
Overexpression of the MexXY Multidrug Efflux System Correlates with Deficient Pyoverdine Production in Pseudomonas aeruginosa


Abstract
:1. Introduction
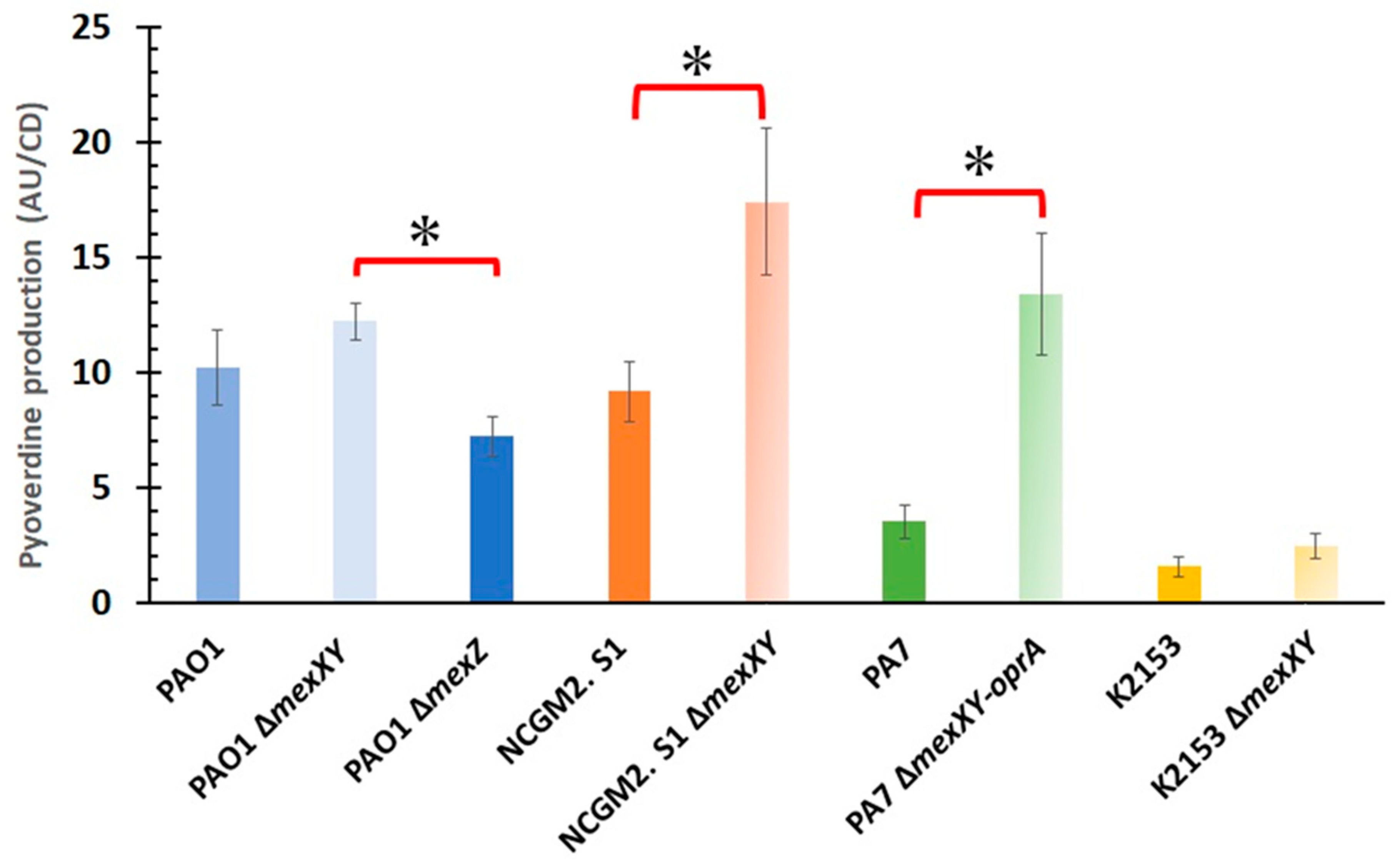
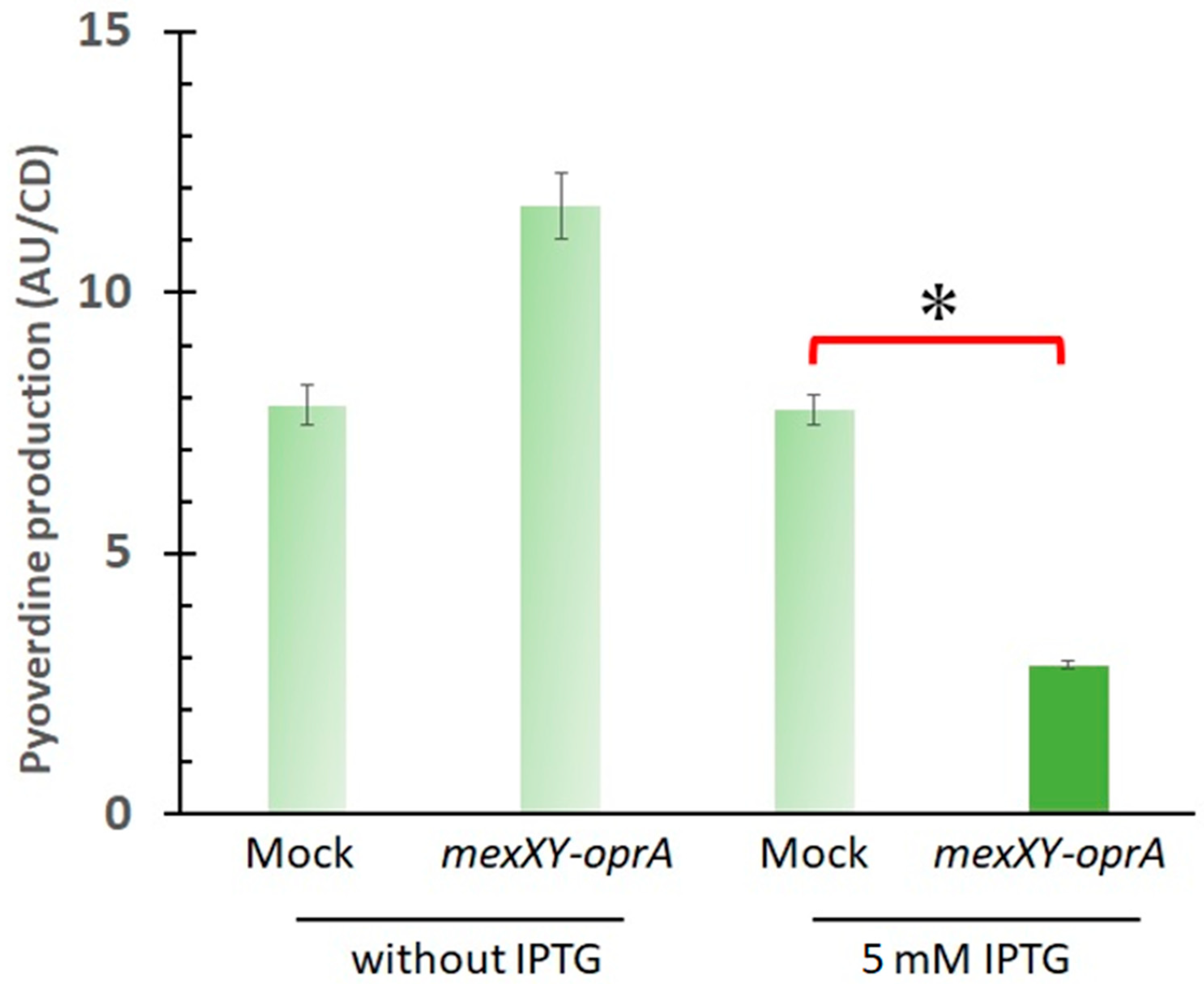
2. Results
3. Discussion
4. Materials and Methods
4.1. Pseudomonas aeruginosa Strains and Growth Conditions
4.2. Assay of Pyoverdine Production
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gellatly, S.L.; Hancock, R.E. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Poole, K. Pseudomonas aeruginosa: Resistance to the Max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, Y.; Tomida, J.; Kawamura, Y. Resistance and Response to Anti-Pseudomonas Agents and Biocides. In Pseudomonas, 1st ed.; Ramos, J.L., Goldberg, J., Filloux, A., Eds.; Springer Science and Business Media: Dordrecht, The Netherlands, 2014; pp. 173–187. [Google Scholar]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The Challenge of Efflux-Mediated Antibiotic Resistance in Gram-Negative Bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [Green Version]
- Morita, Y.; Tomida, J.; Kawamura, Y. MexXY multidrug efflux system of Pseudomonas aeruginosa. Front. Microbiol. 2012, 3, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puja, H.; Bolard, A.; Noguès, A.; Plésiat, P.; Jeannot, K. The Efflux Pump MexXY/OprM Contributes to the Tolerance and Acquired Resistance of Pseudomonas aeruginosa to Colistin. Antimicrob. Agents Chemother. 2020, 64, e02033-19. [Google Scholar] [CrossRef]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef]
- Cabot, G.; Ocampo-Sosa, A.A.; Domínguez, M.A.; Gago, J.F.; Juan, C.; Tubau, F.; Rodríguez, C.; Moyà, B.; Peña, C.; Martínez, L.M.; et al. Genetic Markers of Widespread Extensively Drug-Resistant Pseudomonas aeruginosa High-Risk Clones. Antimicrob. Agents Chemother. 2012, 56, 6349–6357. [Google Scholar] [CrossRef] [Green Version]
- Mulet, X.; Cabot, G.; Ocampo-Sosa, A.A.; Domínguez, M.A.; Zamorano, L.; Juan, C.; Tubau, F.; Rodríguez, C.; Moyà, B.; Peña, C.; et al. Biological Markers of Pseudomonas aeruginosa Epidemic High-Risk Clones. Antimicrob. Agents Chemother. 2013, 57, 5527–5535. [Google Scholar] [CrossRef] [Green Version]
- Cornelis, P.; Dingemans, J. Pseudomonas aeruginosa adapts its iron uptake strategies in function of the type of infections. Front. Cell. Infect. Microbiol. 2013, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Hwang, W.; Yoon, S.S. Virulence Characteristics and an Action Mode of Antibiotic Resistance in Multidrug-Resistant Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, Y.; Tomida, J.; Kawamura, Y. Primary mechanisms mediating aminoglycoside resistance in the multidrug-resistant Pseudomonas aeruginosa clinical isolate PA7. Microbiology 2012, 158, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.H.; Tetu, S.G.; Larouche, A.; Elbourne, L.; Tremblay, S.; Ren, Q.; Dodson, R.; Harkins, D.; Shay, R.; Watkins, K.; et al. Complete Genome Sequence of the Multiresistant Taxonomic Outlier Pseudomonas aeruginosa PA7. PLoS ONE 2010, 5, e8842. [Google Scholar] [CrossRef]
- Imperi, F.; Putignani, L.; Tiburzi, F.; Ambrosi, C.; Cipollone, R.; Ascenzi, P.; Visca, P. Membrane-association determinants of the ω-amino acid monooxygenase PvdA, a pyoverdine biosynthetic enzyme from Pseudomonas aeruginosa. Microbiology 2008, 154, 2804–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobel, M.L.; McKay, G.A.; Poole, K. Contribution of the MexXY Multidrug Transporter to Aminoglycoside Resistance in Pseudomonas aeruginosa Clinical Isolates. Antimicrob. Agents Chemother. 2003, 47, 3202–3207. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi-Akiyama, T.; Kuwahara, T.; Tada, T.; Kitao, T.; Kirikae, T. Complete Genome Sequence of Highly Multidrug-Resistant Pseudomonas aeruginosa NCGM2.S1, a Representative Strain of a Cluster Endemic to Japan. J. Bacteriol. 2011, 193, 7010. [Google Scholar] [CrossRef] [Green Version]
- Morita, Y.; Sobel, M.L.; Poole, K. Antibiotic Inducibility of the MexXY Multidrug Efflux System of Pseudomonas aeruginosa: Involvement of the Antibiotic-Inducible PA5471 Gene Product. J. Bacteriol. 2006, 188, 1847–1855. [Google Scholar] [CrossRef] [Green Version]
- Guénard, S.; Muller, C.; Monlezun, L.; Benas, P.; Broutin, I.; Jeannot, K.; Plésiat, P. Multiple Mutations Lead to MexXY-OprM-Dependent Aminoglycoside Resistance in Clinical Strains of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 58, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Morita, Y.; Nakashima, K.-I.; Nishino, K.; Kotani, K.; Tomida, J.; Inoue, M.; Kawamura, Y. Berberine Is a Novel Type Efflux Inhibitor Which Attenuates the MexXY-Mediated Aminoglycoside Resistance in Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 1223. [Google Scholar] [CrossRef] [PubMed]
- Ringel, M.T.; Brüser, T. The biosynthesis of pyoverdines. Microb. Cell 2018, 5, 424–437. [Google Scholar] [CrossRef]
- Fraud, S.; Poole, K. Oxidative Stress Induction of the MexXY Multidrug Efflux Genes and Promotion of Aminoglycoside Resistance Development in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2010, 55, 1068–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, M.; DeLuca, D.; Worgall, S.; Quadri, L.E.N. Transcriptome Analysis of the Response of Pseudomonas aeruginosa to Hydrogen Peroxide. J. Bacteriol. 2004, 186, 248–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Genet. 2013, 11, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Lab Stock | Strain (=Co-Identical Strain) * | Reference |
|---|---|---|
| IMPU 1 | PAO1 (=K767 or PAGU 974) | [15] |
| IMPU 2 | NCGM2. S1 (=PAGU 1606) | [16] |
| IMPU 9 | PAO1 ΔmexXY (=K1525, PAGU 975) | [15] |
| IMPU 10 | NCGM2. S1 ΔmexXY (=PAGUg1659) | [12] |
| IMPU 17 | PAO1 ΔmexZ (=K2415, PAGUg1659) | [17] |
| IMPU 21 | PA7 (=PAGU 1498) | [13] |
| IMPU 29 | K2153 (=PAGU 1741) | [15] |
| IMPU 44 | PA7 ΔmexXY-oprA (=PAGUg1565) | [12] |
| IMPU 45 | K2153 ΔmexXY (=PAGUg1857) | [15] |
| IMPU 53 | PA7 ΔmexXY-oprA attB::pYM101 (=PAGUg1632) | [12] |
| IMPU 54 | PA7 ΔmexXY-oprA attB::pYM101-mexXY-oprA (=PAGUg1633) | [12] |
| IMPU 61 | PAO1 | [14] |
| IMPU 62 | PAO1ΔpvdA | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikarashi, K.; Kutsuna, R.; Tomida, J.; Kawamura, Y.; Morita, Y. Overexpression of the MexXY Multidrug Efflux System Correlates with Deficient Pyoverdine Production in Pseudomonas aeruginosa. Antibiotics 2021, 10, 658. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060658
Ikarashi K, Kutsuna R, Tomida J, Kawamura Y, Morita Y. Overexpression of the MexXY Multidrug Efflux System Correlates with Deficient Pyoverdine Production in Pseudomonas aeruginosa. Antibiotics. 2021; 10(6):658. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060658
Chicago/Turabian StyleIkarashi, Kei, Ryo Kutsuna, Junko Tomida, Yoshiaki Kawamura, and Yuji Morita. 2021. "Overexpression of the MexXY Multidrug Efflux System Correlates with Deficient Pyoverdine Production in Pseudomonas aeruginosa" Antibiotics 10, no. 6: 658. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060658