
Evaluation of the Photodynamic Therapy with Curcumin on L. braziliensis and L. major Amastigotes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
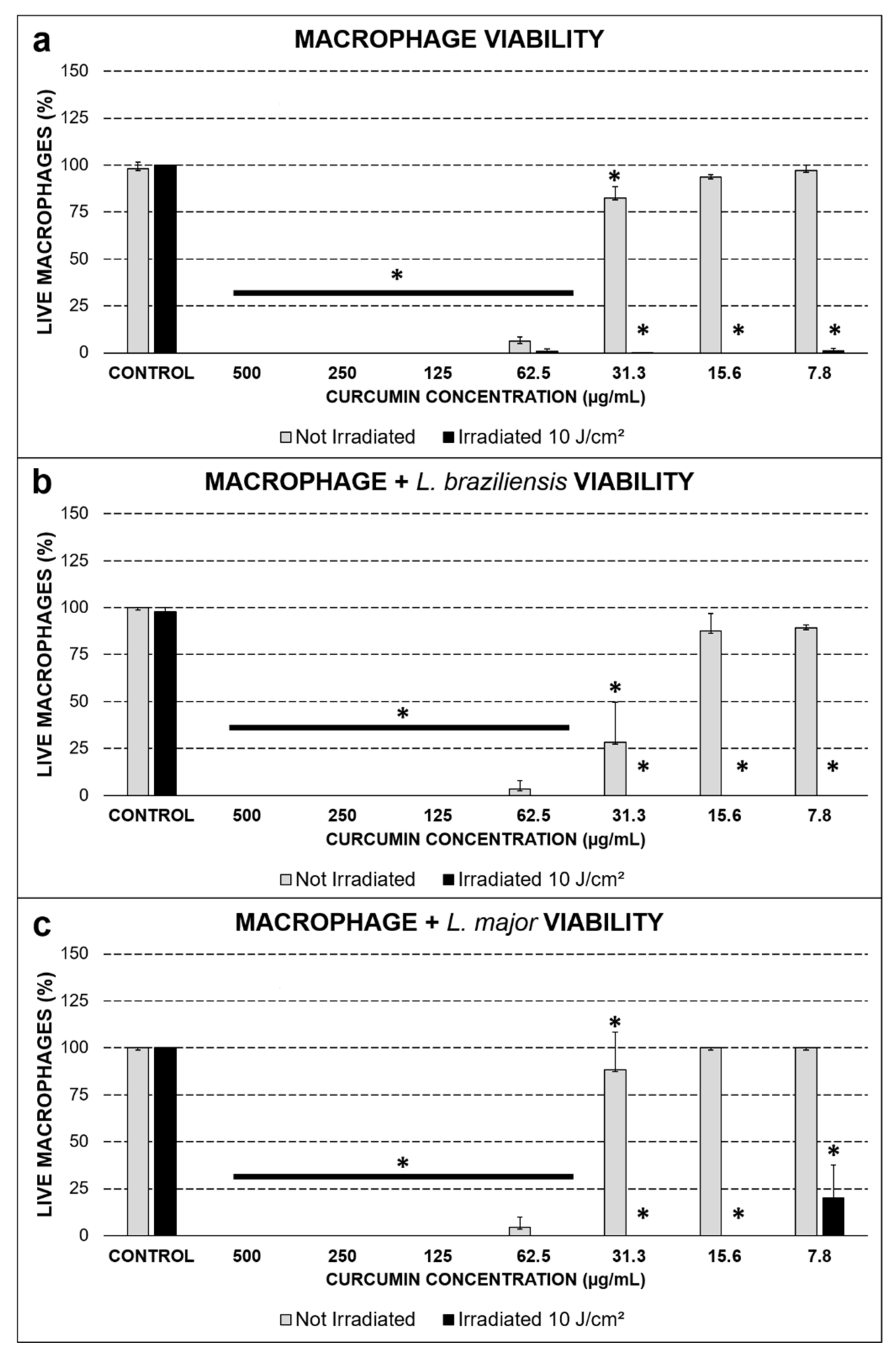
2.1. Viability Assessed by the Trypan Blue Exclusion Test
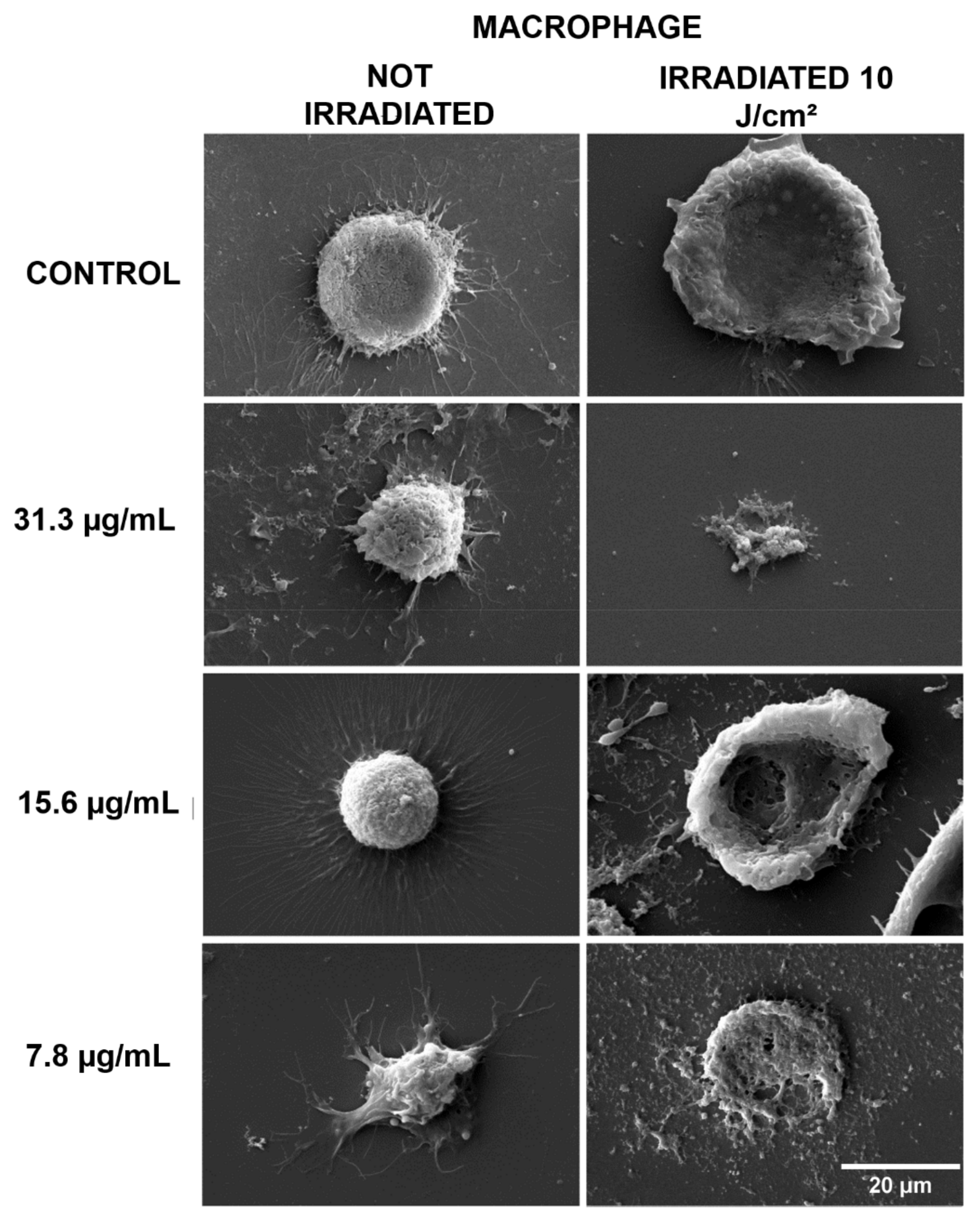
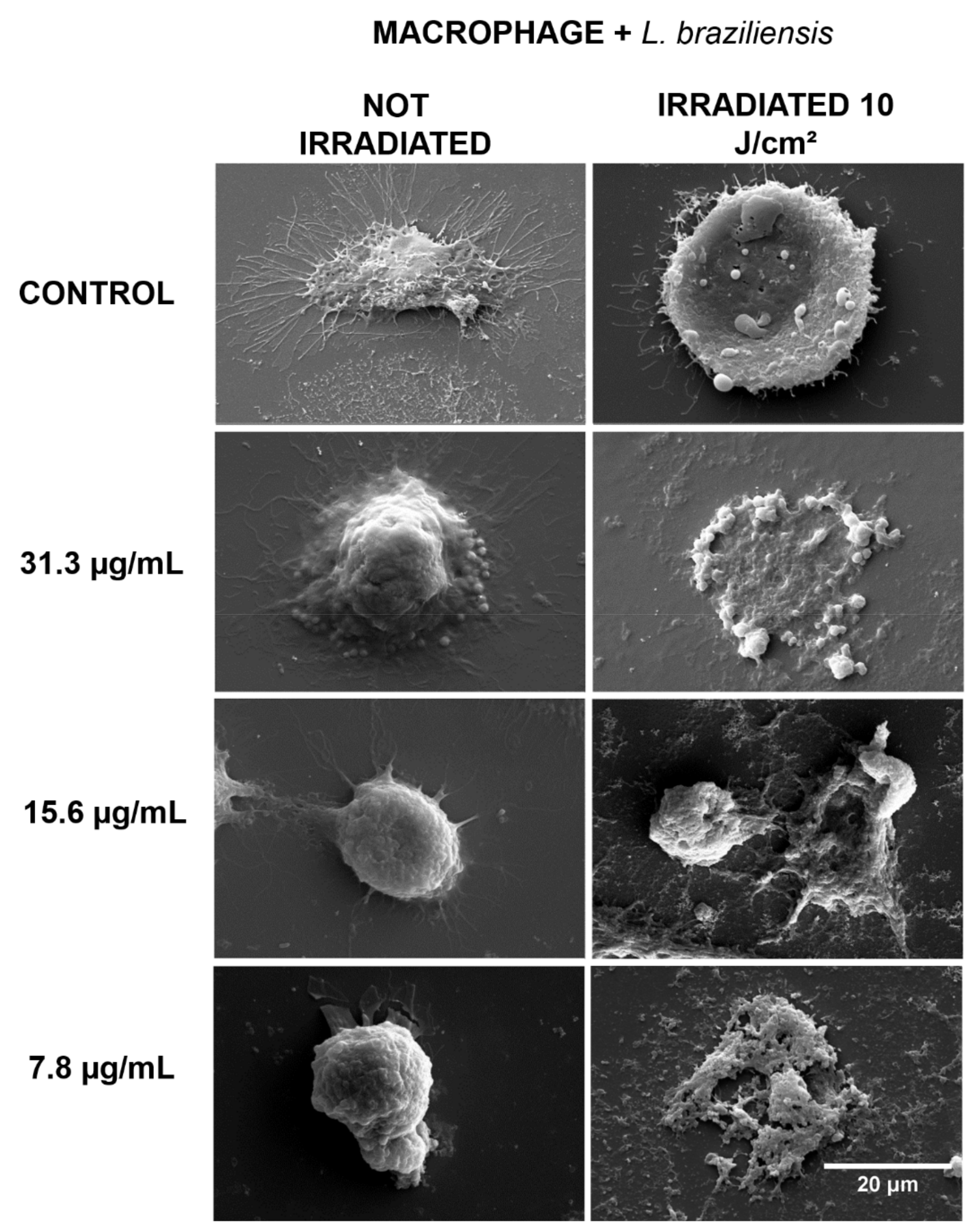
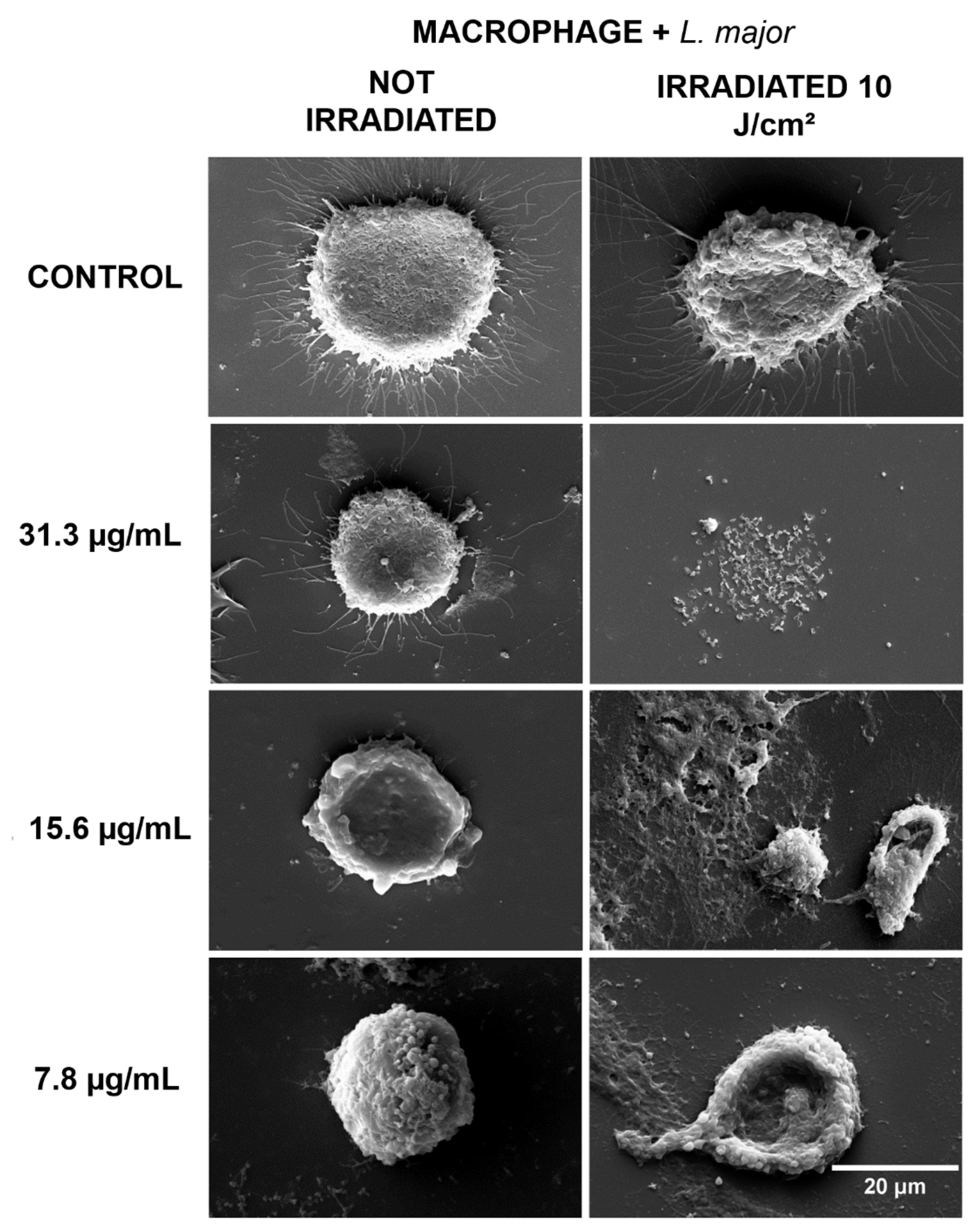
2.2. Morphological Evaluation by Scanning Electron Microscopy
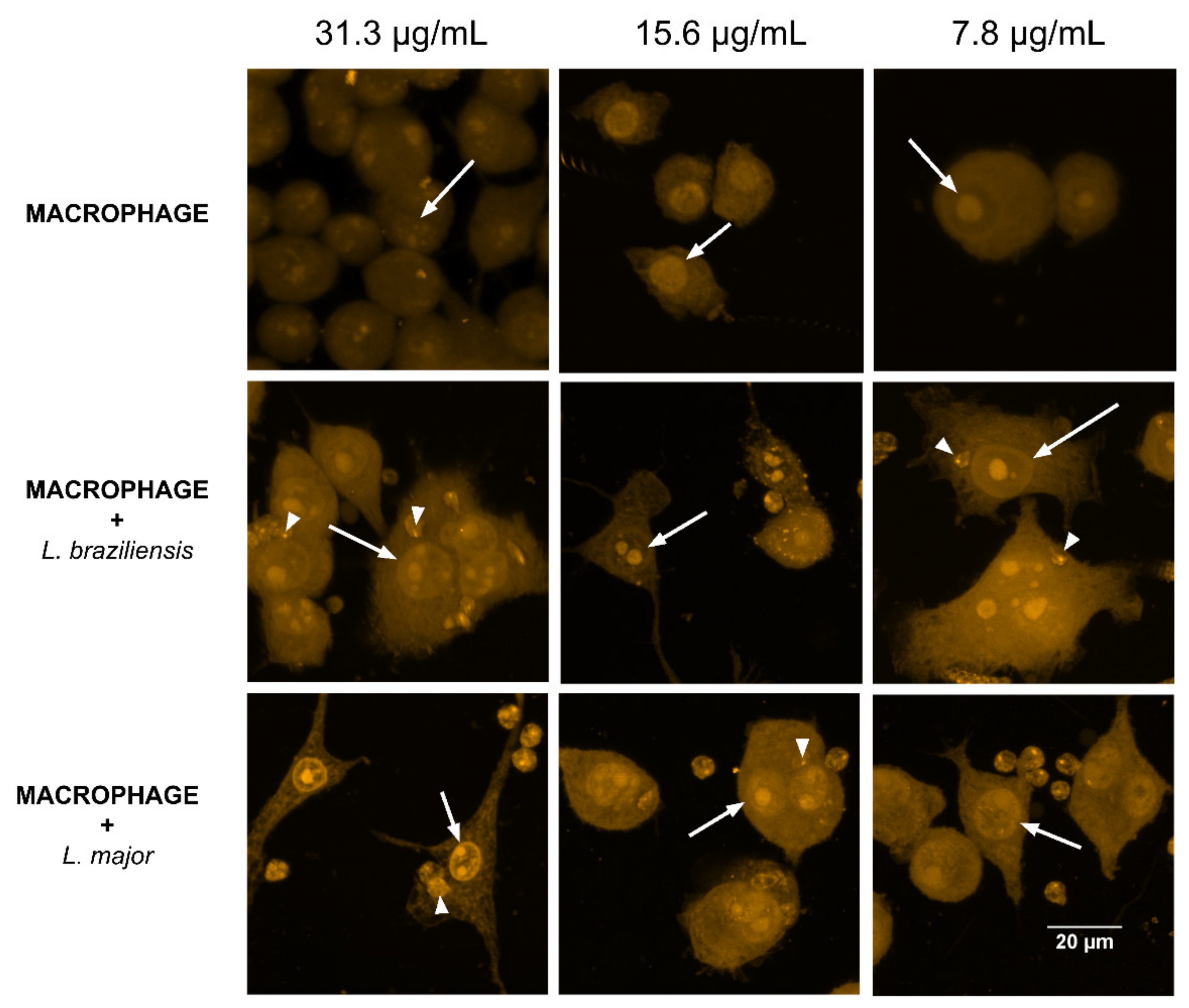
2.3. Internalization of Curcumin in Isolated and Leishmania-Infected Macrophages
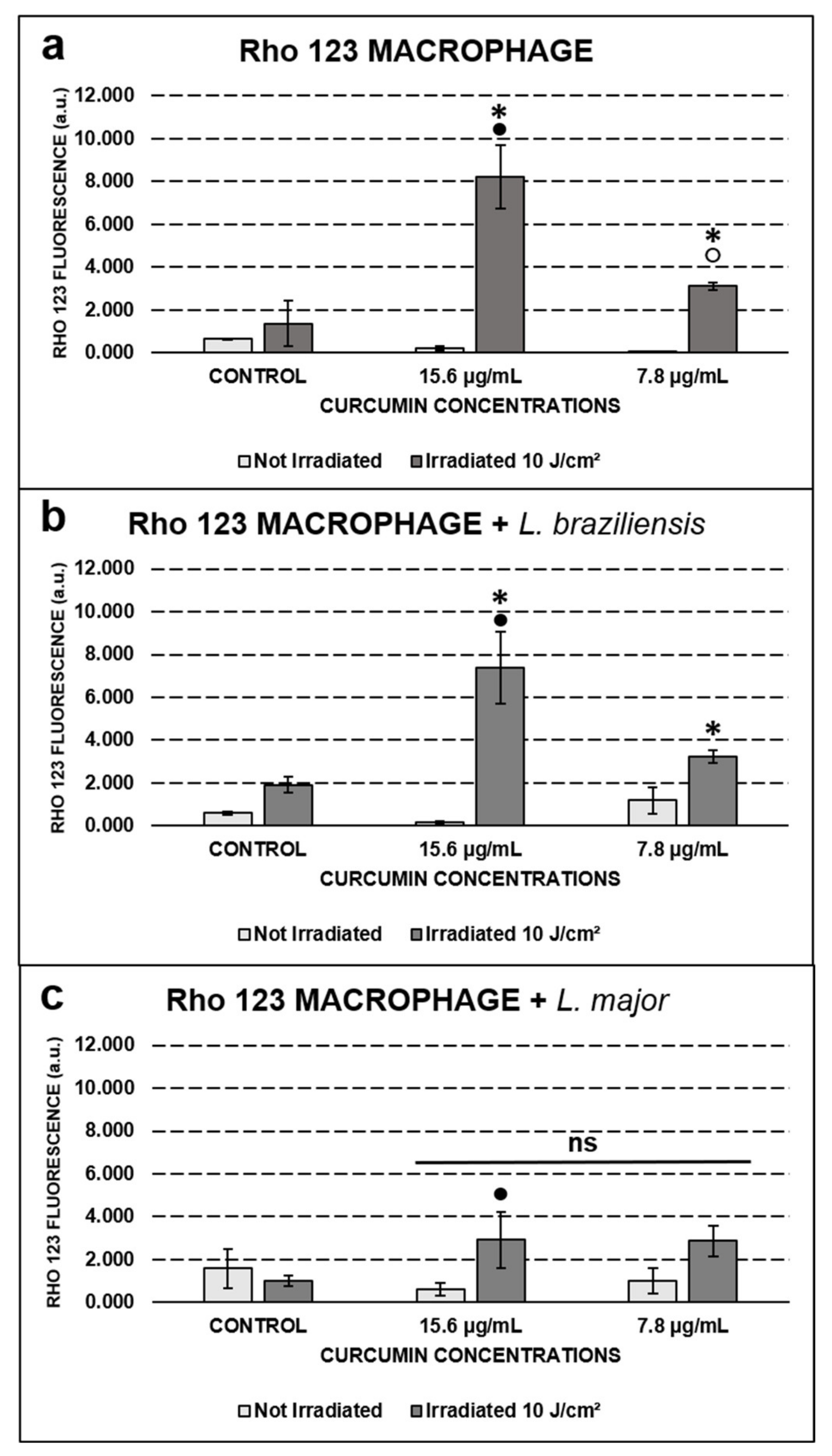
2.4. Qualitative and Quantitative Assessment of Mitochondrial Activity by Rho 123
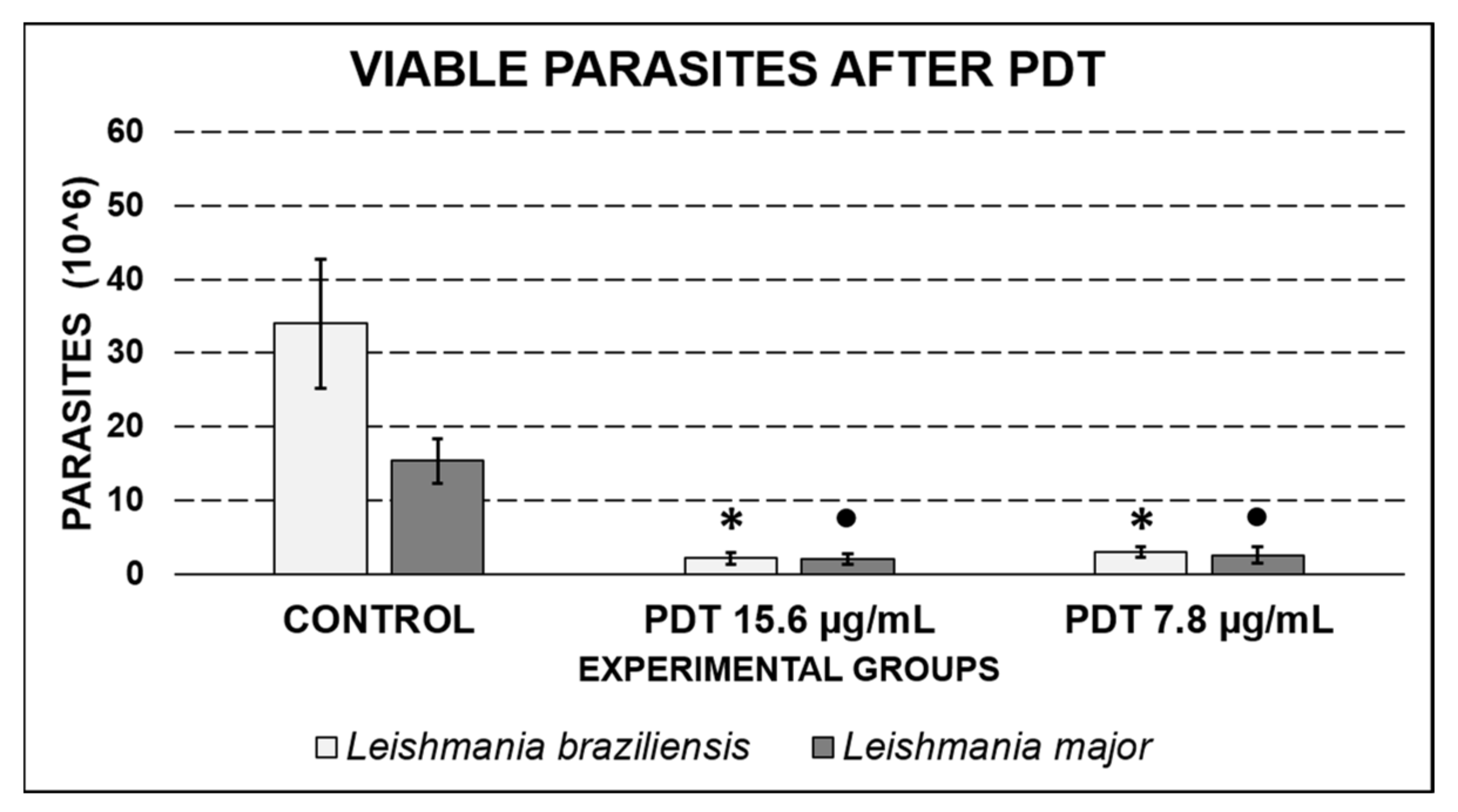
2.5. Retrieval of Leishmania after PDT Exposure
3. Discussion
4. Materials and Methods
4.1. Parasite Culture
4.2. Cultivation of Macrophages
4.3. Macrophage Infection
4.4. Curcumin Preparation
4.5. Photodynamic Therapy
4.6. Viability Assessed by the Trypan Blue Exclusion Test
4.7. Morphological Evaluation by Scanning Electron Microscopy
4.8. Curcumin Internalization in Macrophages Isolated and Infected with Leishmania
4.9. Evaluation of Mitochondrial Activity by Rhodamine 123 Fluorescent Staining
4.10. Retrieval of Leishmania after PDT
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alvar, J.; Yactayo, S.; Bern, C. Leishmaniasis and poverty. Trends Parasitol. 2006, 22, 552–557. [Google Scholar] [CrossRef]
- Aronson, N.; Herwaldt, B.L.; Libman, M.; Pearson, R.; Lopez-velez, R.; Weina, P.; Carvalho, E.M.; Ephros, M.; Jeronimo, S.; Magill, A. Diagnosis and Treatment of Leishmaniasis: Clinical Practice Guidelines by the Infectious Diseases Society of America ( IDSA ) and the American Society of Tropical Medicine and Hygiene ( ASTMH ). Clin. Infect. Dis. 2016, 63, e202–e264. [Google Scholar] [CrossRef]
- Von Stebut, E. Leishmaniasis. J. Ger. Soc. Dermatol. 2015, 191–201. [Google Scholar] [CrossRef]
- Abadir, A.; Patel, A.; Haider, S. Systemic therapy of New World cutaneous leishmaniasis: A case report and review article. Can. J. Infect. Dis. Med. Microbiol. 2010, 21, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatami, A.; Firooz, A.; Gorouhi, F.; Dowlati, Y.; Arabia, S. Treatment of acute Old World cutaneous leishmaniasis: A systematic review of the randomized controlled trials. J. Am. Acad. Dermatol. 2007, 57, 335e1–335e29. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.S.; Wainwright, M. Photodynamic antimicrobial chemotherapy (PACT) for the treatment of malaria, leishmaniasis and trypanosomiasis. Braz. J. Med. Biol. Res. 2011, 44, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Snoek, E.M.; Robinson, D.J.; Van Hellemond, J.J.; Neumann, H.A.M. A review of photodynamic therapy in cutaneous leishmaniasis. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 918–922. [Google Scholar] [CrossRef]
- Slape, D.R.-M.; Kim, E.N.-Y.; Weller, P.; Gupta, M. Leishmania tropica successfully treated with photodynamic therapy. Australas. J. Dermatol. 2019, 60, e64–e65. [Google Scholar] [CrossRef]
- Goldin, H.; Kohen, S.; Taxy, J.; Libman, M.; Cibull, T.; Billick, K. Leishmania tropica infection of the ear treated with photodynamic therapy. JAAD Case Rep. 2020, 6, 514–517. [Google Scholar] [CrossRef]
- Khan, A.; Khan, K.; Biradar, G.; Khan, M. Treatment of Cutaneous Leishmaniasis with Photodynamic Therapy: The First Case Report from Khyber Pakhthunkhwa (KPK), Swat Pakistan. Clin. Dermatol. Open Access J. 2020, 5, 120–124. [Google Scholar] [CrossRef]
- Sunyoto, T.; Boelaert, M.; Meheus, F. Understanding the economic impact of leishmaniasis on households in endemic countries: A systematic review. Expert Rev. Anti Infect. Ther. 2019, 17, 57–69. [Google Scholar] [CrossRef]
- Nistico, S.; Tamburi, F.; Bennardo, L.; Dastoli, S.; Schipani, G.; Caro, G.; Fortuna, M.C.; Rossi, A. Treatment of telogen effluvium using a dietary supplement containing Boswellia serrata, Curcuma longa, and Vitis vinifera: Results of an observational study. Dermatol. Ther. 2019, 32, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Okwor, I.; Uzonna, J. Social and economic burden of human leishmaniasis. Am. J. Trop. Med. Hyg. 2016, 94, 489–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, S.; João, E.; Monteiro, J.; Neto, M.B.; Pinheiro, A. Leishmanicidal effect of antiparasitic photodynamic therapy—ApPDT on infected macrophages. Lasers Med. Sci. 2017, 32, 1959–1964. [Google Scholar] [CrossRef]
- Aureliano, D.P.; Ribeiro, M.S.; Angelo, J.; Lindoso, L.; Pogliani, F.C.; Sellera, F.P.; Song, D.; Baptista, M.S. Treatment and Control of Leishmaniasis Using Photodynamic Therapy. In Leishmaniasis-Trends in Epidemiology, Diagnosis and Treatment [Internet]; Intech: Rijeka, Croatia; pp. 393–412.
- Pinto, J.G.; de Martins, J.F.S.; Pereira, A.H.C.; Mittmann, J.; Raniero, L.J.; Ferreira-Strixino, J. Evaluation of methylene blue as photosensitizer in promastigotes of Leishmania major and Leishmania braziliensis. Photodiagn. Photodyn. Ther. 2017, 18, 325–330. [Google Scholar] [CrossRef]
- Song, D.; Lindoso, J.A.L.; Oyafuso, L.K.; Kanashiro, E.H.Y.; Cardoso, J.L.; Uchoa, A.F.; Tardivo, J.P.; Baptista, M.S. Photodynamic therapy using methylene blue to treat cutaneous leishmaniasis. Photomed. Laser Surg. 2011, 29, 711–715. [Google Scholar] [CrossRef]
- De Silva, E.P.O.; Mittmann, J.; Ferreira, V.T.P.; Cardoso, M.A.G.; Beltrame, M. Photodynamic effects of zinc phthalocyanines on intracellular amastigotes of Leishmania amazonensis and Leishmania braziliensis. Lasers Med. Sci. 2015, 30, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Nesi-reis, V.; Navasconi, T.R.; Lera-nosone, D.S.S.L.; Oliveira, E.L.; Barbosa, P.M.; Caetano, W.; Silveira, T.G.V.; Aristides, S.M.A.; Hioka, N.; Lonardoni, M.V.C. Photodiagnosis and Photodynamic Therapy Phototoxic effect of aluminium-chlorine and aluminium-hydroxide phthalocyanines on Leishmania (l.) amazonensis. Photodiagn. Photodyn. Ther. 2018, 21, 239–245. [Google Scholar] [CrossRef]
- Pinto, J.G.; Pacheco-Soares, C.; Mittmann, J. Assessment of Leishmania major and Leishmania braziliensis promastigote viability after photodynamic treatment with aluminum phthalocyanine tetrasulfonate (AlPcS4). J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 300–307. [Google Scholar]
- Andrade, C.G.; Figueiredo, R.C.B.Q.; Ribeiro, K.R.C.; Souza, L.I.O.; Santos, B.S.; Ribeiro, M.S.; Fontes, A. Photodynamic effect of zinc porphyrin on the promastigote and amastigote forms of Leishmania braziliensis. Photochem. Photobiol. Sci. 2018, 17, 482–490. [Google Scholar] [CrossRef]
- Pinto, J.G.; Marcolino, L.M.C.; Ferreira-Strixino, J. Photodynamic activity of Photogem® in Leishmania promastigotes and infected macrophages. Future Microbiol. 2021, 16, 95–106. [Google Scholar] [CrossRef]
- Pinto, J.G.; Fontana, L.C.; de Oliveira, M.A.; Kurachi, C.; Raniero, L.J.; Ferreira-Strixino, J. In vitro evaluation of photodynamic therapy using curcumin on Leishmania major and Leishmania braziliensis. Lasers Med. Sci. 2016, 31, 883–890. [Google Scholar] [CrossRef] [PubMed]
- De Souza, C.; Carvalho, J.A.; Abreu, A.S.; De, L.P.; Ambrósio, J.A.R.; Junior, M.B.; de Oliveira, M.A.; Mittmann, J.; Simioni, A.R.; de Souza, C.; et al. Polyelectrolytic gelatin nanoparticles as a drug delivery system for the promastigote form of Leishmania amazonensis treatment. J. Biomater. Sci. Polym. Ed. 2021, 32, 1–21. [Google Scholar] [CrossRef]
- Pinto, J.G.; Pereira, A.H.C.; de Oliveira, M.A.; Kurachi, C.; Raniero, L.J.; Ferreira-Strixino, J. Chlorin E6 phototoxicity in L. major and L. braziliensis promastigotes—In vitro study. Photodiagnosis Photodyn. Ther. 2016, 15, 19–24. [Google Scholar] [CrossRef]
- Awad, M.M.; Tovmasyan, A.; Craik, J.D.; Batinic-Haberle, I.; Benov, L.T. Important cellular targets for antimicrobial photodynamic therapy. Appl. Microbiol. Biotechnol. 2016, 100, 7679–7688. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.S.; Turchiello, R.; Kowaltowski, A.J.; Indig, G.L.; Baptista, M.S. Major determinants of photoinduced cell death: Subcellular localization versus photosensitization efficiency. Free Radic. Biol. Med. 2011, 51, 824–833. [Google Scholar] [CrossRef]
- Ogilby, P.R. Singlet oxygen: There is indeed something new under the sun. Chem. Soc. Rev. 2010, 39, 3181–3209. [Google Scholar] [CrossRef]
- Nafisi, S.; Adelzadeh, M.; Norouzi, Z.; Sarbolouki, M.N. Curcumin Binding to DNA and RNA. DNA Cell Biol. 2009, 28, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Prasad, S.; Kim, H.; Patchva, S.; Webb, L.J.; Priyadarsini, K.; Aggarwal, B.B. Multitargeting by curcumin as revealed by molecular interaction studies. Nat. Prod. Rep. 2011, 28, 1937–1955. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Kumar, G.S. Biophysical studies on curcumin—Deoxyribonucleic acid interaction: Spectroscopic and calorimetric approach. Int. J. Biol. Macromol. 2013, 62, 257–264. [Google Scholar] [CrossRef]
- Di Lisa, F.; Bernardi, P. Mitochondrial function as a determinant of recovery or death in cell response to injury. Mol. Cell. Biochem. 1998, 184, 379–391. [Google Scholar] [CrossRef]
- Petit, P.X.; O’Connor, J.E.; Grunwald, D.; Brown, S.C. Analysis of the membrane potential of rat- and mouse-liver mitochondria by flow cytometry and possible applications. Eur. J. Biochem. 1990, 194, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Camara, A.K.S.; Stowe, D.F.; Qi, F.; Beard, D.A. Mitochondrial inner membrane electrophysiology assessed by rhodamine-123 transport and fluorescence. Ann. Biomed. Eng. 2007, 35, 1276–1285. [Google Scholar] [CrossRef] [PubMed]
- Fedyaeva, A.V.; Stepanov, A.V.; Lyubushkina, I.V.; Pobezhimova, T.P.; Rikhvanov, E.G. Heat Shock Induces Production of Reactive Oxygen Species and Increases Inner Mitochondrial Membrane Potential in Winter Wheat Cells. Russ. Biokhimiya 2014, 79, 1202–1210. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasbihi, M.; Shekari, F.; Hajjaran, H.; Khanmohammadi, M.; Hadighi, R. Comparative mitochondrial proteomics of Leishmania tropica clinical isolates resistant and sensitive to meglumine antimoniate. Parasitol. Res. 2020, 119, 1857–1871. [Google Scholar] [CrossRef]
- Yue, J.; Shen, Y.; Liang, L.; Cong, L.; Xu, W.; Shi, W.; Liang, C.; Xu, S. Revealing Mitochondrial Microenvironmental Evolution Triggered by Photodynamic Therapy. Anal. Chem. 2020, 92, 6081–6087. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, A.H.C.; Marcolino, L.M.C.; Pinto, J.G.; Ferreira-Strixino, J. Evaluation of the Photodynamic Therapy with Curcumin on L. braziliensis and L. major Amastigotes. Antibiotics 2021, 10, 634. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060634
Pereira AHC, Marcolino LMC, Pinto JG, Ferreira-Strixino J. Evaluation of the Photodynamic Therapy with Curcumin on L. braziliensis and L. major Amastigotes. Antibiotics. 2021; 10(6):634. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060634
Chicago/Turabian StylePereira, André Henrique Correia, Luciana Maria Cortez Marcolino, Juliana Guerra Pinto, and Juliana Ferreira-Strixino. 2021. "Evaluation of the Photodynamic Therapy with Curcumin on L. braziliensis and L. major Amastigotes" Antibiotics 10, no. 6: 634. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060634