formation and hyphal growth of C. albicans/Nährböden zur Untersuchung der Wirkungen von Azolderivaten auf die Keimschlauchbildung und das Hyphenwachstum bei C. albicans. Mycoses 2009, 29, 76–81. [Google Scholar] [CrossRef]
- Alves, M.; Gonçalves, M.J.; Zuzarte, M.; Alves-Silva, J.M.; Cavaleiro, C.; Cruz, M.T.; Salgueiro, L. Unveiling the antifungal potential of two Iberian thyme essential oils: Effect on C. albicans germ tube and preformed biofilms. Front. Pharm. 2019, 10. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound * | RI SPB-1 a | RI SW 10 b | Coimbra (%) | Fátima (%) |
|---|---|---|---|---|
| α-Thujene | 922 | 1029 | 0.3 | 0.1 |
| α-Pinene | 930 | 1030 | 36.0 | 29.0 |
| Camphene | 943 | 1077 | 0.2 | 0.4 |
| Fenchene | 943 | 1067 | 0.1 | 0.1 |
| Sabinene | 964 | 1128 | 2.0 | 2.6 |
| β-Pinene | 970 | 1118 | 26.1 | 30.7 |
| Myrcene | 981 | 1160 | 1.8 | 10.0 |
| α-Phellandrene | 997 | 1171 | 0.3 | 0.1 |
| α-Terpinene | 1006 | 1183 | t | t |
| p-Cymene | 1009 | 1271 | 0.2 | t |
| Limonene | 1021 | 1204 | 10.5 | 13.5 |
| β-Phellandrene | 1021 | 1214 | 9.4 | 4.5 |
| Z-β-Ocimene | 1025 | 1235 | 4.1 | 1.0 |
| E-β-Ocimene | 1035 | 1253 | 5.1 | 2.0 |
| γ-Terpinene | 1046 | 1249 | 0.3 | 0.2 |
| Terpinolene | 1076 | 1288 | 0.1 | 0.1 |
| Monoterpene hydrocarbons | 96.6 | 94.4 | ||
| Linalool | 1082 | 1543 | t | t |
| α-Pinene-xide | 1077 | 1370 | t | t |
| allo-Ocimene | 1118 | 1370 | 0.1 | 0.1 |
| Terpinene-4-ol | 1159 | 1595 | 0.1 | t |
| Myrtenal | 1176 | 1786 | 0.1 | t |
| α-Terpineol | 1168 | 1692 | t | t |
| Bornyl acetate | 1266 | 1578 | 0.1 | 0.1 |
| Oxygen containing monoterpenes | 0.5 | 0.3 | ||
| α-Copaene | 1364 | 1487 | 0.1 | t |
| β-Cubebene | 1380 | 1538 | t | t |
| E-Caryophyllene | 1408 | 1590 | 0.2 | 0.1 |
| γ-Elemene | 1417 | 2136 | t | t |
| α-Humulene | 1442 | 1665 | t | t |
| Germacrene-D | 1466 | 1699 | 1.7 | 0.5 |
| Bicyclogermacrene | 1478 | 1727 | 0.2 | 0.1 |
| δ-Cadinene | 1508 | 1751 | 0,1 | t |
| Sesquiterpene hydrocarbons | 2.4 | 0.8 | ||
| Spathulenol | 1551 | 2112 | t | t |
| Caryophyllene oxide | 1557 | 1968 | t | t |
| Oxygen containing sesquiterpenes | 0.1 | 0.1 | ||
| Total identified | 99.6 | 95.6 |
| Strains | Essential Oil | Fluconazole | ||
|---|---|---|---|---|
| MIC | MLC | MIC | MLC | |
| Candida albicans ATCC 10231 | 576 | 576 | 1 | >128 |
| Candida tropicalis ATCC 13803 | 1125 | 1125 | 4 | >128 |
| Candida krusei H9 | 288 | 288 | 64 | 64–128 |
| Candida guillermondii MAT23 | 288 | 288 | 8 | 8 |
| Candida parapsilosis ATCC 90018 | 576 | 576 | 1 | 1–2 |
| Cryptococcus neoformans CECT 1078 | 72 | 144 | 16 | 128 |
| Trichophytum mentagrophytes FF7 | 288 | 288 | 16–32 | 32–64 |
| Trichophytum mentagrophytes var. interdigitale CECT 2958 | 288 | 288 | 128 | ≥128 |
| Trichophyton rubrum CECT 2794 | 72 | 72 | 16 | 64 |
| Trichophytum verrucosum CECT 2992 | 288 | 288 | 128 | >128 |
| Microsporum canis FF1 | 144 | 144 | 128 | 128 |
| Microsporum gypseum CECT 2905 | 288 | 288 | 128 | >128 |
| Epidermophyton floccosum FF9 | 144 | 144 | 16 | 16 |
| Aspergillus niger ATCC 16404 | 288 | 1125 | n.t | n.t |
| Aspergillus fumigatus ATCC 46645 | 576 | 576 | n.t | n.t |
| Aspergillus flavus F44 | 576 | 576 | n.t | n.t |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuzarte, M.; Correia, P.M.P.; Alves-Silva, J.M.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, T.; Salgueiro, L. Antifungal and Anti-Inflammatory Potential of Bupleurum rigidum subsp. paniculatum (Brot.) H.Wolff Essential Oil. Antibiotics 2021, 10, 592. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050592
Zuzarte M, Correia PMP, Alves-Silva JM, Gonçalves MJ, Cavaleiro C, Cruz T, Salgueiro L. Antifungal and Anti-Inflammatory Potential of Bupleurum rigidum subsp. paniculatum (Brot.) H.Wolff Essential Oil. Antibiotics. 2021; 10(5):592. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050592
Chicago/Turabian StyleZuzarte, Mónica, Pedro M. P. Correia, Jorge M. Alves-Silva, Maria J. Gonçalves, Carlos Cavaleiro, Teresa Cruz, and Lígia Salgueiro. 2021. "Antifungal and Anti-Inflammatory Potential of Bupleurum rigidum subsp. paniculatum (Brot.) H.Wolff Essential Oil" Antibiotics 10, no. 5: 592. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050592