
Analysis of Wastewater Reveals the Spread of Diverse Extended-Spectrum β-Lactamase-Producing E. coli Strains in uMgungundlovu District, South Africa



Abstract
:1. Introduction
2. Results
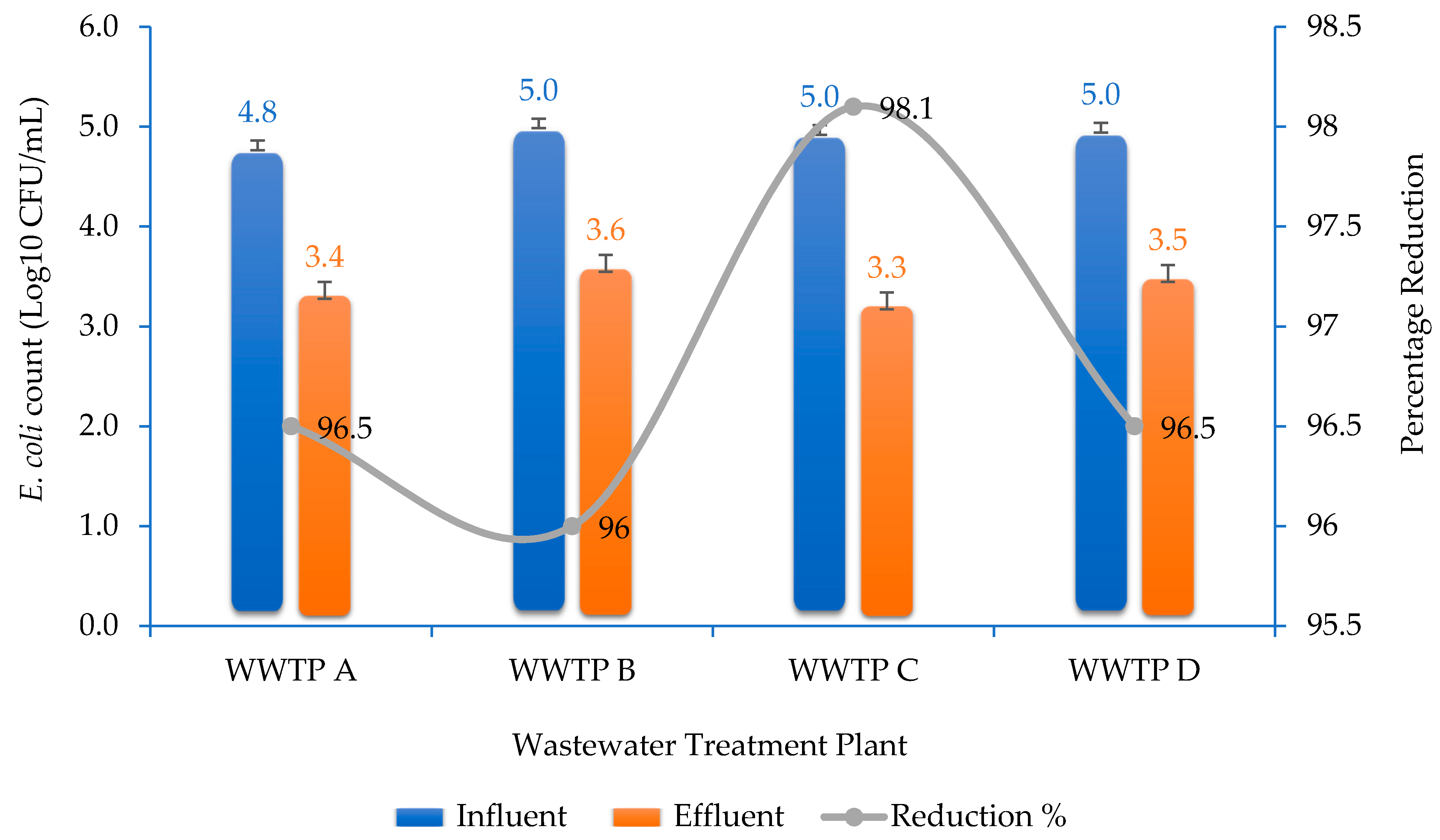
2.1. Enumeration of Isolates
2.2. Antimicrobial Susceptibility
2.3. Prevalence of Multidrug Resistance in E. coli Isolates
2.4. ESBL Resistance Genes
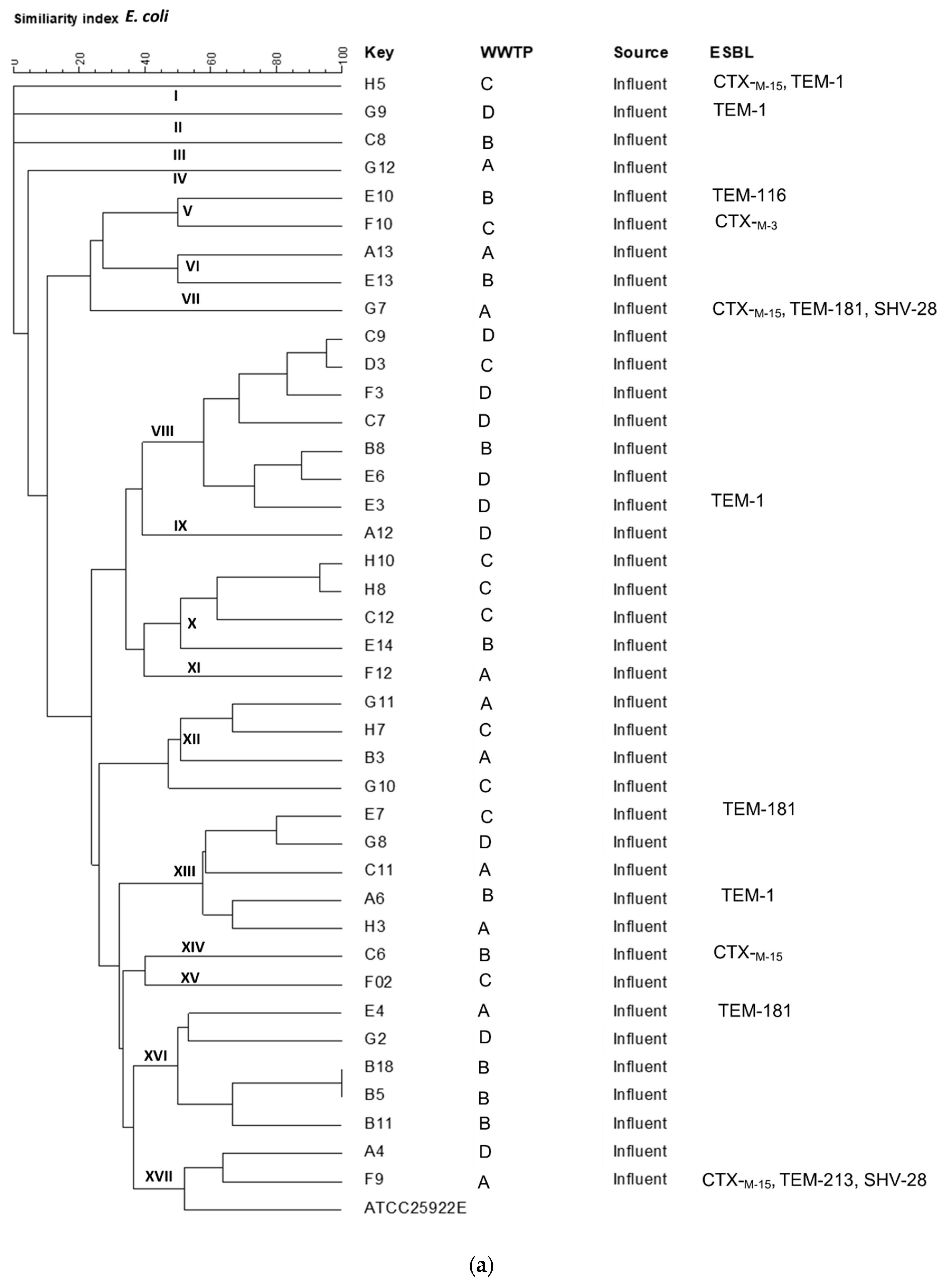
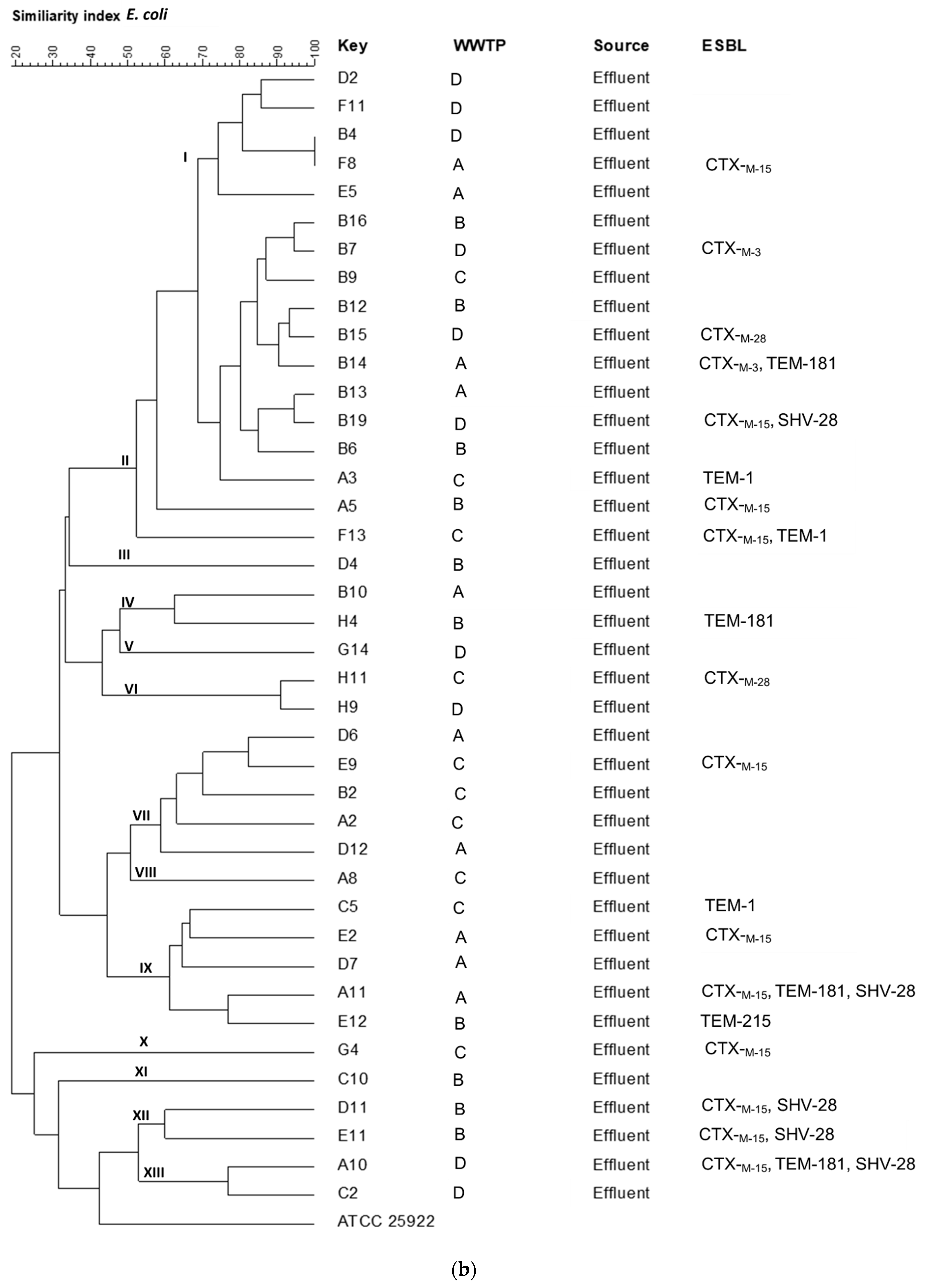
2.5. Strain Typing by ERIC-PCR
3. Discussion
3.1. Enumeration of E. coli
3.2. Antibiotic Susceptibility
3.3. Multidrug Resistance and ESBL Production
3.4. Clonality of Isolates
4. Materials and Methods
4.1. Study Sites
4.2. Sampling
4.3. Quantitative Analysis and Isolate Identification
4.4. Molecular Confirmation of E. coli by Real-Time Polymerase Chain Reaction (PCR)
4.5. Antibiotic Susceptibility Testing and Determination of ESBL Production
4.6. Genotypic Characterisation of ESBLs
4.7. Strain Typing by ERIC-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Priority List of Antibiotic-Resistant Batceria to Guide Research, Discovery, and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Ramadan, A.A.; Abdelaziz, N.A.; Amin, M.A.; Aziz, R.K. Novel bla CTX-M variants and genotype-phenotype correlations among clinical isolates of extended spectrum beta lactamase-producing Escherichia coli. Sci. Rep. 2019, 9, 4224. [Google Scholar] [CrossRef] [PubMed]
- Franz, E.; Veenman, C.; Van Hoek, A.H.A.M.; Husman, A.D.R.; Blaak, H. Pathogenic Escherichia coli producing Extended-Spectrum β-Lactamases isolated from surface water and wastewater. Sci. Rep. 2015, 5, 14372. [Google Scholar] [CrossRef] [Green Version]
- Furlan, J.P.R.; Stehling, E.G. Presence of Β-Lactamases Encoding Genes in Soil Samples from Different Origins. Water Air Soil Pollut. 2017, 228, 11270. [Google Scholar] [CrossRef]
- Adegoke, A.A.; Madu, C.E.; Aiyegoro, O.A.; Stenström, T.A.; Okoh, A.I. Antibiogram and beta-lactamase genes among cefotaxime resistant E. coli from wastewater treatment plant. Antimicrob. Resist. Infect. Control 2020, 9, 46. [Google Scholar] [CrossRef]
- Bush, K. Past and present perspectives on β-lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum β-lactamases (ESBLs) in the developed world. J. Travel Med. 2017, 24, S44–S51. [Google Scholar] [CrossRef]
- Ur Rahman, S.; Ali, T.; Ali, I.; Khan, N.A.; Han, B.; Gao, J. The Growing Genetic and Functional Diversity of Extended Spectrum Beta-Lactamases. Biomed Res. Int. 2018, 2018, 9519718. [Google Scholar] [CrossRef] [PubMed]
- Mita, Y.; Shigemura, K.; Osawa, K.; Kitagawa, K.; Kotaki, T.; Shirakawa, T.; Miyara, T.; Fujisawa, M. Clinical Risk Factors for Death Caused by Extended-Spectrum Beta-Lactamase: Producing Bacteria. Urol. Int. 2019, 102, 181–186. [Google Scholar] [CrossRef]
- Pandit, R.; Awal, B.; Shrestha, S.S.; Joshi, G.; Rijal, B.P.; Parajuli, N.P. Extended-Spectrum β-Lactamase (ESBL) Genotypes among Multidrug-Resistant Uropathogenic Escherichia coli Clinical Isolates from a Teaching Hospital of Nepal. Interdiscip. Perspect. Infect. Dis. 2020, 2020, 6525826. [Google Scholar] [CrossRef] [Green Version]
- Jena, J.; Sahoo, R.K.; Debata, N.K.; Subudhi, E. Prevalence of TEM, SHV, and CTX-M genes of extended-spectrum β-lactamase-producing Escherichia coli strains isolated from urinary tract infections in adults. Biotech 2017, 7, 244. [Google Scholar] [CrossRef] [PubMed]
- Mirkalantari, S.; Masjedian, F.; Irajian, G.; Siddig, E.E.; Fattahi, A. Determination of the frequency of β-lactamase genes (bla SHV, bla TEM, bla CTX-M) and phylogenetic groups among ESBL-producing uropathogenic Escherichia coli isolated from outpatients. J. Lab. Med. 2020, 44, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Alqasim, A.; Abu Jaffal, A.; Alyousef, A.A. Prevalence of multidrug resistance and extended-spectrum β -Lactamase carriage of clinical uropathogenic Escherichia coli isolates in Riyadh, Saudi Arabia. Int. J. Microbiol. 2018, 2018, 3026851. [Google Scholar] [CrossRef] [Green Version]
- Ghafourian, S.; Sadeghifard, N.; Soheili, S.; Sekawi, Z. Extended spectrum beta-lactamases: Definition, classification and epidemiology. Curr. Issues Mol. Biol. 2014, 17, 11–22. [Google Scholar]
- Ashok, A.K.; Jaryal, S.C.; Thakur, K.; Sood, A.; Gupta, P.K.; Thakur, S. Detection of Inducible and Non-inducible (constitutive) AmpC β-lactamase-producing Gram-Negative Bacteria among Family Enterobacteriaceae by Two Phenotypic Methods-Disk Antagonism Test (DAT) and AmpC disk Test at a Tertiary Care Hospital, Himachal Pradesh. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Antunes, N.T.; Lamoureaux, T.L.; Toth, M.; Stewart, N.K.; Frase, H.; Vakulenko, S.B. Class D β-lactamases: Are they all carbapenemases? Antimicrob. Agents Chemother. 2014, 58, 2119–2125. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.H.; Lee, J.H.; Lee, J.J.; Park, K.S.; Karim, A.M.; Lee, C.R.; Jeong, B.C.; Lee, S.H. Structural basis for carbapenem-hydrolyzing mechanisms of carbapenemases conferring antibiotic resistance. Int. J. Mol. Sci. 2015, 16, 9654–9692. [Google Scholar] [CrossRef]
- Kotsakis, S.D.; Flach, C.F.; Razavi, M.; Joakim Larsson, D.G. Characterization of the first OXA-10 natural variant with increased carbapenemase activity. Antimicrob. Agents Chemother. 2019, 63, e01817-18. [Google Scholar] [CrossRef] [Green Version]
- Shaikh, S.; Fatima, J.; Shakil, S.; Rizvi, S.M.D.; Kamal, M.A. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J. Biol. Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Bogner, C.; Miethke, T.; Wantia, N.; Gebhard, F.; Busch, D.; Hoffmann, R. Differences in ESBL Genes between E. coli, Klebsiella spp. and Enterobacter Cloacae Strains. Int. J. Clin. Med. Microbiol. 2016, 1, 106. [Google Scholar] [CrossRef]
- Dangelo, R.G.; Johnson, J.K.; Bork, J.T.; Heil, E.L. Treatment options for extended-spectrum beta-lactamase (ESBL) and AmpC-producing bacteria. Expert Opin. Pharmacother. 2016, 17, 953–967. [Google Scholar] [CrossRef]
- Bajaj, P.; Singh, N.S.; Virdi, J.S. Escherichia coli β-lactamases: What really matters. Front. Microbiol. 2016, 7, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bevan, E.R.; Jones, A.M.; Hawkey, P.M. Global epidemiology of CTX-M β-lactamases: Temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 2017, 72, 2145–2155. [Google Scholar] [CrossRef] [Green Version]
- Nzima, B.; Adegoke, A.A.; Ofon, U.A.; Al-Dahmoshi, H.O.M.; Saki, M.; Ndubuisi-Nnaji, U.U.; Inyang, C.U. Resistotyping and extended-spectrum beta-lactamase genes among Escherichia coli from wastewater treatment plants and recipient surface water for reuse in South Africa. New Microbes New Infect. 2020, 38, 100803. [Google Scholar] [CrossRef]
- Hocquet, D.; Muller, A.; Bertrand, X. What happens in hospitals does not stay in hospitals: Antibiotic-resistant bacteria in hospital wastewater systems. J. Hosp. Infect. 2016, 93, 395–402. [Google Scholar] [CrossRef]
- King, T.L.B.; Schmidt, S.; Essack, S.Y. Antibiotic resistant Klebsiella spp. from a hospital, hospital effluents and wastewater treatment plants in the uMgungundlovu District, KwaZulu-Natal, South Africa. Sci. Total Environ. 2020, 712, 135550. [Google Scholar] [CrossRef]
- Rodriguez-Mozaz, S.; Chamorro, S.; Marti, E.; Huerta, B.; Gros, M.; Sànchez-Melsió, A.; Borrego, C.M.; Barceló, D.; Balcázar, J.L. Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river. Water Res. 2015, 69, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Kim, T.G.; Cho, K.S. Fate and behavior of extended-spectrum β-lactamase-producing genes in municipal sewage treatment plants. J. Environ. Sci. Health Part A Toxic/Hazard. Subst. Environ. Eng. 2015, 50, 1160–1168. [Google Scholar] [CrossRef]
- Aubertheau, E.; Stalder, T.; Mondamert, L.; Ploy, M.C.; Dagot, C.; Labanowski, J. Impact of wastewater treatment plant discharge on the contamination of river biofilms by pharmaceuticals and antibiotic resistance. Sci. Total Environ. 2017, 579, 1387–1398. [Google Scholar] [CrossRef]
- Sims, N.; Kasprzyk-Hordern, B. Future perspectives of wastewater-based epidemiology: Monitoring infectious disease spread and resistance to the community level. Environ. Int. 2020, 139, 105689. [Google Scholar] [CrossRef]
- Fernando, D.M.; Tun, H.M.; Poole, J.; Patidar, R.; Li, R.; Mi, R.; Amarawansha, G.E.A.; Dilantha Fernando, W.G.; Khafipour, E.; Farenhorst, A.; et al. Detection of antibiotic resistance genes in source and drinking water samples from a first nations community in Canada. Appl. Environ. Microbiol. 2016, 82, 4767–4775. [Google Scholar] [CrossRef] [Green Version]
- Conte, D.; Palmeiro, J.K.; da Silva Nogueira, K.; de Lima, T.M.R.; Cardoso, M.A.; Pontarolo, R.; Degaut Pontes, F.L.; Dalla-Costa, L.M. Characterization of CTX-M enzymes, quinolone resistance determinants, and antimicrobial residues from hospital sewage, wastewater treatment plant, and river water. Ecotoxicol. Environ. Saf. 2017, 136, 62–69. [Google Scholar] [CrossRef]
- Osińska, A.; Korzeniewska, E.; Harnisz, M.; Niestępski, S. The prevalence and characterization of antibiotic-resistant and virulent Escherichia coli strains in the municipal wastewater system and their environmental fate. Sci. Total Environ. 2017, 577, 367–375. [Google Scholar] [CrossRef]
- South African Department of Water and Environmental Affairs National Water Act 36 of 1998. In Wastewater Limit Values Applicable to the Irrigation of Any Land or Property up to 50 Cubic Metres; Government Gazette No. 19182; Department of Water Affairs: Pretoria, South Africa, 2013; pp. 17–18.
- South African National Department of Health. Standard Treatment Guidelines and Essential Medicines List for South Africa: Hospital Level (Adults), 4th ed.; National Deaprtment of Health: Pretoria, South Africa, 2015; ISBN 9781920031718.
- Kubone, P.Z.; Mlisana, K.P.; Govinden, U.; Luther, A.; Abia, K.; Essack, S.Y. Antibiotic Susceptibility and Molecular Characterization of Uropathogenic Escherichia coli Associated with Community-Acquired Urinary Tract Infections in Urban and Rural Settings in South Africa. Trop. Med. Infect. Dis. 2020, 5, 1–13. [Google Scholar]
- McIver, K.S.; Amoako, D.G.; Abia, A.L.K.; Bester, L.A.; Chenia, H.Y.; Essack, S.Y. Molecular epidemiology of antibiotic-resistant escherichia coli from farm-to-fork in intensive poultry production in KwaZulu-Natal, South Africa. Antibiotics 2020, 9, 850. [Google Scholar] [CrossRef]
- Abdalla, S.E.; Abia, A.L.K.; Amoako, D.G.; Perrett, K.; Bester, L.A.; Essack, S.Y. From Farm-to-Fork: E. Coli from an Intensive Pig Production System in South Africa Shows High Resistance to Critically Important Antibiotics for Human and Animal Use. Antibiotics 2021, 10, 178. [Google Scholar]
- Mbanga, J.; Amoako, D.G.; Abia, A.L.K.; Allam, M.; Ismail, A.; Essack, S.Y. Genomic Insights of Multidrug-Resistant Escherichia coli From Wastewater Sources and Their Association With Clinical Pathogens in South Africa. Front. Vet. Sci. 2021, 8, 137. [Google Scholar] [CrossRef]
- Shi, P.; Jia, S.; Zhang, X.X.; Zhang, T.; Cheng, S.; Li, A. Metagenomic insights into chlorination effects on microbial antibiotic resistance in drinking water. Water Res. 2013, 47, 111–120. [Google Scholar] [CrossRef]
- Kutilova, I.; Medvecky, M.; Leekitcharoenphon, P.; Munk, P.; Masarikova, M.; Davidova-Gerzova, L.; Jamborova, I.; Bortolaia, V.; Pamp, S.J.; Dolejska, M. Extended-spectrum beta-lactamase-producing Escherichia coli and antimicrobial resistance in municipal and hospital wastewaters in Czech Republic: Culture-based and metagenomic approaches. Environ. Res. 2021, 193, 110487. [Google Scholar] [CrossRef]
- Liu, S.S.; Qu, H.M.; Yang, D.; Hu, H.; Liu, W.L.; Qiu, Z.G.; Hou, A.M.; Guo, J.; Li, J.W.; Shen, Z.Q.; et al. Chlorine disinfection increases both intracellular and extracellular antibiotic resistance genes in a full-scale wastewater treatment plant. Water Res. 2018, 136, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connelly, C.M.; Ph, D.; Torres, M.P.; Ph, D.; Lechner, J.; Porter, L.R.; Morrow, S.; Ascp, M.T.; Hanson, N.D. Plasmid-mediated β-lactamases: Nomadic microbial assassins lurking in multidrug resistant Gram-negative strains. Biotechniques 2013, 55, 212. [Google Scholar] [CrossRef] [Green Version]
- Korzeniewska, E.; Harnisz, M. Extended-spectrum beta-lactamase (ESBL)-positive Enterobacteriaceae in municipal sewage and their emission to the environment. J. Environ. Manag. 2013, 128, 904–911. [Google Scholar] [CrossRef]
- Ojer-Usoz, E.; González, D.; García-Jalón, I.; Vitas, A.I. High dissemination of extended-spectrum β-lactamase-producing Enterobacteriaceae ineffluents from wastewater treatment plants. Water Res. 2014, 56, 37–47. [Google Scholar] [CrossRef]
- Njage, P.M.K.K.; Buys, E.M. Pathogenic and commensal Escherichiacoli from irrigation water show potential in transmission of extended spectrum and AmpC ? ²-lactamases determinants to isolates from lettuce. Microb. Biotechnol. 2015, 8, 462–473. [Google Scholar]
- Li, L.; Nesme, J.; Quintela-Baluja, M.; Balboa, S.; Hashsham, S.; Williams, M.R.; Yu, Z.; Sørensen, S.J.; Graham, D.W.; Romalde, J.L.; et al. Extended-Spectrum β-Lactamase and Carbapenemase Genes are Substantially and Sequentially Reduced during Conveyance and Treatment of Urban Sewage. Environ. Sci. Technol. 2021, 55, 5939–5949. [Google Scholar] [CrossRef]
- Bréchet, C.; Plantin, J.; Sauget, M.; Thouverez, M.; Talon, D.; Cholley, P.; Guyeux, C.; Hocquet, D.; Bertrand, X. Wastewater treatment plants release large amounts of extended-spectrum β-lactamase-producing escherichia coli into the environment. Clin. Infect. Dis. 2014, 58, 1658–1665. [Google Scholar] [CrossRef]
- Li, Q.; Chang, W.; Zhang, H.; Hu, D.; Wang, X. The role of plasmids in the multiple antibiotic resistance transfer in ESBLs-producing Escherichia coli isolated from wastewater treatment plants. Front. Microbiol. 2019, 10, 633. [Google Scholar] [CrossRef]
- Pärnänen, K.M.M.; Narciso-Da-Rocha, C.; Kneis, D.; Berendonk, T.U.; Cacace, D.; Do, T.T.; Elpers, C.; Fatta-Kassinos, D.; Henriques, I.; Jaeger, T.; et al. Antibiotic resistance in European wastewater treatment plants mirrors the pattern of clinical antibiotic resistance prevalence. Sci. Adv. 2019, 5, eaau9124. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO List of Critically Important Antimicrobials for Human Medicine (WHO CIA List); World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Suarez, S.; Lema, J.M.; Omil, F. Pre-treatment of hospital wastewater by coagulation-flocculation and flotation. Bioresour. Technol. 2009, 100, 2138–2146. [Google Scholar] [CrossRef]
- Amos, G.C.A.; Hawkey, P.M.; Gaze, W.H.; Wellington, E.M. Waste water effluent contributes to the dissemination of CTX-M-15 in the natural environment. J. Antimicrob. Chemother. 2014, 69, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Dolejska, M.; Frolkova, P.; Florek, M.; Jamborova, I.; Purgertova, M.; Kutilova, I.; Cizek, A.; Guenther, S.; Literak, I. CTX-M-15-producing Escherichia coli clone B2-O25b-ST131 and Klebsiella spp. isolates in municipal wastewater treatment plant effluents. J. Antimicrob. Chemother. 2011, 66, 2784–2790. [Google Scholar]
- D’Andrea, M.M.; Arena, F.; Pallecchi, L.; Rossolini, G.M. CTX-M-type β-lactamases: A successful story of antibiotic resistance. Int. J. Med. Microbiol. 2013, 303, 305–317. [Google Scholar] [CrossRef]
- Palzkill, T. Structural and mechanistic basis for extended-spectrum drug-resistance mutations in altering the specificity of TEM, CTX-M, and KPC β-lactamases. Front. Mol. Biosci. 2018, 5, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Alfaresi, M.; Kim Sing, G.; Senok, A. First Report of bla CTX-M-28 in Enterobacteriaceae Isolates in the United Arab Emirates. J. Pathog. 2018, 2018, 1304793. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.H.; Hu, Z.Q. Epidemiology and genetics of CTX-M extended-spectrum β-lactamases in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 79–101. [Google Scholar] [CrossRef]
- Raven, K.E.; Ludden, C.; Gouliouris, T.; Blane, B.; Naydenova, P.; Brown, N.M.; Parkhill, J.; Peacock, S.J. Genomic surveillance of Escherichia coli in municipal wastewater treatment plants as an indicator of clinically relevant pathogens and their resistance genes. Microb. Genomics 2019, 5, e000267. [Google Scholar] [CrossRef]
- Paulshus, E.; Thorell, K.; Guzman-Otazo, J.; Joffre, E.; Colque, P.; Kühn, I.; Möllby, R.; Sørum, H.; Sjöling, Å. Repeated isolation of extended-spectrum-β-lactamase-positive Escherichia coli sequence types 648 and 131 from Community wastewater indicates that sewage systems are important sources of emerging clones of antibiotic-resistant bacteria. Antimicrob. Agents Chemother. 2019, 63, e00823-19. [Google Scholar] [CrossRef] [Green Version]
- Bridgewater, L.; APHA/AWWA/WEF. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; Rice, E.W., Bridgewater, L., Eds.; American Public Health Association: Washingdon, DC, USA, 2012; ISBN 0738133175. [Google Scholar]
- Tarr, C.L.; Large, T.M.; Moeller, C.L.; Lacher, D.W.; Tarr, P.I.; Acheson, D.W.; Whittam, T.S. Molecular characterization of a serotype O121:H19 clone, a distinct Shiga toxin-producing clone of pathogenic Escherichia coli. Infect. Immun. 2002, 70, 6853–6859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abia, A.L.K.; Ubomba-Jaswa, E.; Momba, M.N.B. Impact of seasonal variation on Escherichia coli concentrations in the riverbed sediments in the Apies River, South Africa. Sci. Total Environ. 2015, 537, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.; Kirby, W.; Sherris, J.; Turck, M. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 493. [Google Scholar] [CrossRef]
- CLSI Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; Informational Supplement M100-S27; CLSI: Wayne, PA, USA, 2017.
- Ezeanya, C.; Agbakoba, N.; Ejike, C.; Okwelogu, S. Evaluation of a Chromogenic Medium for the Detection of ESBL with Comparison to Double Disk Synergy Test. Br. J. Med. Med. Res. 2017, 21, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Versalovic, J.; Koeuth, T.; Lupski, R. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef] [PubMed]
- Pusparini, N.; Waturangi, D.E.; Usia, T.; Nikastri, E. Genetic diversity of Escherichia coli isolated from ice cube production sites. BMC Res. Notes 2018, 11, 4–11. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antibiotics | A | B | C | D | ||||
|---|---|---|---|---|---|---|---|---|
| Influent | Effluent | Influent | Effluent | Influent | Effluent | Influent | Effluent | |
| FEP | 30 | 30 | 30 | 20 | 10 | 10 | 10 | 20 |
| CAZ | 30 | 30 | 30 | 30 | 10 | 10 | 10 | 20 |
| CTX | 30 | 50 | 40 | 90 | 20 | 70 | 60 | 90 |
| CFM | 40 | 50 | 40 | 90 | 20 | 60 | 50 | 90 |
| FOX | 50 | 20 | 30 | 20 | 10 | 10 | 10 | 20 |
| LEX | 50 | 60 | 80 | 80 | 30 | 60 | 80 | 100 |
| CXM | 30 | 60 | 50 | 100 | 20 | 90 | 60 | 90 |
| CEF | 30 | 60 | 60 | 100 | 30 | 80 | 70 | 100 |
| MEM | 30 | 10 | 20 | 20 | 10 | 10 | 0 | 20 |
| IPM | 30 | 10 | 20 | 20 | 10 | 10 | 0 | 20 |
| AMK | 20 | 10 | 20 | 20 | 10 | 10 | 0 | 20 |
| GEN | 20 | 20 | 20 | 20 | 10 | 20 | 10 | 20 |
| TGC | 30 | 20 | 20 | 20 | 10 | 10 | 0 | 20 |
| TET | 30 | 80 | 60 | 100 | 20 | 100 | 80 | 90 |
| NAL | 40 | 80 | 50 | 90 | 30 | 70 | 70 | 90 |
| CIP | 20 | 50 | 30 | 90 | 20 | 60 | 70 | 80 |
| CHL | 50 | 20 | 20 | 0 | 10 | 20 | 0 | 10 |
| NIT | 20 | 10 | 0 | 10 | 10 | 10 | 0 | 10 |
| SXT | 70 | 90 | 60 | 100 | 40 | 90 | 90 | 90 |
| AMC | 50 | 20 | 30 | 20 | 20 | 20 | 10 | 30 |
| AMX | 70 | 90 | 80 | 100 | 80 | 100 | 90 | 90 |
| PIP | 30 | 70 | 30 | 80 | 40 | 60 | 60 | 80 |
| AMP | 90 | 90 | 90 | 100 | 80 | 100 | 100 | 100 |
| β-Lactamase Enzymes Detected | ||||
|---|---|---|---|---|
| WWTP | Sampling Point | CTX-M-Type Variants (n) | SHV-Type Variants (n) | TEM-Type Variants (n) |
| A | Influent | CTX-M-15 (2) | SHV-28 (2) | TEM-181 (2) TEM-213 (1) |
| Effluent | CTX-M-15 (3) CTX-M-3 (1) | SHV-28 (1) | TEM-181 (2) | |
| B | Influent | CTX-M-15 (1) | Not detected | TEM-116 (1) TEM-1 (1) |
| Effluent | CTX-M-15 (3) | SHV-28 (2) | TEM-181 (1) TEM-215 (1) | |
| C | Influent | CTX-M-15(1) CTX-M-3 (1) | Not detected | TEM-1 (1) TEM-181 (1) |
| Effluent | CTX-M-28 (1) CTX-M-15 (3) | SHV-28 (1) | TEM-1 (3) | |
| D | Influent | Not detected | Not detected | TEM-1 (2) |
| Effluent | CTX-M-3 (1) CTX-M-28 (1) CTX-M-15 (2) | SHV-28 (2) | TEM-181 (1) | |
| TOTAL | 20 | 8 | 17 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gumede, S.N.; Abia, A.L.K.; Amoako, D.G.; Essack, S.Y. Analysis of Wastewater Reveals the Spread of Diverse Extended-Spectrum β-Lactamase-Producing E. coli Strains in uMgungundlovu District, South Africa. Antibiotics 2021, 10, 860. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070860
Gumede SN, Abia ALK, Amoako DG, Essack SY. Analysis of Wastewater Reveals the Spread of Diverse Extended-Spectrum β-Lactamase-Producing E. coli Strains in uMgungundlovu District, South Africa. Antibiotics. 2021; 10(7):860. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070860
Chicago/Turabian StyleGumede, Siyabonga N., Akebe L. K. Abia, Daniel G. Amoako, and Sabiha Y. Essack. 2021. "Analysis of Wastewater Reveals the Spread of Diverse Extended-Spectrum β-Lactamase-Producing E. coli Strains in uMgungundlovu District, South Africa" Antibiotics 10, no. 7: 860. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070860