
Elucidating the Antimycobacterial Mechanism of Action of Decoquinate Derivative RMB041 Using Metabolomics

 , , and
, , and
Abstract
:1. Introduction
2. Results
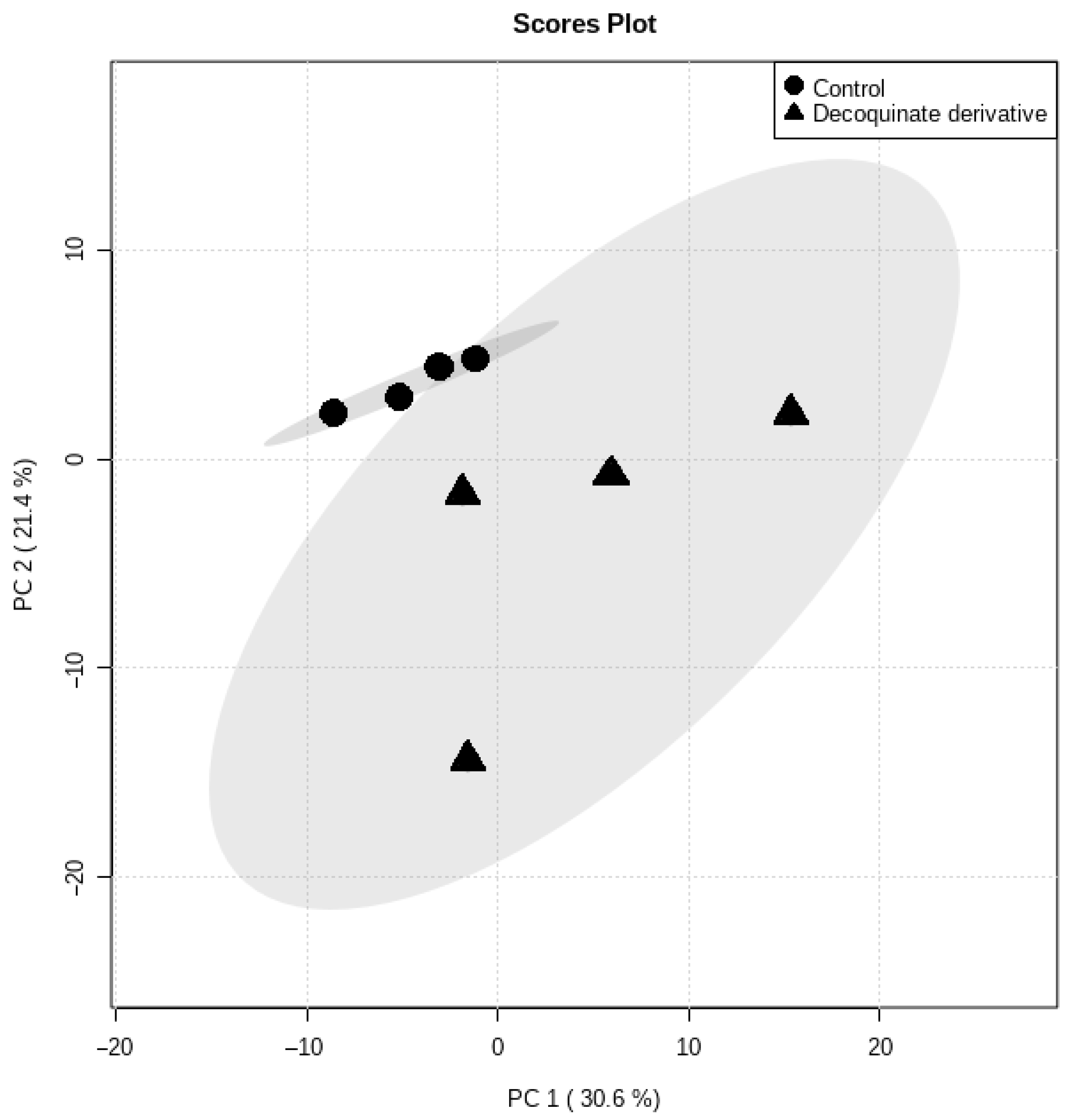
2.1. Data Overview
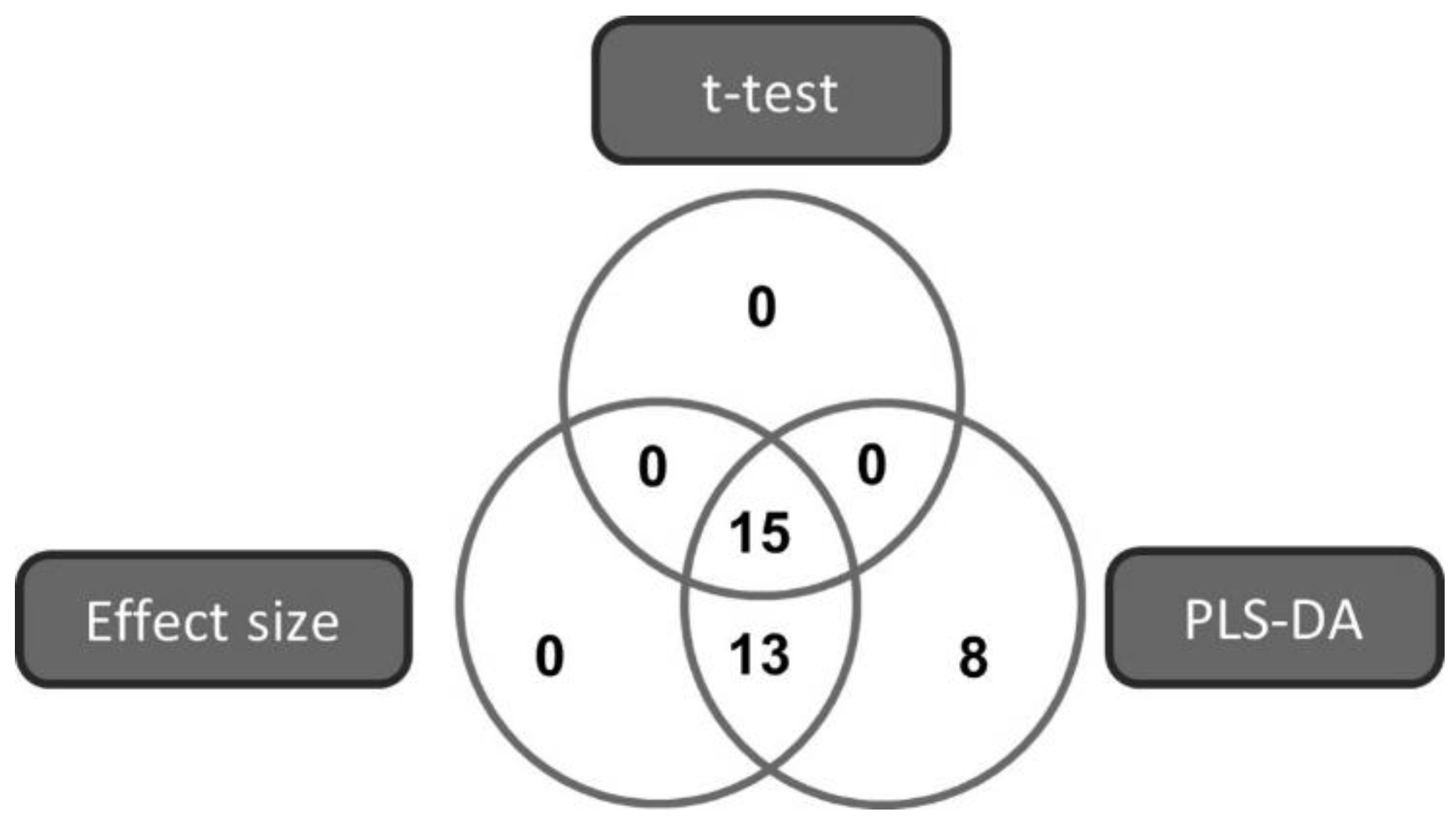
2.2. Marker Selection
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Whole Metabolome Extraction Procedure and Derivatization
4.3. GCxGC-TOFMS Analysis
4.4. Data Processing, Clean-Up, and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harding, E. WHO global progress report on tuberculosis elimination. Lancet Respir. Med. 2020, 8, 19. [Google Scholar] [CrossRef]
- Tanner, L.; Haynes, R.K.; Wiesner, L. An in vitro ADME and in vivo Pharmacokinetic Study of Novel TB-Active Decoquinate Derivatives. Front. Pharmacol. 2019, 10, 120. [Google Scholar] [CrossRef]
- Nguyen, T.V.A.; Anthony, R.M.; Bañuls, A.-L.; Nguyen, T.V.A.; Vu, D.H.; Alffenaar, J.-W.C. Bedaquiline resistance: Its emergence, mechanism, and prevention. Clin. Infect. Dis. 2018, 66, 1625–1630. [Google Scholar] [CrossRef]
- Polsfuss, S.; Hofmann-Thiel, S.; Merker, M.; Krieger, D.; Niemann, S.; Rüssmann, H.; Schönfeld, N.; Hoffmann, H.; Kranzer, K. Emergence of low-level delamanid and bedaquiline resistance during extremely drug-resistant tuberculosis treatment. Clin. Infect. Dis. 2019, 69, 1229–1231. [Google Scholar] [CrossRef]
- World Health Organization. Consolidated Guidelines on Drug-Resistant Tuberculosis Treatment; World Health Assembly: Geneva, Switzerland, 2019. [Google Scholar]
- Da Silva, P.E.A.; Palomino, J.C. Molecular basis and mechanisms of drug resistance in Mycobacterium tuberculosis: Classical and new drugs. J. Antimicrob. Chemother. 2011, 66, 1417–1430. [Google Scholar] [CrossRef]
- Holmes, K.K.; Bertozzi, S.; Bloom, B.R.; Jha, P. Major Infectious Diseases: Key Messages in Disease Control Priorities, 3rd ed.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2017; Chapter 1. [Google Scholar]
- Vernon, A.; Fielding, K.; Savic, R.; Dodd, L.; Nahid, P. The importance of adherence in tuberculosis treatment clinical trials and its relevance in explanatory and pragmatic trials. PLoS Med. 2019, 16, e1002884. [Google Scholar] [CrossRef]
- Migliori, G.B.; Thong, P.M.; Akkerman, O.; Alffenaar, J.-W.; Álvarez-Navascués, F.; Assao-Neino, M.M.; Bernard, P.V.; Biala, J.S.; Blanc, F.-X.; Bogorodskaya, E.M. Worldwide effects of coronavirus disease pandemic on tuberculosis services, January–April 2020. Emerg. Infect. Dis. 2020, 26, 2709. [Google Scholar] [CrossRef] [PubMed]
- Duarte, R.; Lönnroth, K.; Carvalho, C.; Lima, F.; Carvalho, A.; Muñoz-Torrico, M.; Centis, R. Tuberculosis, social determinants and co-morbidities (including HIV). Pulmonology 2018, 24, 115–119. [Google Scholar] [CrossRef]
- Watermeyer, J.; Penn, C. Community perspectives on tuberculosis care in rural South Africa. HSCC 2019, 27, 182–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuaid, C.F.; McCreesh, N.; Read, J.M.; Sumner, T.; Houben, R.M.; White, R.G.; Harris, R.C.; Group, C.C.-W. The potential impact of COVID-19-related disruption on tuberculosis burden. Eur. Respir. J. 2020, 56, 2001718. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Critically Important Antimicrobials for Human Medicine, 6th ed.; World Health Assembly: Geneva, Switzerland, 2019. [Google Scholar]
- Sloan, D.J.; Lewis, J.M. Management of multidrug-resistant TB: Novel treatments and their expansion to low resource settings. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Kolyva, A.S.; Karakousis, P.C. Old and new TB drugs: Mechanisms of action and resistance. In Understanding Tuberculosis—New Approaches to Fighting against Drug Resistance; InTech: Plantation, FL, USA, 2012; pp. 209–232. [Google Scholar]
- Ginsberg, A.M.; Spigelman, M. Challenges in tuberculosis drug research and development. Nat. Med. 2007, 13, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Beteck, R.M.; Seldon, R.; Jordaan, A.; Warner, D.F.; Hoppe, H.C.; Laming, D.; Khanye, S.D. New quinolone-based thiosemicarbazones showing activity against Plasmodium falciparum and Mycobacterium tuberculosis. Molecules 2019, 24, 1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLoughlin, D.; Chute, M. Efficacy of decoquinate against eleven strains of Eimeria tenella and development of a decoquinate-resistant strain. Avian Dis. 1971, 15, 342–345. [Google Scholar] [CrossRef]
- Beteck, R.M.; Coertzen, D.; Smit, F.J.; Birkholtz, L.M.; Haynes, R.K.; N’Da, D.D. Straightforward conversion of decoquinate into inexpensive tractable new derivatives with significant antimalarial activities. Bioorg. Med. Chem. Lett. 2016, 26, 3006–3009. [Google Scholar] [CrossRef] [Green Version]
- Ginsburg, A.S.; Grosset, J.H.; Bishai, W.R. Fluoroquinolones, tuberculosis, and resistance. Lancet Infect. Dis. 2003, 3, 432–442. [Google Scholar] [CrossRef]
- Wohnsland, F.; Faller, B. High-throughput permeability pH profile and high-throughput alkane/water log P with artificial membranes. J. Med. Chem. 2001, 44, 923–930. [Google Scholar] [CrossRef]
- Nam, J.; Beales, P.A.; Vanderlick, T.K. Giant phospholipid/block copolymer hybrid vesicles: Mixing behavior and domain formation. Langmuir 2011, 27, 1–6. [Google Scholar] [CrossRef]
- Tanner, L.; Haynes, R.K.; Wiesner, L. Accumulation of TB-Active Compounds in Murine Organs Relevant to Infection by Mycobacterium tuberculosis. Front. Pharmacol. 2020, 11, 724. [Google Scholar] [CrossRef] [PubMed]
- Lenaerts, A.; Barry, C.E., III; Dartois, V. Heterogeneity in tuberculosis pathology, microenvironments and therapeutic responses. Immunol. Rev. 2015, 264, 288–307. [Google Scholar] [CrossRef] [Green Version]
- Beteck, R.M.; Seldon, R.; Coertzen, D.; van der Watt, M.E.; Reader, J.; Mackenzie, J.S.; Lamprecht, D.A.; Abraham, M.; Eribez, K.; Müller, J.; et al. Accessible and distinct decoquinate derivatives active against Mycobacterium tuberculosis and apicomplexan parasites. Commun. Chem. 2018, 1, 1–11. [Google Scholar] [CrossRef]
- Lyons, M.A.; Reisfeld, B.; Yang, R.S.; Lenaerts, A.J. A physiologically based pharmacokinetic model of rifampin in mice. Antimicrob. Agents Chemother. 2013, 57, 1763–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, B.; Truffot-Pernot, C.; Lacroix, C.; Raviglione, M.C.; O’Brien, R.J.; Olliaro, P.; Roscigno, G.; Grosset, J. Effectiveness of rifampin, rifabutin and rifapentine for preventive therapy of tuberculosis in mice. Am. J. Respir. Crit. Care Med. 1993, 148, 1541–1546. [Google Scholar] [CrossRef] [PubMed]
- Arbex, M.A.; Varella, M.D.C.L.; Siqueira, H.R.D.; Mello, F.A.F.D. Antituberculosis drugs: Drug interactions, adverse effects, and use in special situations-part 2: Second line drugs. J. Bras. Pneum. 2010, 36, 641–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaram, R.; Shandil, R.K.; Gaonkar, S.; Kaur, P.; Suresh, B.; Mahesh, B.; Jayashree, R.; Nandi, V.; Bharath, S.; Kantharaj, E. Isoniazid pharmacokinetics-pharmacodynamics in an aerosol infection model of tuberculosis. Antimicrob. Agents Chemother. 2004, 48, 2951–2957. [Google Scholar] [CrossRef] [Green Version]
- Grosset, J.; Ji, B. Experimental chemotherapy of mycobacterial diseases. In Mycobacteria; Springer: Berlin/Heidelberg, Germany, 1998; pp. 51–97. [Google Scholar]
- Lakshminarayana, S.B.; Huat, T.B.; Ho, P.C.; Manjunatha, U.H.; Dartois, V.; Dick, T.; Rao, S.P. Comprehensive physicochemical, pharmacokinetic and activity profiling of anti-TB agents. J. Antimicrob. Chemother. 2015, 70, 857–867. [Google Scholar] [CrossRef]
- Dheda, K.; Gumbo, T.; Gandhi, N.R.; Murray, M.; Theron, G.; Udwadia, Z.; Migliori, G.B.; Warren, R. Global control of tuberculosis: From extensively drug-resistant to untreatable tuberculosis. Lancet Respir. Med. 2014, 2, 321–338. [Google Scholar] [CrossRef] [Green Version]
- Warner, D.F.; Mizrahi, V. Shortening treatment for tuberculosis--to basics. N. Engl. J. Med. 2014, 371, 1642–1643. [Google Scholar] [CrossRef]
- Cho, H.-W.; Kim, S.B.; Jeong, M.K.; Park, Y.; Miller, N.; Ziegler, T.; Jones, D. Discovery of metabolite features for the modelling and analysis of high-resolution NMR spectra. Int. J. Data Min. Bioinform. 2008, 2, 176–192. [Google Scholar] [CrossRef]
- Byliński, H.; Dymerski, T.; Gębicki, J.; Namieśnik, J. Complementary use of GCxGC–TOF–MS and statistics for differentiation of variety in biosolid samples. Monatshefte Chem. 2018, 149, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, G.M.; Feinn, R. Using effect size—Or why the P value is not enough. J. Grad. Med. Educ. 2012, 4, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, I.; Rohilla, A.; Khuller, G.K. Alterations in macromolecular composition and cell wall integrity by ciprofloxacin in Mycobacterium smegmatis. Lett. Appl. Microbiol. 1999, 29, 113–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, D.; Bozic, C.M.; McNeil, M.; Brennan, P.J. Structural features of the arabinan component of the lipoarabinomannan of Mycobacterium tuberculosis. J. Biol. Chem. 1991, 266, 9652–9660. [Google Scholar] [CrossRef]
- Jankute, M.; Cox, J.A.; Harrison, J.; Besra, G.S. Assembly of the Mycobacterial Cell Wall. Annu. Rev. Microbiol. 2015, 69, 405–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Matsumura, T.; Ato, M.; Hamasaki, M.; Nishiuchi, Y.; Murakami, Y.; Maeda, Y.; Yoshimori, T.; Matsumoto, S.; Kobayashi, K. Critical roles for lipomannan and lipoarabinomannan in cell wall integrity of mycobacteria and pathogenesis of tuberculosis. MBio 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal-Mutalik, R.; Nikaido, H. Mycobacterial outer membrane is a lipid bilayer and the inner membrane is unusually rich in diacyl phosphatidylinositol dimannosides. Proc. Natl. Acad. Sci. USA 2014, 111, 4958–4963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Movahedzadeh, F.; Wheeler, P.R.; Dinadayala, P.; Av-Gay, Y.; Parish, T.; Daffé, M.G.; Stoker, N.G. Inositol monophosphate phosphatase genes of Mycobacterium tuberculosis. Microbiol. Mol. Biol. Rev. 2010, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Rottig, A.; Steinbuchel, A. Acyltransferases in bacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 277–321. [Google Scholar] [CrossRef] [Green Version]
- Barksdale, L.; Kim, K.-S. Mycobacterium. Bacteriol. Rev. 1977, 41, 217. [Google Scholar] [CrossRef]
- Abrahams, K.A.; Besra, G.S. Mycobacterial cell wall biosynthesis: A multifaceted antibiotic target. Parasitology 2018, 145, 116–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, O.B.; Tomasek, D.; Jorge, C.D.; Dufrisne, M.B.; Kim, M.; Banerjee, S.; Rajashankar, K.R.; Shapiro, L.; Hendrickson, W.A.; Santos, H.; et al. Structural basis for phosphatidylinositol-phosphate biosynthesis. Nat. Commun. 2015, 6, 8505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crellin, P.K.; Luo, C.-Y.; Morita, Y.S. Metabolism of Plasma Membrane Lipids in Mycobacteria and Corynebacteria. In Lipid Metabolism; Rodrigo, V.B., Ed.; InTechOpen: Plantation, FL, USA, 2013. [Google Scholar]
- Vilcheze, C.; Hartman, T.; Weinrick, B.; Jacobs, W.R., Jr. Mycobacterium tuberculosis is extraordinarily sensitive to killing by a vitamin C-induced Fenton reaction. Nat. Commun. 2013, 4, 1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salman, M.; Lonsdale, J.T.; Besra, G.S.; Brenna, P.J. Phosphatidylinositol synthesis in mycobacteria. Biochim. Biophys. Acta 1999, 1436, 437–450. [Google Scholar] [CrossRef]
- Ramakrishnan, T.; Murthy, P.S.; Gobinathan, K.P. Intermediary Metabolism of Mycobacteria. Bacteriol. Rev. 1972, 36, 65–108. [Google Scholar] [CrossRef] [PubMed]
- Fulco, A.J.; Bloch, K. Cofactor requirements for the formation of Δ9-unsaturated fatty acids in Mycobacterium phlei. J. Biol. Chem. 1964, 239, 993–997. [Google Scholar] [CrossRef]
- Fulco, A.J.; Levy, R.; Bloch, K. The Biosynthesis of Δ9-and Δ5-Monosaturated Fatty Acids by Bacteria. J. Biol. Chem. 1964, 239, 998–1003. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef]
- Antonova, A.V.; Gryadunov, D.A.; Zimenkov, D.V. Molecular Mechanisms of Drug Tolerance in Mycobacterium tuberculosis. Mol. Biol. 2018, 52, 372–384. [Google Scholar] [CrossRef]
- Wong, D.K.; Lee, B.H.; Horwitz, M.A.; Gibson, B.W. Identification of Fur, Aconitase, and Other Proteins Expressed by Mycobacterium tuberculosis under Conditions of Low and High Concentrations of Iron by Combined Two-Dimensional Gel Electrophoresis and Mass Spectrometry. Infect. Immun. 1999, 67, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bothra, A.; Arumugam, P.; Panchal, V.; Menon, D.; Srivastava, S.; Shankaran, D.; Nandy, A.; Jaisinghani, N.; Singh, A.; Gokhale, R.S.; et al. Phospholipid homeostasis, membrane tenacity and survival of Mtb in lipid rich conditions is determined by MmpL11 function. Sci. Rep. 2018, 8, 8317. [Google Scholar] [CrossRef]
- Wakamoto, Y.; Dhar, N.; Chait, R.; Schneider, K.; Signorino-Gelo, F.; Leibler, S.; McKinney, J.D. Dynamic persistence of antibiotic-stressed mycobacteria. Science 2013, 339, 91–95. [Google Scholar] [CrossRef]
- Gago, G.; Diacovich, L.; Gramajo, H. Lipid metabolism and its implication in mycobacteria-host interaction. Curr. Opin. Microbiol. 2018, 41, 36–42. [Google Scholar] [CrossRef]
- Kumar, R.; Sanyal, S. Mycobacterium tuberculosis: Dormancy, Persistence and Survival in the Light of Protein Synthesis. In Understanding Tuberculosis—Deciphering the Secret Life of the Bacilli; Cardona, P.J., Ed.; IntechOpen: Uppsala, Sweden, 2012. [Google Scholar]
- Gomez, J.E.; McKinney, J.D. M. tuberculosis persistence, latency, and drug tolerance. Tuberculosis 2004, 84, 29–44. [Google Scholar] [CrossRef]
- Dhar, N.; McKinney, J.D. Mycobacterium tuberculosis persistence mutants identified by screening in isoniazid-treated mice. Proc. Natl. Acad. Sci. USA 2010, 107, 12275–12280. [Google Scholar] [CrossRef] [Green Version]
- Horecker, B.L. The pentose phosphate pathway. J. Biol. Chem. 2002, 277, 47965–47971. [Google Scholar] [CrossRef] [Green Version]
- Boshoff, H.I.; Xu, X.; Tahlan, K.; Dowd, C.S.; Pethe, K.; Camacho, L.R.; Park, T.-H.; Yun, C.-S.; Schnappinger, D.; Ehrt, S. Biosynthesis and recycling of nicotinamide cofactors in Mycobacterium tuberculosis: An essential role for NAD in nonreplicating bacilli. J. Biol. Chem. 2008, 283, 19329–19341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wayne, L. Dormancy of Mycobacterium tuberculosis and latency of disease. Eur. J. Clin. Microbiol. Infect. Dis. 1994, 13, 908–914. [Google Scholar] [CrossRef]
- Rhee, K.Y.; de Carvalho, L.P.; Bryk, R.; Ehrt, S.; Marrero, J.; Park, S.W.; Schnappinger, D.; Venugopal, A.; Nathan, C. Central carbon metabolism in Mycobacterium tuberculosis: An unexpected frontier. Trends Microbiol. 2011, 19, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, C.M.; Collier, J.L.; Berg, G.M.; Glibert, P.M. Role of urea in microbial metabolism in aquatic systems: A biochemical and molecular review. Aquat. Microb. Ecol. 2010, 59, 67–88. [Google Scholar] [CrossRef]
- Majumdar, R.; Barchi, B.; Turlapati, S.A.; Gagne, M.; Minocha, R.; Long, S.; Minocha, S.C. Glutamate, Ornithine, Arginine, Proline, and Polyamine Metabolic Interactions: The Pathway Is Regulated at the Post-Transcriptional Level. Front. Plant. Sci. 2016, 7, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.J.; Lim, J.; Gao, S.; Lawson, C.P.; Odell, M.; Raheem, S.; Woo, J.; Kang, S.H.; Kang, S.S.; Jeon, B.Y.; et al. Glutamate mediated metabolic neutralization mitigates propionate toxicity in intracellular Mycobacterium tuberculosis. Sci. Rep. 2018, 8, 8506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yelamanchi, S.D.; Surolia, A. Targeting amino acid metabolism of Mycobacterium tuberculosis for developing inhibitors to curtail its survival. IUBMB Life 2021, 73, 643–658. [Google Scholar] [CrossRef]
- Quartararo, C.E.; Hazra, S.; Hadi, T.; Blanchard, J.S. Structural, kinetic and chemical mechanism of isocitrate dehydrogenase-1 from Mycobacterium tuberculosis. Biochemistry 2013, 52, 1765–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, J.J.; Dechow, S.J.; Abramovitch, R.B. Acid Fasting: Modulation of Mycobacterium tuberculosis Metabolism at Acidic pH. Trends Microbiol. 2019, 27, 942–953. [Google Scholar] [CrossRef]
- Hu, Y.M.; Butcher, P.D.; Sole, K.; Mitchison, D.; Coates, A.R. Protein synthesis is shutdown in dormant Mycobacterium tuberculosis and is reversed by oxygen or heat shock. FEMS Microbiol. Lett. 1998, 158, 139–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopinath, V.; Raghunandanan, S.; Gomez, R.L.; Jose, L.; Surendran, A.; Ramachandran, R.; Pushparajan, A.R.; Mundayoor, S.; Jaleel, A.; Kumar, R.A. Profiling the proteome of Mycobacterium tuberculosis during dormancy and reactivation. Mol. Cell. Proteomics 2015, 14, 2160–2176. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, K.; Yuan, J.; Hohn, M.J.; Jester, B.; Devine, K.M.; Soll, D. From one amino acid to another: tRNA-dependent amino acid biosynthesis. Nucleic Acids Res. 2008, 36, 1813–1825. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.D.; Aínsa, J.A.; Riccardi, G. Role of mycobacterial efflux transporters in drug resistance: An unresolved question. FEMS Microbiol. Rev. 2006, 30, 36–52. [Google Scholar] [CrossRef]
- Yancey, P.H. Water Stress, Osmolytes and Proteins. Am. Zool. 2001, 41, 699–709. [Google Scholar] [CrossRef]
- Vogels, G.D.; Van der Drift, C. Degradation of Purines and Pyrimidines by Microorganisms. Microbiol. Mol. Biol. Rev. 1976, 40, 403–468. [Google Scholar] [CrossRef]
- Villela, A.D.; Sanchez-Quitian, Z.A.; Ducati, R.G.; Santos, D.S.; Basso, L.A. Pyrimidine salvage pathway in Mycobacterium tuberculosis. Curr. Med. Chem. 2011, 18, 1286–1298. [Google Scholar] [CrossRef] [PubMed]
- Knoll, K.E.; Lindeque, Z.; Adenijia, A.A.; Oosthuizen, C.B.; Lall, N.; Loots, D.T. Elucidating the antimycobacterial mechanism of action of ciprofloxacin (accepted). Microorganisms 2021. [Google Scholar] [CrossRef]
- Rizvi, A.; Yousf, S.; Balakrishnan, K.; Dubey, H.K.; Mande, S.C.; Chugh, J.; Banerjee, S. Metabolomics Studies to Decipher Stress Responses in Mycobacterium smegmatis Point to a Putative Pathway of Methylated Amine Biosynthesis. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- Hasenoehrl, E.J.; Rae Sajorda, D.; Berney-Meyer, L.; Johnson, S.; Tufariello, J.M.; Fuhrer, T.; Cook, G.M.; Jacobs, W.R., Jr.; Berney, M. Derailing the aspartate pathway of Mycobacterium tuberculosis to eradicate persistent infection. Nat. Commun. 2019, 10, 4215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudo, F.; Miyanaga, A.; Eguchi, T. Biosynthesis of natural products containing beta-amino acids. Nat. Prod. Rep. 2014, 31, 1056–1073. [Google Scholar] [CrossRef] [Green Version]
- Surger, M.J.; Angelov, A.; Stier, P.; Ubelacker, M.; Liebl, W. Impact of Branched-Chain Amino Acid Catabolism on Fatty Acid and Alkene Biosynthesis in Micrococcus luteus. Front. Microbiol. 2018, 9, 374. [Google Scholar] [CrossRef]
- Serafini, A.; Tan, L.; Horswell, S.; Howell, S.; Greenwood, D.J.; Hunt, D.M.; Phan, M.D.; Schembri, M.; Monteleone, M.; Montague, C.R.; et al. Mycobacterium tuberculosis requires glyoxylate shunt and reverse methylcitrate cycle for lactate and pyruvate metabolism. Mol. Microbiol. 2019, 112, 1284–1307. [Google Scholar] [CrossRef] [Green Version]
- Cassilly, C.D.; Reynolds, T.B. PS, It’s Complicated: The Roles of Phosphatidylserine and Phosphatidylethanolamine in the Pathogenesis of Candida albicans and Other Microbial Pathogens. J. Fungi 2018, 4, 28. [Google Scholar] [CrossRef] [Green Version]
- Rachman, H.; Kim, N.; Ulrichs, T.; Baumann, S.; Pradl, L.; Nasser Eddine, A.; Bild, M.; Rother, M.; Kuban, R.J.; Lee, J.S.; et al. Critical role of methylglyoxal and AGE in mycobacteria-induced macrophage apoptosis and activation. PLoS ONE 2006, 1, e29. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Zeng, J.; Wang, X.; Drlica, K.; Zhao, X. Post-stress bacterial cell death mediated by reactive oxygen species. Proc. Natl. Acad. Sci. USA 2019, 116, 10064–10071. [Google Scholar] [CrossRef] [Green Version]
- Iacobino, A.; Piccaro, G.; Pardini, M.; Fattorini, L.; Giannoni, F. Moxifloxacin Activates the SOS Response in Mycobacterium tuberculosis in a Dose- and Time-Dependent Manner. Microorganisms 2021, 9, 255. [Google Scholar] [CrossRef] [PubMed]
- Mueller, E.A.; Levin, P.A. Bacterial Cell Wall Quality Control during Environmental Stress. Mol. Biol. Physiol. 2020, 11, e02456-20. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spetometry: The combination of targeted and untargeted profiling. Curr. Protoc. Mol. Biol. 2017, 114, 1–32. [Google Scholar]
- Franzblau, S.G.; Witzig, R.S.; McLaughlin, J.C.; Torres, P.; Madico, G.; Hernandez, A.; Degnan, M.T.; Cook, M.B.; Quenzer, V.K.; Ferguson, R.M. Rapid, low-technology MIC determination with clinical Mycobacterium tuberculosis isolates by using the microplate Alamar Blue assay. J. Clin. Microbiol. 1998, 36, 362–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Breda, S.V.; Buys, A.; Apostolides, Z.; Nardell, E.A.; Stoltz, A.C. The antimicrobial effect of colistin methanesulfonate on Mycobacterium tuberculosis in vitro. Tuberculosis 2015, 95, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomino, J.; Portaels, F. Effects of decontamination methods and culture conditions on viability of Mycobacterium ulcerans in the BACTEC system. J. Clin. Microbiol. 1998, 36, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Beukes, D.; du Preez, I.; Loots, D.T. Total Metabolome Extraction from Mycobacterial Cells for GC-MS Metabolomics Analysis. Methods Mol. Biol. 2019, 1859, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Gromski, P.S.; Xu, Y.; Kotze, H.L.; Correa, E.; Ellis, D.I.; Armitage, E.G.; Turner, M.L.; Goodacre, R. Influence of missing values substitutes on multivariate analysis of metabolomics data. Metabolites 2014, 4, 433–452. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Wang, J.; Su, M.; Jia, E.; Chen, S.; Chen, T.; Ni, Y. Missing value imputation approach for mass spectrometry-based metabolomics data. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Zhu, Z.-J. MetFlow: An interactive and integrated workflow for metabolomics data cleaning and differential metabolite discovery. Bioinformatics 2019, 35, 2870–2872. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef] [PubMed]
- Meissner-Roloff, R.J.; Koekemoer, G.; Warren, R.M. A metabolomics investigation of a hyper-and hypo-virulent phenotype of Beijing lineage M. tuberculosis. Metabolomics 2012, 8, 1194–1203. [Google Scholar] [CrossRef]
- Caño-Muñiz, S.; Anthony, R.; Niemann, S.; Alffenaa, J.C. New Approaches and Therapeutic Options for Mycobacterium tuberculosis in a Dormant State. Clin. Microbiol. Rev. 2018, 31, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Metabolite Name (CHEBI ID) | Average Concentration (mg/g Cells) (Standard Deviation) | t-Test (p-Value) | Effect Size (d-Value) | PLS-DA (VIP) | Fold Change (Log2) | |
|---|---|---|---|---|---|---|
| Mtb with DQ derivative RMB041 | Mtb controls | |||||
| 9-Octadecenoic acid (36021) | 0.967 (0.192) | 0.560 (0.041) | 0.000 | 34.47 | 2.59 | 0.73 |
| Myo-Inositol (17268) | 0.024 (0.032) | 0.020 (0.002) | 0.000 | 5.298 | 1.99 | 0.20 |
| Ribitol (15963) | 0.318 (0.513) | 0.100 (0.005) | 0.006 | 2.098 | 1.98 | 2.18 |
| Arabitol (18403) | 0.877 (0.962) | 0.790 (0.199) | 0.010 | 2.206 | 1.98 | 0.11 |
| Eicosanoic acid (28822) | 0.018 (0.025) | 0.004 (0.001) | 0.000 | 3.968 | 1.94 | 3.50 |
| Erythritol (17113) | 0.023 (0.027) | 0.028 (0.001) | 0.001 | 3.176 | 1.92 | −0.18 |
| N-Acetyl-Lysine (35704) | 0.000 (0.000) | 0.004 (0.003) | 0.009 | 1.938 | 1.81 | −1.00 |
| 9,12-Octadecadienoic acid (17351) | 0.578 (0.827) | 0.111 (0.008) | 0.016 | 1.835 | 1.67 | 4.21 |
| Valine (16414) | 0.039 (0.063) | 0.022 (0.005) | 0.019 | 1.908 | 1.65 | 0.77 |
| Aspartic acid (17053) | 0.018 (0.033) | 0.013 (0.004) | 0.038 | 1.341 | 1.65 | 0.38 |
| Pentadecanoic acid (42504) | 0.017 (0.023) | 0.004 (0.001) | 0.021 | 1.648 | 1.64 | 3.25 |
| Galactose (28260) | 0.004 (0.002) | 0.011 (0.001) | 0.185 | 0.747 | 1.57 | −0.64 |
| Glutamic acid (16015) | 0.024 (0.043) | 0.015 (0.006) | 0.032 | 1.549 | 1.56 | 0.60 |
| Lysine (18019) | 0.016 (0.030) | 0.009 (0.006) | 0.047 | 1.291 | 1.56 | 0.78 |
| 2-Hydroxyglutaric acid (32796) | 0.003 (0.004) | 0.002 (0.001) | 0.037 | 1.384 | 1.53 | 0.50 |
| Proline (26271) | 0.022 (0.041) | 0.016 (0.013) | 0.060 | 1.202 | 1.50 | 0.38 |
| Glycerol-3-phosphate (15978) | 0.012 (0.016) | 0.007 (0.002) | 0.049 | 1.308 | 1.47 | 0.71 |
| Malonic acid (30794) | 0.173 (0.213) | 0.039 (0.045) | 0.249 | 0.684 | 1.46 | 3.44 |
| Glycerol (17754) | 0.994 (1.45) | 0.633 (0.031) | 0.057 | 1.171 | 1.44 | 0.57 |
| Nonadecanoic acid (NSC11914) | 0.499 (0.731) | 0.055 (0.009) | 0.065 | 1.127 | 1.41 | 8.07 |
| Alanine (16977) | 0.321 (0.621) | 0.207 (0.152) | 0.067 | 1.156 | 1.40 | 0.55 |
| Octadecanoic acid (28842) | 0.027 (0.034) | 0.005 (0.000) | 0.068 | 1.150 | 1.40 | 4.40 |
| β-Aminoisobutanoic acid (33094) | 0.035 (0.057) | 0.014 (0.004) | 0.073 | 1.310 | 1.38 | 1.50 |
| γ-Aminobutyric acid (16865) | 0.011 (0.018) | 0.016 (0.006) | 0.085 | 1.315 | 1.34 | −0.31 |
| Tyrosine (17895) | 0.006 (0.012) | 0.003 (0.001) | 0.147 | 0.843 | 1.32 | 1.00 |
| 9-Hexadecenoic acid (59265) | 0.105 (0.153) | 0.018 (0.003) | 0.099 | 0.975 | 1.29 | 4.83 |
| Heptadecanoic acid (32365) | 0.052 (0.069) | 0.011 (0.002) | 0.103 | 0.975 | 1.29 | 3.73 |
| Ribofuranose (33942) | 0.003 (0.003) | 0.007 (0.002) | 0.239 | 0.653 | 1.28 | −0.57 |
| Glucose (17243) | 0.045 (0.031) | 0.037 (0.005) | 0.174 | 0.770 | 1.24 | 0.22 |
| Isoleucine (17191) | 0.018 (0.032) | 0.008 (0.008) | 0.216 | 0.702 | 1.21 | 1.25 |
| 9Z-Octadec-1-ol (73504) | 0.026 (0.036) | 0.004 (0.003) | 0.276 | 0.601 | 1.17 | 5.50 |
| Decanoic acid (30813) | 0.008 (0.009) | 0.005 (0.001) | 0.157 | 0.836 | 1.18 | 0.60 |
| Malic acid (6650) | 0.084 (0.127) | 0.054 (0.004) | 0.172 | 0.775 | 1.11 | 0.56 |
| Ornithine (15729) | 0.029 (0.054) | 0.025 (0.015) | 0.191 | 0.841 | 1.09 | 0.16 |
| Serine (17822) | 0.011 (0.021) | 0.004 (0.002) | 0.151 | 0.850 | 1.06 | 1.75 |
| Threonine (16857) | 0.049 (0.088) | 0.016 (0.008) | 0.173 | 0.791 | 0.93 | 2.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knoll, K.E.; Lindeque, Z.; Adeniji, A.A.; Oosthuizen, C.B.; Lall, N.; Loots, D.T. Elucidating the Antimycobacterial Mechanism of Action of Decoquinate Derivative RMB041 Using Metabolomics. Antibiotics 2021, 10, 693. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060693
Knoll KE, Lindeque Z, Adeniji AA, Oosthuizen CB, Lall N, Loots DT. Elucidating the Antimycobacterial Mechanism of Action of Decoquinate Derivative RMB041 Using Metabolomics. Antibiotics. 2021; 10(6):693. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060693
Chicago/Turabian StyleKnoll, Kirsten E., Zander Lindeque, Adetomiwa A. Adeniji, Carel B. Oosthuizen, Namrita Lall, and Du Toit Loots. 2021. "Elucidating the Antimycobacterial Mechanism of Action of Decoquinate Derivative RMB041 Using Metabolomics" Antibiotics 10, no. 6: 693. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060693