
Bioguided Isolation of Antibiofilm and Antibacterial Pregnane Glycosides from Caralluma quadrangula: Disarming Multidrug-Resistant Pathogens



Abstract
:1. Introduction
2. Results
2.1. Minimum Bactericidal Concentration (MBC) and Antibiofilm Assays for C. quadrangula Extracts
2.2. In Vivo Evaluation of C. quadrangula Extracts against MRSA USA300 Skin Infection
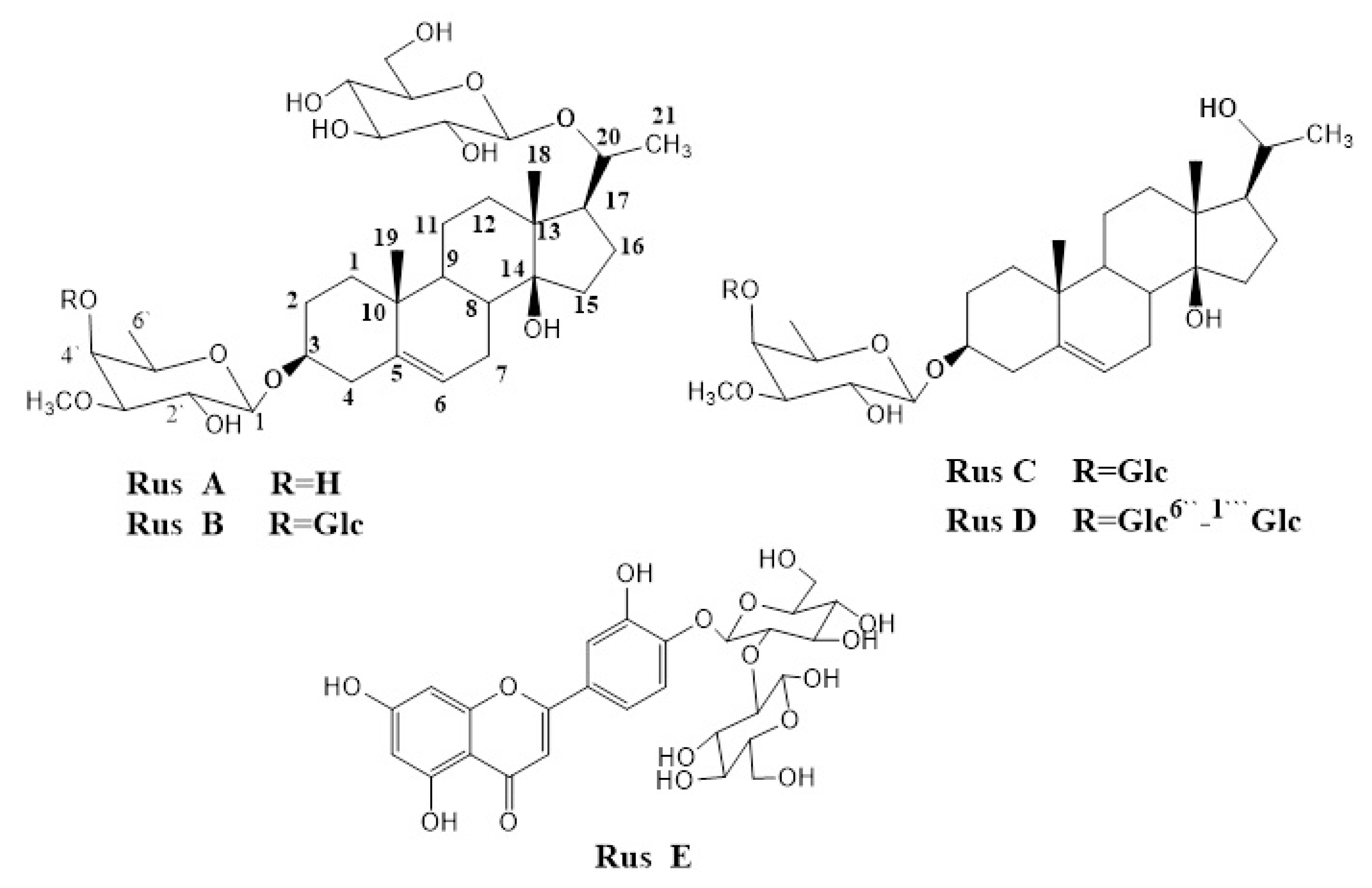
2.3. Extraction and Isolation of the Major Glycosides from N-Butanol Fraction
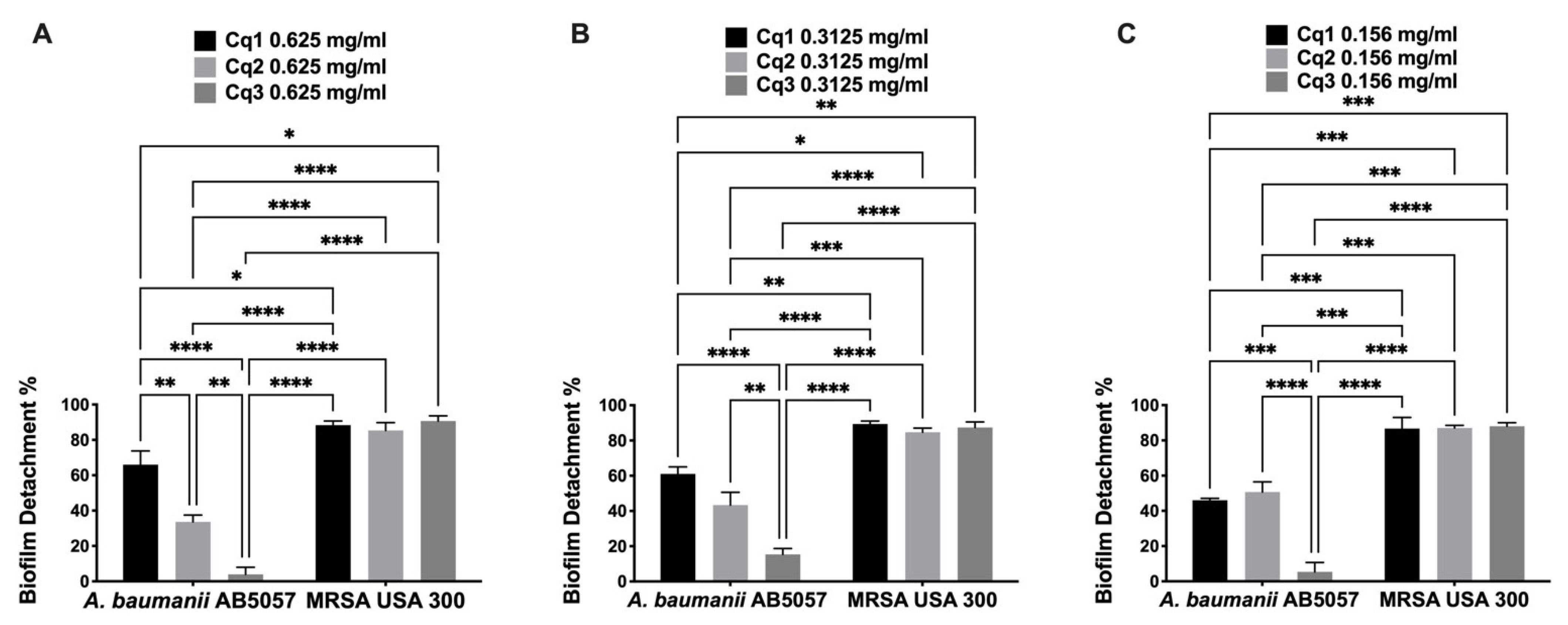
2.4. MBC and Antibiofilm Assays of the Isolated Compounds
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. Extraction and Isolation of Major Compounds
4.4. Biological Assays
4.4.1. In Vitro Studies
Bacterial Strains and Growth Conditions
Minimum Bactericidal Concentration (MBC)
Antibiofilm Screening Assay
Biofilm Detachment Activity
4.4.2. In Vivo Study
Animals
In Vivo Evaluation of C. quadrangula Extracts against MRSA Skin Infection in Mice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between Antibiotic Resistance, Biofilm Formation, and Biofilm-Specific Resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genovese, C.; D’Angeli, F.; Bellia, F.; Distefano, A.; Spampinato, M.; Attanasio, F.; Nicolosi, D.; Di Salvatore, V.; Tempera, G.; Lo Furno, D.; et al. In Vitro Antibacterial, Anti-Adhesive and Anti-Biofilm Activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson Root Extract against Methicillin-Resistant Staphylococcus aureus Strains. Antibiotics 2021, 10, 428. [Google Scholar] [CrossRef]
- Liu, H.; Wu, Y.-Q.; Chen, L.-P.; Gao, X.; Huang, H.-N.; Qiu, F.-L.; Wu, D.-C. Biofilm-Related Genes: Analyses in Multi-Antibiotic Resistant Acinetobacter baumannii isolates from Mainland China. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 1801–1807. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Xie, H.; Zhang, B.; Zhou, J.; Dou, Z.; Wang, X.; Wang, N. A Novel Ivermectin-Derived Compound D4 and Its Antimicrobial/Biofilm Properties against MRSA. Antibiotics 2021, 10, 208. [Google Scholar] [CrossRef]
- Bagińska, N.; Cieślik, M.; Górski, A.; Jończyk-Matysiak, E. The Role of Antibiotic Resistant A. baumannii in the Pathogenesis of Urinary Tract Infection and the Potential of Its Treatment with the Use of Bacteriophage Therapy. Antibiotics 2021, 10, 281. [Google Scholar] [CrossRef]
- Delancey, E.; Allison, D.; Kc, H.R.; Gilmore, D.F.; Fite, T.; Basnakian, A.G.; Alam, M.A. Synthesis of 4,4′-(4-Formyl-1H-pyrazole-1,3-diyl)dibenzoic Acid Derivatives as Narrow Spectrum Antibiotics for the Potential Treatment of Acinetobacter Baumannii Infections. Antibiotics 2020, 9, 650. [Google Scholar] [CrossRef] [PubMed]
- Han, H.-W.; Kwak, J.-H.; Jang, T.-S.; Knowles, J.C.; Kim, H.-W.; Lee, H.-H.; Lee, J.-H. Grapefruit Seed Extract as a Natural Derived Antibacterial Substance against Multidrug-Resistant Bacteria. Antibiotics 2021, 10, 85. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.Q.; Estellés, A.; Li, L.; Abdelhady, W.; Gonzales, R.; Bayer, A.S.; Tenorio, E.; Leighton, A.; Ryser, S.; Kauvar, L.M. A Human Biofilm-Disrupting Monoclonal Antibody Potentiates Antibiotic Efficacy in Rodent Models of both Staphylococcus aureus and Acinetobacter baumannii Infections. Antimicrob. Agents Chemother. 2017, 61, e00904–e00917. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, A.C.; Thompson, M.G.; Black, C.C.; Kessler, J.L.; Clark, L.P.; McQueary, C.N.; Gancz, H.Y.; Corey, B.W.; Moon, J.K.; Si, Y.; et al. AB5075, a Highly Virulent Isolate of Acinetobacter baumannii, as a Model Strain for the Evaluation of Pathogenesis and Antimicrobial Treatments. mBio 2014, 5, e01076. [Google Scholar] [CrossRef] [Green Version]
- Lesho, E.; Yoon, E.J.; McGann, P.; Snesrud, E.; Kwak, Y.; Milillo, M.; Onmus-Leone, F.; Preston, L.; St Clair, K.; Nikolich, M.; et al. Emergence of colistin-resistance in extremely drug-resistant Acinetobacter baumannii containing a novel pmrCAB operon during colistin therapy of wound infections. J. Infect. Dis. 2013, 208, 1142–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.A.; El-Shiekh, R.A.; Hashem, R.A.; Hassan, M. In vivo Antibacterial Activity of Star Anise (Illicium verum Hook.) Extract Using Murine MRSA Skin Infection Model in Relation to Its Metabolite Profile. Infect. Drug Resist. 2021, 14, 33–48. [Google Scholar] [CrossRef]
- Ezzat, M.I.; Hassan, M.; Abdelhalim, M.A.; El-Desoky, A.M.; Mohamed, S.O.; Ezzat, S.M. Immunomodulatory effect of Noni fruit and its isolates: Insights into cell-mediated immune response and inhibition of LPS-induced THP-1 macrophage inflammation. Food Funct. 2021, 12, 3170–3179. [Google Scholar] [CrossRef]
- Elzeini, H.M.; Ali, A.R.A.A.; Nasr, N.F.; Hassan, M.; Hassan, A.A.m.; Elenany, Y.E. Probiotic capability of novel lactic acid bacteria isolated from worker honey bees gut microbiota. FEMS Microbiol. Lett. 2021, 368. [Google Scholar] [CrossRef]
- Bin-Jumah, M.N. Antidiabetic Effect of Monolluma quadrangula Is Mediated via Modulation of Glucose Metabolizing Enzymes, Antioxidant Defenses, and Adiponectin in Type 2 Diabetic Rats. Oxid. Med. Cell. Longev. 2019, 2019, 6290143. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Jan, S.; Mussarat, S.; Tariq, A.; Begum, S.; Afroz, A.; Shinwari, Z.K. A review on ethnobotany, phytochemistry and pharmacology of plant genus Caralluma r. Br. J. Pharm. Pharmacol. 2014, 66, 1351–1368. [Google Scholar] [CrossRef]
- Farouk, A.-E.; Ahamed, N.T.; AlZahrani, O.; Alamer, K.H.; Al-Sodany, Y.; Bahobail, A.A. Antimicrobial Activity of Caralluma quadrangula (Forssk) N.E. Br Latex from Al-Shafa Taif, Kingdom of Saudi Arabia. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 284–298. [Google Scholar] [CrossRef] [Green Version]
- Babu, K.S.; Malladi, S.; Nadh, R.V.; Rambabu, S.S. Evaluation of in Vitro Antibacterial Activity of Caralluma umbellata Haw Used in Traditional Medicine by Indian Tribes. Annu. Res. Rev. Biol. 2013, 4, 840–855. [Google Scholar] [CrossRef]
- Reddy, K.D.; Reddy, K.H.; Rao, G.V.; Brenda, M.; Patrick, G.; Koorbanally, N.A. In vitro Antimicrobial, Antioxidant and Cytotoxic Activities of New Pregnane Glycosides and Pregnanes Isolated from the Carallum adescendens var. gracilis and Caralluma pauciflora. J. Pure Appl. Microbiol. 2013, 7, 2707–2712. [Google Scholar]
- Malladi, S.; Ratnakaram, V.N.; Suresh Babu, K.; Pullaiah, T. Evaluation of in vitro antibacterial activity of Caralluma lasiantha for scientific validation of Indian traditional medicine. Cogent Chem. 2017, 3, 1374821. [Google Scholar] [CrossRef]
- Amrati, F.E.; Bourhia, M.; Saghrouchni, H.; Slighoua, M.; Grafov, A.; Ullah, R.; Ezzeldin, E.; Mostafa, G.A.; Bari, A.; Ibenmoussa, S.; et al. Caralluma europaea (Guss.) N.E.Br.: Anti-Inflammatory, Antifungal, and Antibacterial Activities against Nosocomial Antibiotic-Resistant Microbes of Chemically Characterized Fractions. Molecules 2021, 26, 636. [Google Scholar] [CrossRef]
- Gushash, A. Plants in the Mountains of Sarat and Hejaz; Sarawat Designer and Printers: Madinah, Saudi Arabia, 2006. [Google Scholar]
- Abdel-Sattar, E.; El-Maraghy, S.A.; El-Dine, R.S.; Rizk, S.M. Antihyperglycemic activity of Caralluma quadrangula in streptozotocin-induced diabetic rats. Bull. Fac. Pharm. Cairo Univ. 2017, 55, 269–272. [Google Scholar] [CrossRef]
- Abdel-Sattar, E.; El-Maraghy, S.A.; El-Dine, R.S.; Rizk, S.M. Russelioside B, a pregnane glycoside ameliorates hyperglycemia in streptozotocin induced diabetic rats by regulating key enzymes of glucose metabolism. Chem. Biol. Interact. 2016, 252, 47–53. [Google Scholar] [CrossRef]
- Abdel-Sattar, E.; Mehanna, E.T.; El-Ghaiesh, S.H.; Mohammad, H.M.F.; Elgendy, H.A.; Zaitone, S.A. Pharmacological Action of a Pregnane Glycoside, Russelioside B, in Dietary Obese Rats: Impact on Weight Gain and Energy Expenditure. Front. Pharmacol. 2018, 9, 990. [Google Scholar] [CrossRef]
- El-Shiekh, R.A.; El-Mekkawy, S.; Mouneir, S.M.; Hassan, A.; Abdel-Sattar, E. Therapeutic potential of russelioside B as anti-arthritic agent in Freund’s adjuvant-induced arthritis in rats. J. Ethnopharmacol. 2021, 270, 113779. [Google Scholar] [CrossRef] [PubMed]
- El-Shiekh, R.A.; Salama, A.; Al-Mokaddem, A.K.; Bader, A.; Abdel-Sattar, E.A. Russelioside B, a pregnane glycoside for treatment of gastric ulcer via modulation of heat shock protein-70 and vascular endothelial growth factor. Steroids 2021, 165, 108759. [Google Scholar] [CrossRef]
- Abdul-Aziz Al-Yahya, M.; Abdel-Sattar, E.; Guittet, E. Pregnane glycosides from Caralluma russeliana. J. Nat. Prod. 2000, 63, 1451–1453. [Google Scholar] [CrossRef]
- Govindaraj Vaithinathan, A.; Vanitha, A. WHO global priority pathogens list on antibiotic resistance: An urgent need for action to integrate One Health data. Perspect. Public Health 2018, 138, 87–88. [Google Scholar] [CrossRef]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Belanger, C.R.; Mansour, S.C.; Pletzer, D.; Hancock, R.E.W. Alternative strategies for the study and treatment of clinical bacterial biofilms. Emerg. Top. Life Sci. 2017, 1, 41–53. [Google Scholar] [CrossRef]
- Imperi, F.; Fiscarelli, E.V.; Visaggio, D.; Leoni, L.; Visca, P. Activity and Impact on Resistance Development of Two Antivirulence Fluoropyrimidine Drugs in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2019, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Krysinski, E.P.; Brown, L.J.; Marchisello, T.J. Effect of cleaners and sanitizers on Listeria monocytogenes attached to product contact surfaces. J. Food Prot. 1992, 55, 246–251. [Google Scholar] [CrossRef]
- Sandasi, M.; Leonard, C.M.; Viljoen, A.M. The in vitro antibiofilm activity of selected culinary herbs and medicinal plants against Listeria monocytogenes. Lett. Appl. Microbiol. 2010, 50, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.; Costa, S.S.; Serrano, M.; Oliveira, K.; Trigueiro, G.; Pomba, C.; Couto, I. Clonal Lineages, Antimicrobial Resistance, and PVL Carriage of Staphylococcus aureus Associated to Skin and Soft-Tissue Infections from Ambulatory Patients in Portugal. Antibiotics 2021, 10, 345. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Chiller, K.; Selkin, B.A.; Murakawa, G.J. Skin microflora and bacterial infections of the skin. J. Investig. Dermatol. Symp. Proc. 2001, 6, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Buommino, E.; Vollaro, A.; Nocera, F.P.; Lembo, F.; DellaGreca, M.; De Martino, L.; Catania, M.R. Synergistic Effect of Abietic Acid with Oxacillin against Methicillin-Resistant Staphylococcus pseudintermedius. Antibiotics 2021, 10, 80. [Google Scholar] [CrossRef]
- Komarnytsky, S.; Esposito, D.; Poulev, A.; Raskin, I. Pregnane glycosides interfere with steroidogenic enzymes to down-regulate corticosteroid production in human adrenocortical H295R cells. J. Cell. Physiol. 2013, 228, 1120–1126. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Sattar, E.A.; Abdallah, H.M.; Khedr, A.; Abdel-Naim, A.B.; Shehata, I.A. Antihyperglycemic activity of Caralluma tuberculata in streptozotocin-induced diabetic rats. Food Chem. Toxicol. 2013, 59, 111–117. [Google Scholar] [CrossRef]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; et al. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K.; CLSI Methods Development and Standardization Working Group of the Subcommittee on Antimicrobial Susceptibility Testing. CLSI Methods Development and Standardization Working Group Best Practices for Evaluation of Antimicrobial Susceptibility Tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biofilm Inhibition | Biofilm Detachment | |||||||
|---|---|---|---|---|---|---|---|---|
| LogIC50 ± SE | Mean IC50 | LogIC50 ± SE | Mean IC50 | |||||
| - | (μg/mL) | (µmole) | (mmol) | - | (μg/mL) | (µmole) | (mmol) | |
| Rus A | 2.286 ± 0.083 | 193.2026 | 294.62± | 0.295 | 3.62 ± 0.318 | 4164.101 | 6347.71 | 6.347 |
| Rus B | 2.358 ± 0.098 | 227.9783 | 278.7 | 0.279 | 2.842 ± 0.272 | 694.9902 | 849.62 | 0.849 |
| Rus C | 1.961 ± 0.141 | 91.43766 | 139.39 | 0.139 | 2.919 ± 0.125 | 829.7031 | 1264.79 | 1.264 |
| Rus D | 2.385 ± 0.122 | 242.6153 | 296.6 | 0.296 | 3.166 ± 0.496 | 1465.124 | 1791.1 | 1.791 |
| Rus E | 2.687 ± 0.093 | 486.1934 | 818.51 | 0.818 | 5.142 ± 0.943 | 138,566.6 | 233,277.1 | 233.277 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Shiekh, R.A.; Hassan, M.; Hashem, R.A.; Abdel-Sattar, E. Bioguided Isolation of Antibiofilm and Antibacterial Pregnane Glycosides from Caralluma quadrangula: Disarming Multidrug-Resistant Pathogens. Antibiotics 2021, 10, 811. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070811
El-Shiekh RA, Hassan M, Hashem RA, Abdel-Sattar E. Bioguided Isolation of Antibiofilm and Antibacterial Pregnane Glycosides from Caralluma quadrangula: Disarming Multidrug-Resistant Pathogens. Antibiotics. 2021; 10(7):811. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070811
Chicago/Turabian StyleEl-Shiekh, Riham A., Mariam Hassan, Rasha A. Hashem, and Essam Abdel-Sattar. 2021. "Bioguided Isolation of Antibiofilm and Antibacterial Pregnane Glycosides from Caralluma quadrangula: Disarming Multidrug-Resistant Pathogens" Antibiotics 10, no. 7: 811. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070811