
Rapid and Accurate Detection of Escherichia coli and Klebsiella pneumoniae Strains Susceptible/Resistant to Cotrimoxazole through Evaluation of Cell Elongation


Abstract
:1. Introduction
2. Results
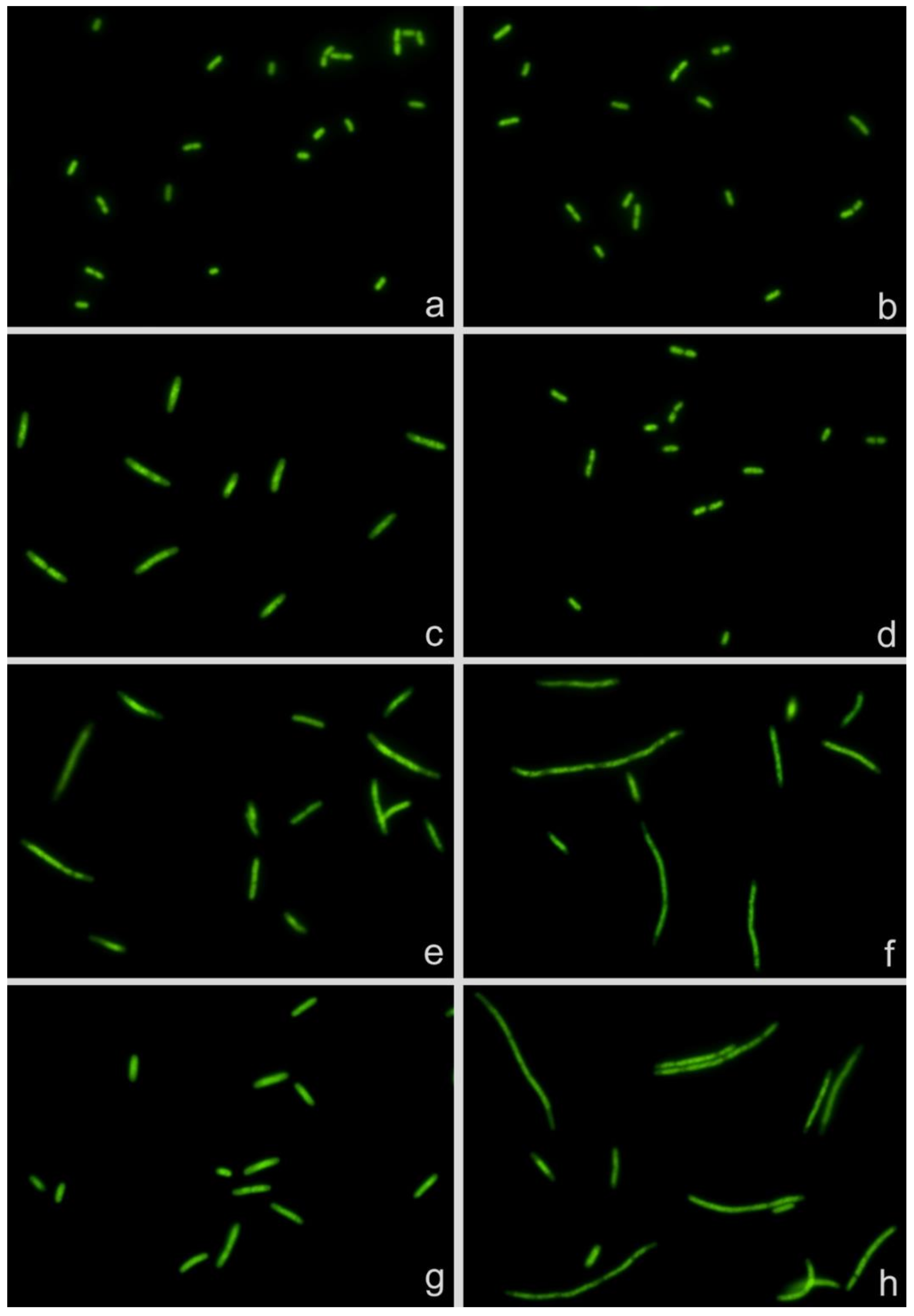
2.1. Technical Assay
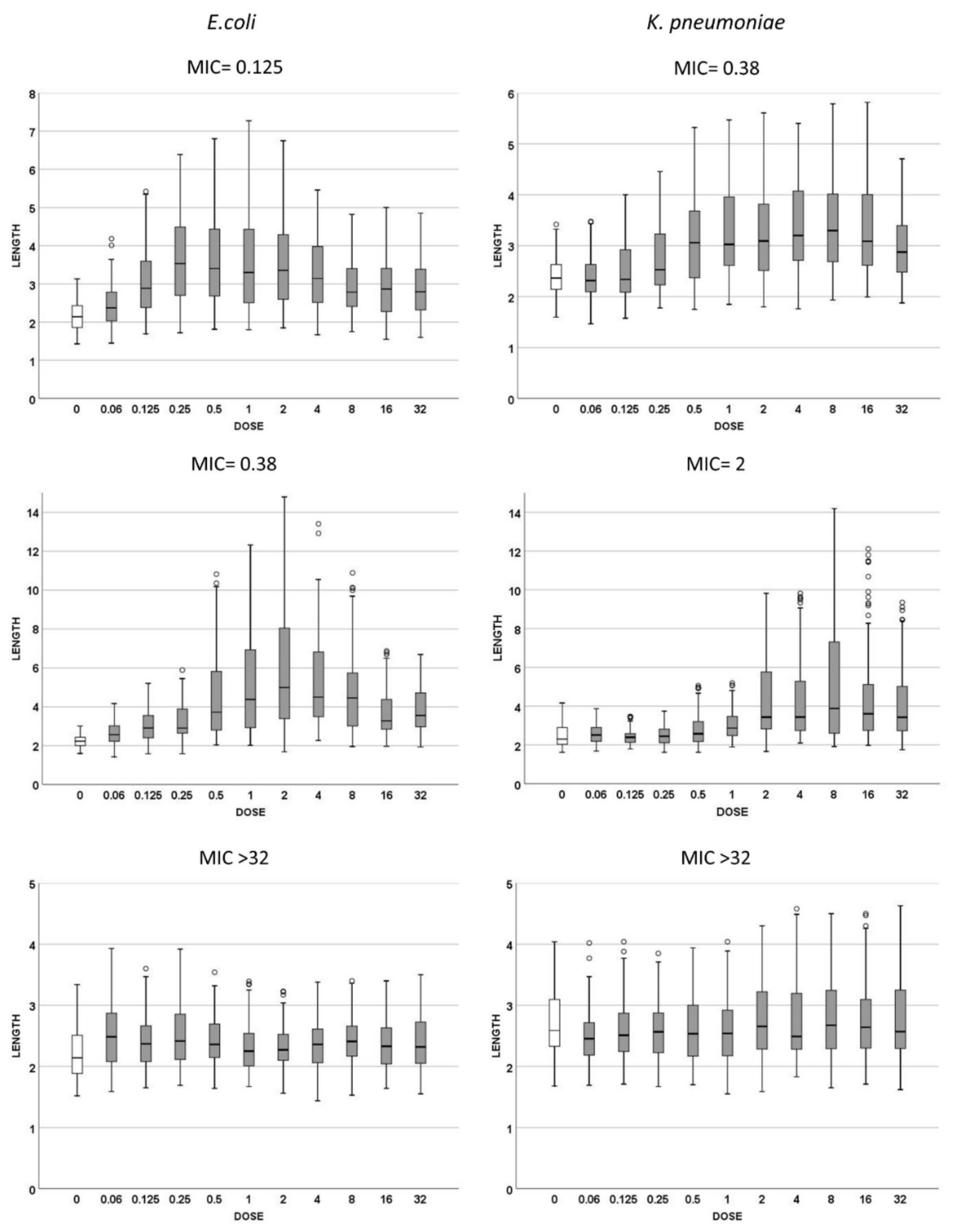
2.2. Dose Response
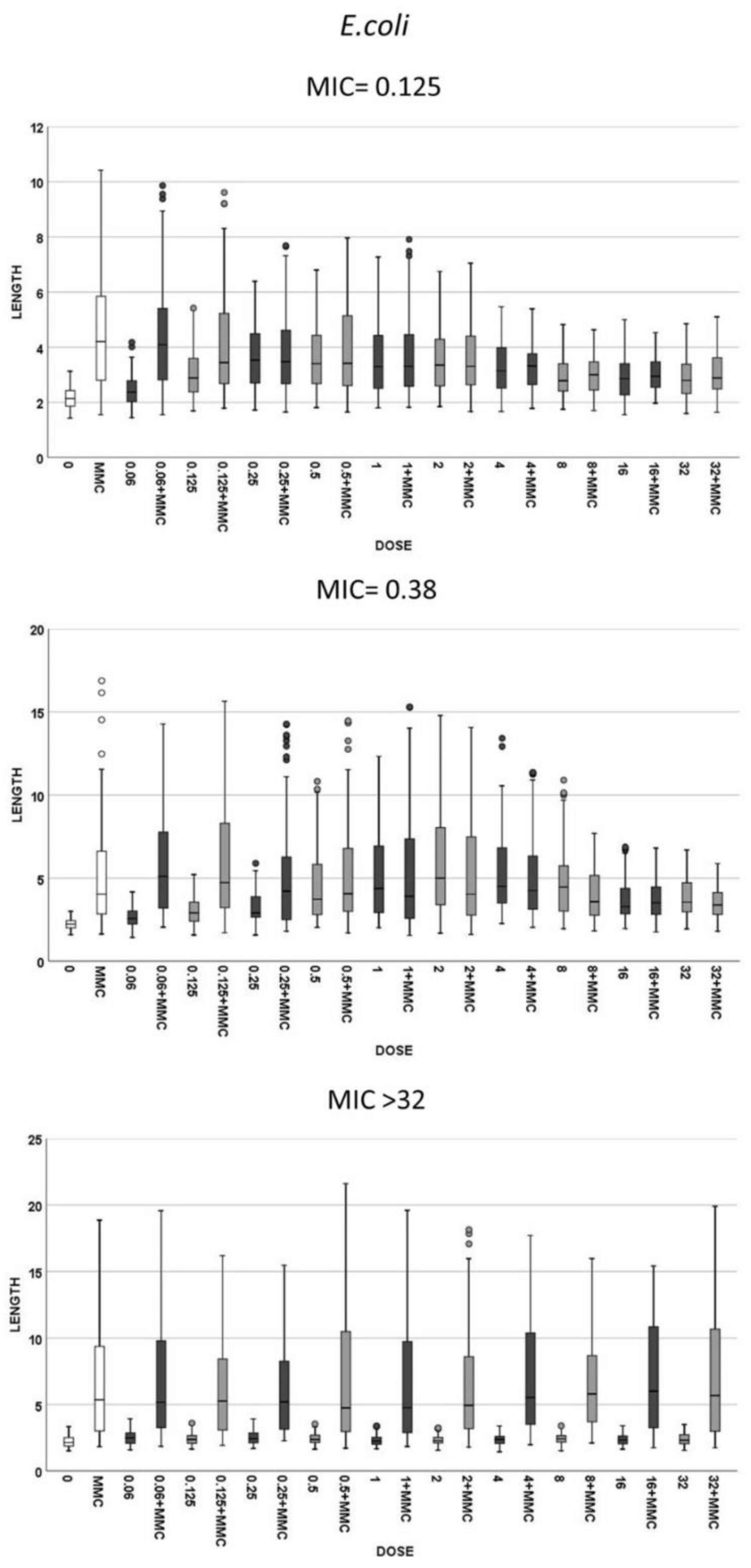
2.3. Assay Validation
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Minimum Inhibitory Concentrations (MICs) Determination
4.2. Determining Susceptibility-Resistance Using Cell Elongation
4.3. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bushby, S.R.M.; Hitchings, G.H. Thrimethoprim, a sulphonamide potentiator. Br. J. Pharmac. Chemother 1968, 33, 72–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darrell, J.H.; Garrod, L.P.; Waterworth, P.M. Trimethoprim: Laboratory and clinical studies. J. Clin. Pathol. 1968, 21, 202–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masters, P.A.; O’Bryan, T.A.; Zurlo, J.; Miller, D.Q.; Joshi, N. Trimethoprim-sulfamethoxazole revisited. Arch. Intern. Med. 2003, 163, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Smilack, J.D. Trimethoprim-sulfamethoxazole. Mayo Clin. Proc. 1999, 74, 730–744. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Bátori, Z.; Ábrók, M.; Lázár, A.; Burián, K. Characterization of Resistance in Gram-Negative Urinary Isolates Using Existing and Novel Indicators of Clinical Relevance: A 10-Year Data Analysis. Life 2020, 10, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooke, J.S. Stenotrophomonas maltophilia: An Emerging Global Opportunistic Pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Grammatikos, A.P.; Michalopoulos, A. Potential of old-generation antibiotics to address current need for new antibiotics. Expert Rev. Anti-Infect. Ther. 2008, 6, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Cassir, N.; Rolain, J.M.; Brouqui, P. A new strategy to fight antimicrobial resistance: The revival of old antibiotics. Front. Microbiol. 2014, 5, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaye, K.S.; Gales, A.C.; Dubourg, G. Old antibiotics for multidrug-resistant pathogens: From in vitro activity to clinical outcomes. Int. J. Antimicrob. Agents 2017, 49, 542–548. [Google Scholar] [CrossRef]
- Huovinen, P. Resistance to trimethoprim–sulfamethoxazole. Clin. Infect. Dis. 2001, 32, 1608–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingham, C.J.; van den Ende, M.; Wever, P.C.; Schneeberger, P.M. Rapid antibiotic sensitivity testing and trimethoprim-mediated filamentation of clinical isolates of the Enterobacteriaceae assayed on a novel porous culture support. J. Med. Microbiol. 2006, 55, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Otero, F.; Gosálvez, J.; Bou, G.; Fernández, J.L. Simple and fast detection of resistance to antibiotic inhibitors of protein synthesis in Gram-negative pathogens through evaluation of mitomycin C-induced cell elongation. Microb. Drug Resist. 2017, 23, 973–981. [Google Scholar] [CrossRef]
- Suzuki, H.; Kilgore, W.W. Effects of mitomycin C on macromolecular synthesis in Escherichia coli. J. Bacteriol. 1967, 93, 675–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisman, O.; D’Ari, R.; Gottesman, S. Cell-division control in Escherichia coli: Specific induction of the SOS function SfiA protein is sufficient to block septation. Proc. Natl. Acad. Sci. USA 1984, 81, 4490–4494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasz, M.; Lipman, R.; Chowdary, D. Isolation and structure of a covalent cross-link adduct between mitomycin C and DNA. Science 1987, 235, 1204–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baharoglu, Z.; Mazel, D. SOS, the formidable strategy of bacteria against aggressions. FEMS Microbiol. 2014, 38, 1126–1145. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Milan, S.L.; Erickson, H.P. SulA inhibits assembly of FtsZ by a simple sequestration mechanism. Biochemistry 2012, 51, 3100–3109. [Google Scholar] [CrossRef]
- Margolin, W. FtsZ and the division of prokaryotic cells and organelles. Nat. Rev. Mol. Cell. Biol. 2005, 6, 862–871. [Google Scholar] [CrossRef] [Green Version]
- Gunderson, C.W.; Segall, A.M. DNA repair, a novel antibacterial target: Holliday junction-trapping peptides induce DNA damage and chromosome segregation defects. Mol. Microbiol. 2006, 59, 1129–1148. [Google Scholar] [CrossRef]
- Boberek, J.M.; Stach, J.; Good, L. Genetic evidence for inhibition of bacterial division protein FtsZ by berberine. PLoS ONE 2010, 10, e13745. [Google Scholar] [CrossRef]
- Singh, S.B.; Young, K.; Silver, L.L. What is an “ideal” antibiotic? Discovery challenges and path forward. Biochem Pharmacol. 2017, 133, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Lewin, C.S.; Amyes, S.G. The role of the SOS response in bacteria exposed to zidovudine or trimethoprim. J. Med. Microbiol. 1991, 34, 329–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. CLSI Document M100, 30th ed.; CLSI: Wayne, PA, USA, 2020. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | CLSI Classification | Technical Assay | Sensitivity (%), 95% CI | Specificity (%), 95% CI | AUC, 95% CI, p < 0.0001 | |
|---|---|---|---|---|---|---|
| S | NS | |||||
| E. coli | 23 | 27 | CTX | 92.59 (80.86–100) | 95.65 (85.14–100) | 0.941 (0.866–1.00) |
| CTX+MMC | 100 (98.15–100) | 100 (97.83–100) | 1 (1.00–1.00) | |||
| K. pneumoniae | 34 | 18 | CTX | 83.33 (63.34–100) | 91.18 (80.17–100) | 0.873 (0.757–0.988) |
| CTX+MMC | 100 (97.22–100) | 100 (98.53–100) | 1 (1.00–1.00) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López, I.; Otero, F.; Guillén, R.; Fernández, M.d.C.; Bou, G.; Gosálvez, J.; Fernández, J.L. Rapid and Accurate Detection of Escherichia coli and Klebsiella pneumoniae Strains Susceptible/Resistant to Cotrimoxazole through Evaluation of Cell Elongation. Antibiotics 2021, 10, 720. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060720
López I, Otero F, Guillén R, Fernández MdC, Bou G, Gosálvez J, Fernández JL. Rapid and Accurate Detection of Escherichia coli and Klebsiella pneumoniae Strains Susceptible/Resistant to Cotrimoxazole through Evaluation of Cell Elongation. Antibiotics. 2021; 10(6):720. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060720
Chicago/Turabian StyleLópez, Isidoro, Fátima Otero, Rebeca Guillén, María del Carmen Fernández, Germán Bou, Jaime Gosálvez, and José Luis Fernández. 2021. "Rapid and Accurate Detection of Escherichia coli and Klebsiella pneumoniae Strains Susceptible/Resistant to Cotrimoxazole through Evaluation of Cell Elongation" Antibiotics 10, no. 6: 720. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060720