
Subinhibitory Antibiotic Concentrations Enhance Biofilm Formation of Clinical Enterococcus faecalis Isolates

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates, Antibiotics and Inoculation
2.2. Biofilm Formation Evaluation
2.3. Statistical Analysis
3. Results
4. Discussion
4.1. Subinhibitory Concentrations of Different Antibiotics Increase Biofilm Production in E. faecalis
4.2. Differences between Cell Wall Synthesis and Protein Synthesis Inhibitors
4.3. E. faecalis from Endodontic Lesions and the Higher Risks of Sub-MIC Exposure
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparna, M.S.; Yadav, S. Biofilms: Microbes and disease. Braz. J. Infect. Dis. 2008, 12, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubovics, N.S.; Goodman, S.D.; Mashburn-Warren, L.; Stafford, P.G.; Cieplik, F. The dental plaque biofilm matrix. Periodontology 2000 2021, 86, 32–56. [Google Scholar] [CrossRef]
- Al-Ahmad, A.; Bucher, M.; Anderson, A.C.; Tennert, C.; Hellwig, E.; Wittmer, A.; Vach, K.; Karygianni, L. Antimicrobial Photoinactivation Using Visible Light Plus Water-Filtered Infrared-A (VIS + wIRA) Alters In Situ Oral Biofilms. PLoS ONE 2015, 10, e0132107. [Google Scholar] [CrossRef]
- Dufour, D.; Leung, V.; Lévesque, C.M. Bacterial biofilm: Structure, function, and antimicrobial resistance. Endod. Top. 2012, 2–16. [Google Scholar] [CrossRef]
- Do, T.; Devine, D.; Marsh, P.D. Oral biofilms: Molecular analysis, challenges, and future prospects in dental diagnostics. Clin. Cosmet. Investig. Dent. 2013, 5, 11–19. [Google Scholar]
- Bernardi, S.; Continenza, M.A.; Al-Ahmad, A.; Karygianni, L.; Follo, M.; Filippi, A.; Macchiarelli, G. Streptococcus spp. and Fusobacterium nucleatum in tongue dorsum biofilm from halitosis patients: A fluorescence in situ hybridization (FISH) and confocal laser scanning microscopy (CLSM) study. New Microbiol. 2019, 42, 108–113. [Google Scholar]
- Bernardi, S.; Bianchi, S.; Botticelli, G.; Rastelli, E.; Tomei, A.R.; Palmerini, M.G.; Continenza, M.A.; Macchiarelli, G. Scanning electron microscopy and microbiological approaches for the evaluation of salivary microorganisms behaviour on anatase titanium surfaces: In vitro study. Morphologie 2018, 102, 1–6. [Google Scholar] [CrossRef]
- Bianchi, S.; Fantozzi, G.; Bernardi, S.; Antonouli, S.; Continenza, M.A.; Macchiarelli, G. Commercial oral hygiene products and implant collar surfaces: Scanning electron microscopy observations. Can. J. Dent. Hyg. 2020, 54, 26–31. [Google Scholar]
- Bernardi, S.; Bianchi, S.; Tomei, A.R.; Continenza, M.A.; Macchiarelli, G. Microbiological and SEM-EDS evaluation of titanium surfaces exposed to periodontal gel: In vitro study. Materials 2019, 12, 1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.J.; Parks, P.J.; Peterson, M.L. A mucosal model to study microbial biofilm development and anti-biofilm therapeutics. J. Microbiol. Methods 2013, 92, 201–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endodontists, A.A. of AAE Position Statement: AAE Guidance on the Use of Systemic Antibiotics in Endodontics. J. Endod. 2017, 43, 1409–1413. [Google Scholar]
- Penesyan, A.; Paulsen, I.T.; Gillings, M.R.; Kjelleberg, S.; Manefield, M.J. Secondary Effects of Antibiotics on Microbial Biofilms. Front. Microbiol. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Kowalska-Krochmal, B.; Dudek-Wicher, R. The Minimum Inhibitory Concentration of Antibiotics: Methods, Interpretation, Clinical Relevance. Pathogens 2021, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Vasilchenko, A.S.; Rogozhin, E.A. Sub-inhibitory Effects of Antimicrobial Peptides. Front. Microbiol. 2019, 10, 1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Guo, Y.; Zhan, Q.; Shang, Y.; Qu, D.; Yu, F. Subinhibitory concentrations of mupirocin stimulate Staphylococcus aureus biofilm formation by upregulating cidA. Antimicrob. Agents Chemother. 2020, 64, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, R.; Sato, T.; Nomura, N.; Nakamura, T.; Senpuku, H. Potential Risk of Spreading Resistance Genes within Extracellular-DNA-Dependent Biofilms of Streptococcus mutans in Response to Cell Envelope Stress Induced by Sub-MICs of Bacitracin. Appl. Environ. Microbiol. 2020, 86, e00770-20. [Google Scholar] [CrossRef]
- Haapasalo, M.; Endal, U.; Zandi, H.; Coil, J.M. Eradication of endodontic infection by instrumentation and irrigation solutions. Endod. Top. 2005, 10, 77–102. [Google Scholar] [CrossRef]
- Pizzo, G.; Guiglia, R.; Lo Russo, L.; Campisi, G. Dentistry and internal medicine: From the focal infection theory to the periodontal medicine concept. Eur. J. Intern. Med. 2010, 21, 496–502. [Google Scholar] [CrossRef]
- Varvara, G.; Bernardi, S.; Piattelli, M.; Cutilli, T. Rare and life-threatening complication after an attempted lower third molar extraction: Lemierre syndrome. Ann. R. Coll. Surg. Engl. 2019, 101, e52–e54. [Google Scholar] [CrossRef] [PubMed]
- Holland, R.; Eduardo, J.; Filho, G.; Tavares, L.; Cintra, A. Factors affecting the periapical healing process of endodontically treated teeth. J. Appl. Oral Sci. 2017, 25, 465–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, E.Y.; Cho, Y.; Lee, J.Y.; Kim, S.J.; Choi, J. Endodontic treatment enhances the regenerative potential of teeth with advanced periodontal disease with secondary endodontic involvement. J. Periodontal Implant. Sci. 2013, 43, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Sculean, A.; Stavropoulos, A.; Bosshardt, D.D. Self-regenerative capacity of intra-oral bone defects. J. Clin. Periodontol. 2019, 46, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, A.C.; Hellwig, E.; Vespermann, R.; Wittmer, A.; Schmid, M.; Karygianni, L.; Al-Ahmad, A. Comprehensive analysis of secondary dental root canal infections: A combination of culture and culture-independent approaches reveals new insights. PLoS ONE 2012, 7, e49576. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.C.; Al-Ahmad, A.; Elamin, F.; Jonas, D.; Mirghani, Y.; Schilhabel, M.; Karygianni, L.; Hellwig, E.; Rehman, A. Comparison of the bacterial composition and structure in symptomatic and asymptomatic endodontic infections associated with root-filled teeth using pyrosequencing. PLoS ONE 2013, 8, e84960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ahmad, A.; Auschill, T.M.; Braun, G.; Hellwig, E.; Arweiler, N.B. Overestimation of Streptococcus mutans prevalence by nested PCR detection of the 16S rRNA gene. J. Med. Microbiol. 2006, 55, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ahmad, A.; Ameen, H.; Pelz, K.; Karygianni, L.; Wittmer, A.; Anderson, A.C.; Spitzm, B.; Hellwig, E. Antibiotic Resistance and Capacity for Biofilm Formation of Different Bacteria Isolated from Endodontic Infections Associated with Root-filled Teeth. J. Endod. 2014, 40, 223–230. [Google Scholar] [CrossRef]
- Anderson, A.C.; Jonas, D.; Huber, I.; Karygianni, L.; Wölber, J.; Hellwig, E.; Arweiler, N.; Vach, K.; Wittmer, A.; Al-Ahmad, A. Enterococcus faecalis from Food, Clinical Specimens, and Oral Sites: Prevalence of Virulence Factors in Association with Biofilm Formation. Front. Microbiol. 2016, 6, 1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard, 9th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- CLSI. Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria: Approved Standard, 8th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Hickl, J.; Argyropoulou, A.; Sakavitsi, M.E.; Halabalaki, M.; Al-Ahmad, A.; Hellwig, E.; Aligiannis, N.; Skaltsounis, A.L.; Wittmer, A.; Vach, K.; et al. Mediterranean herb extracts inhibit microbial growth of representative oral microorganisms and biofilm formation of Streptococcus mutans. PLoS ONE 2018, 13, e0207574. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef]
- Allkja, J.; van Charante, F.; Aizawa, J.; Reigada, I.; Guarch-Pérez, C.; Vazquez-Rodriguez, J.A.; Cos, P.; Coenye, T.; Fallarero, A.; Zaat, S.; et al. Interlaboratory study for the evaluation of three microtiter plate-based biofilm quantification methods. Sci. Rep. 2021, 11, 13779. [Google Scholar] [CrossRef]
- Tang, Q.; Hao, Y.; Wang, L.; Lu, C.; Li, M.; Si, Z.; Wu, X.; Lu, Z. Characterization of a bacterial strain Lactobacillus paracasei LP10266 recovered from an endocarditis patient in Shandong, China. BMC Microbiol. 2021, 21, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Dale, J.L.; Nilson, J.L.; Barnes, A.M.T.; Dunny, G.M. Restructuring of Enterococcus faecalis biofilm architecture in response to antibiotic-induced stress. NPJ Biofilms Microbiomes 2017, 3, 15–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waack, U.; Nicholson, T.L. Subinhibitory Concentrations of Amoxicillin, Lincomycin, and Oxytetracycline Commonly Used to Treat Swine Increase Streptococcus suis Biofilm Formation. Front. Microbiol. 2018, 9, 2707. [Google Scholar] [CrossRef] [Green Version]
- Poimenidou, S.V.; Dalmasso, M.; Papadimitriou, K.; Fox, E.M.; Skandamis, P.N.; Jordan, K. Virulence Gene Sequencing Highlights Similarities and Differences in Sequences in Listeria monocytogenes Serotype 1:2a and 4b Strains of Clinical and Food Origin From 3 Different Geographic Locations. Front. Microbiol. 2018, 9, 1103. [Google Scholar] [CrossRef]
- Hathroubi, S.; Mekni, M.A.; Domenico, P.; Nguyen, D.; Jacques, M. Biofilms: Microbial Shelters Against Antibiotics. Microb. Drug Resist. 2017, 23, 147–156. [Google Scholar] [CrossRef]
- Cepas, V.; López, Y.; Muñoz, E.; Rolo, D.; Ardanuy, C.; Martí, S.; Xercavins, M.; Horcajada, J.P.; Bosch, J.; Soto, S.M. Relationship Between Biofilm Formation and Antimicrobial Resistance in Gram-Negative Bacteria. Microb. Drug Resist. 2019, 25, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Hallinen, M.K.; Wood, K.B. Interplay between Antibiotic Efficacy and Drug-Induced Lysis Underlies Enhanced Biofilm Formation at Subinhibitory Drug Concentrations. Antimicrob. Agents Chemother. 2018, 62, e01603-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aka, S.T.; Haji, S.H. Sub-MIC of antibiotics induced biofilm formation of Pseudomonas aeruginosa in the presence of chlorhexidine. Braz. J. Microbiol. 2015, 46, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Tong, Z.; Huang, L.; Ling, J.; Mao, X.; Ning, Y.; Deng, D. Effects of Intracanal Irrigant MTAD Combined with Nisin at Sub-Minimum Inhibitory Concentration Levels on Enterococcus faecalis Growth and the Expression of Pathogenic Genes. PLoS ONE 2014, 9, e90235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korir, L.M.; Dale, L.J.; Dunny, M.G. Role of epaQ, a Previously Uncharacterized Enterococcus faecalis Gene, in Biofilm Development and Antimicrobial Resistance. J. Bacteriol. 2019, 201, e00078-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, J.L.; Cagnazzo, J.; Phan, C.Q.; Barnes, A.M.T.; Dunny, G.M. Multiple roles for Enterococcus faecalis glycosyltransferases in biofilm-associated antibiotic resistance, cell envelope integrity, and conjugative transfer. Antimicrob. Agents Chemother. 2015, 59, 4094–4105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scornec, H.; Bellanger, X.; Guilloteau, H.; Groshenry, G.; Merlin, C. Inducibility of Tn 916 conjugative transfer in Enterococcus faecalis by subinhibitory concentrations of ribosome-targeting antibiotics. J. Antimicrob. Chemother. 2017, 72, 2722–2728. [Google Scholar] [CrossRef]
- Maestre, J.R.; Aguilar, L.; Mateo, M.; Giménez, M.-J.; Méndez, M.-L.; Alou, L.; Granizo, J.-J.; Prieto, J. In vitro interference of tigecycline at subinhibitory concentrations on biofilm development by Enterococcus faecalis. J. Antimicrob. Chemother. 2012, 67, 1155–1158. [Google Scholar] [CrossRef] [PubMed]
- Ranieri, M.R.M.; Whitchurch, C.B.; Burrows, L.L. Mechanisms of biofilm stimulation by subinhibitory concentrations of antimicrobials. Curr. Opin. Microbiol. 2018, 45, 164–169. [Google Scholar] [CrossRef]
- Pinheiro, E.T.; Gomes, B.P.F.A.; Drucker, D.B.; Zaia, A.A.; Ferraz, C.C.R.; Souza-Filho, F.J. Antimicrobial susceptibility of Enterococcus faecalis isolated from canals of root filled teeth with periapical lesions. Int. Endod. J. 2004, 37, 756–763. [Google Scholar] [CrossRef]
- Siqueira Junior, J.F.; Rôças, I.D.N.; Marceliano-Alves, M.F.; Pérez, A.R.; Ricucci, D. Unprepared root canal surface areas: Causes, clinical implications, and therapeutic strategies. Braz. Oral Res. 2018, 32, e65. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.B.; Abdulla, N.; Himratul-Aznita, W.H.; Awad, M.; Samaranayake, L.P.; Ahmed, H.M.A. Antibiotic prescribing practices of dentists for endodontic infections; a cross-sectional study. PLoS ONE 2020, 15, e0244585. [Google Scholar] [CrossRef]
- Bolfoni, M.R.; Pappen, F.G.; Pereira-Cenci, T.; Jacinto, R.C. Antibiotic prescription for endodontic infections: A survey of Brazilian Endodontists. Int. Endod. J. 2018, 51, 148–156. [Google Scholar] [CrossRef]
- Al-Ahmad, A.; Elamin, F.; Gärttner, R.; Anderson, A.; Wittmer, A.; Mirghani, Y.; Hellwig, E. New Bacterial Combinations in Secondary Endodontic Infections of Patients with a Recent Systematic Antibiotic Therapy. Monogr. Oral Sci. 2021, 29, 144–154. [Google Scholar]
- Bernardi, S.; Frascarelli, C.; Fantozzi, G.; Caruso, S.; Gatto, R.; Nardi, G.M.; Continenza, M.A. The Importance of Correct Diagnosis and Treatment in Endo- Periodontal Lesions: A Two Cases Comparison. Dent. Update 2016, 43, 766–769. [Google Scholar] [CrossRef]
- Bernardi, S.; Bianchi, S.; Fantozzi, G.; Leuter, C.; Continenza, M.A.; Macchiarelli, G. Morphometric study on single-root premolars in a European population sample: An update on lengths and diameters. Eur. J. Anat. 2019, 23, 17–25. [Google Scholar]
- Arias-Moliz, M.; Baca, P.; Solana, C.; Toledano, M.; Medina-Castillo, A.; Toledano-Osorio, M.; Osorio, R. Doxycycline-functionalized polymeric nanoparticles inhibit Enterococcus faecalis biofilm formation on dentine. Int. Endod. J. 2021, 54, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Segura-Egea, J.J.; Gould, K.; Şen, B.H.; Jonasson, P.; Cotti, E.; Mazzoni, A.; Sunay, H.; Tjäderhane, L.; Dummer, P.M.H. Antibiotics in Endodontics: A review. Int. Endod. J. 2017, 50, 1169–1184. [Google Scholar] [CrossRef]
- Abbott, P.V.; Hume, W.R.; Pearman, J.W. Antibiotics and endodontics. Aust. Dent. J. 1990, 3, 50–60. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
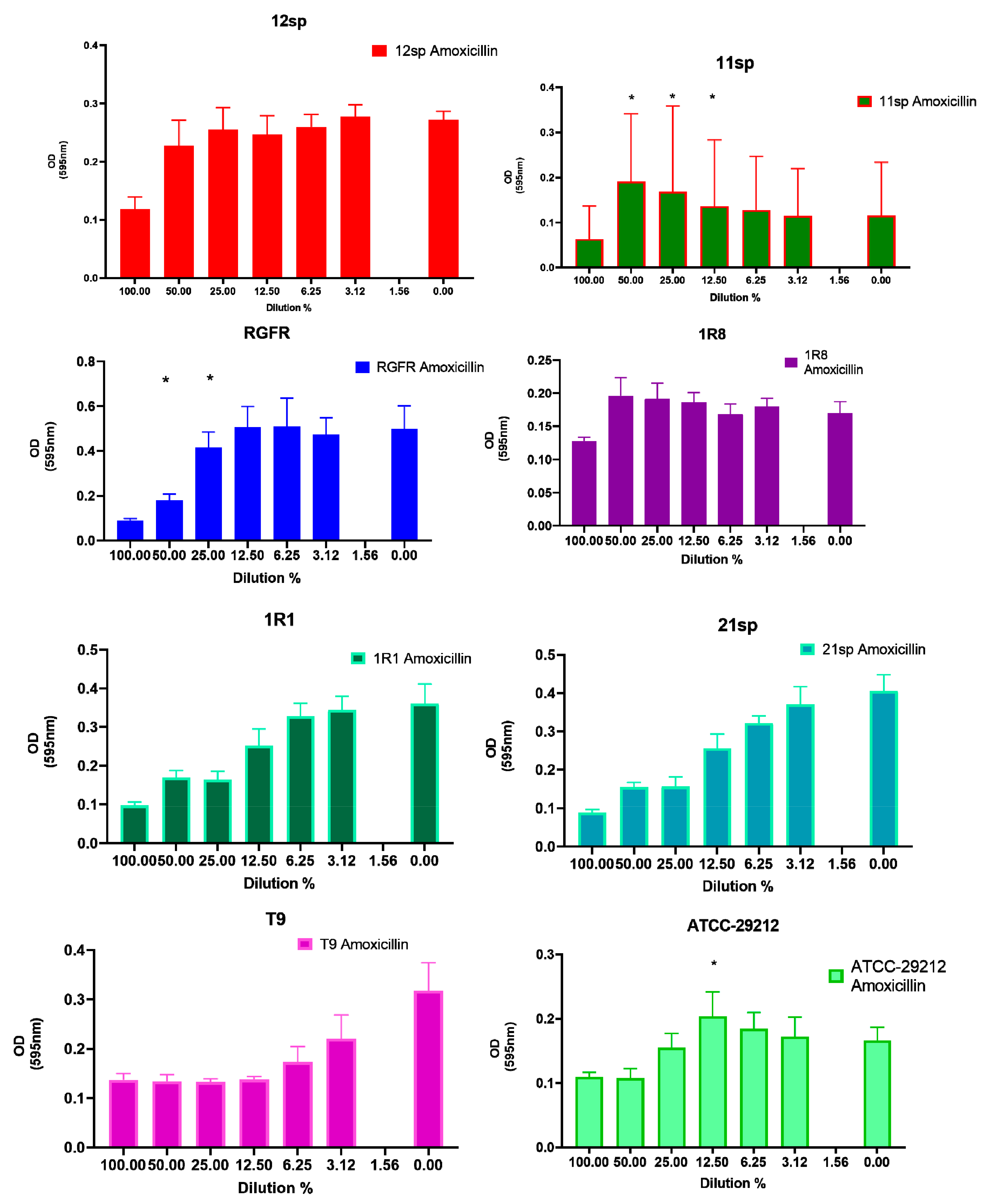
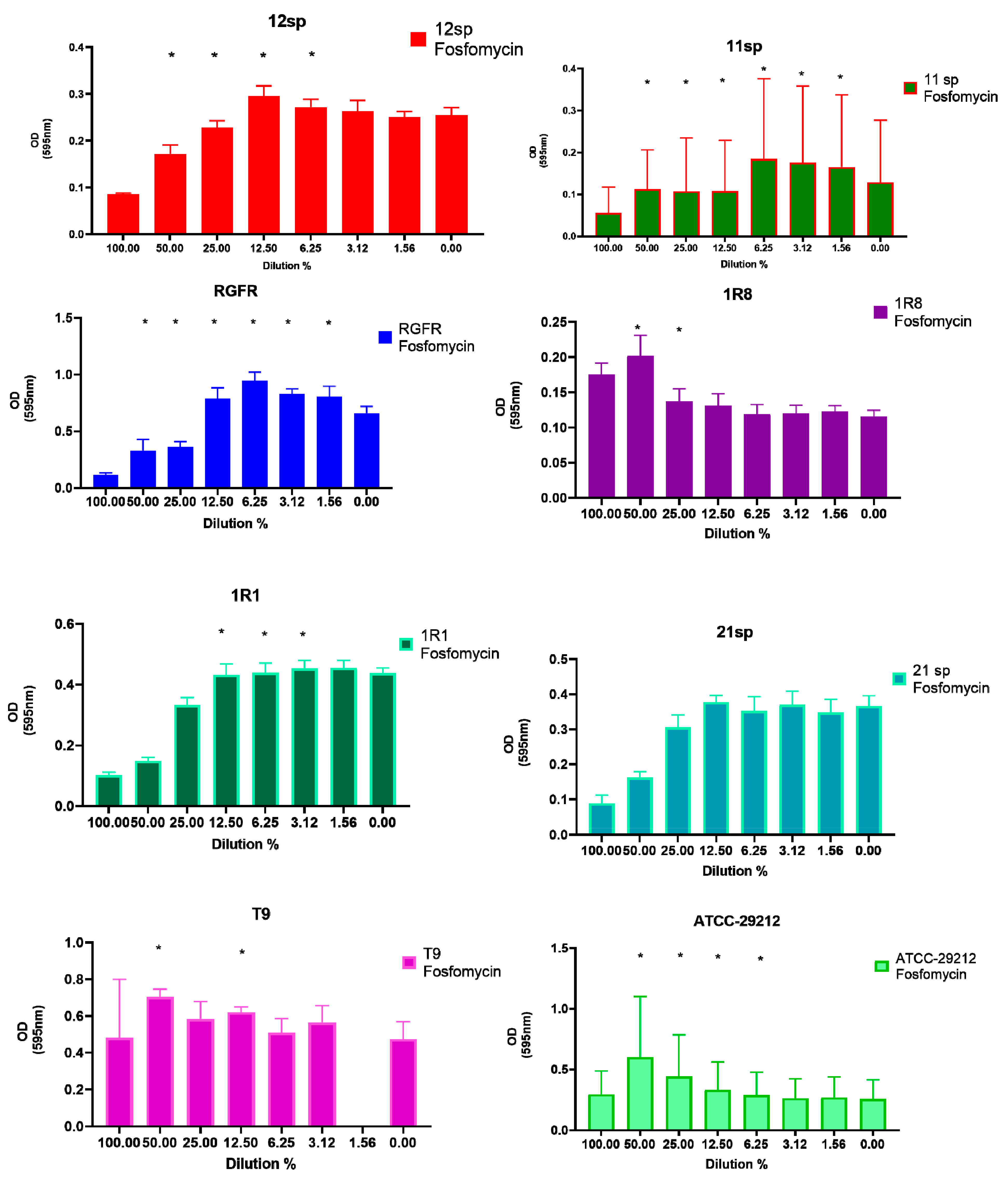
| E. faecalis Isolate Number | Amoxicillin | Doxycycline | Fosfomycin | Penicillin G | Tetracycline | Vancomycin |
|---|---|---|---|---|---|---|
| 12sp | N.S. 1 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; 1:64 MIC–1.56% p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:16 MIC–6.25%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; 1:64 MIC–1.56% p < 0.05 | 1:2 MIC–50%; p < 0.05 |
| 11sp | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; p < 0.05 | 1:2 MIC–50%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; 1:64 MIC–1.56% p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; p < 0.05 | 1:2 MIC–50%; 1:8 MIC–12.5%; p < 0.05 |
| RGFR | 1:2 MIC–50%; 1:4 MIC–25%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; 1:64 MIC–1.56% p < 0.05 | 1:2 MIC–50%; 1:8 MIC–12.5%; p < 0.05 | 1:2 MIC–50%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; p < 0.05 |
| 1R8 | N.S. 1 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; 1:64 MIC–1.56% p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; 1:64 MIC–1.56% p < 0.05 | 1:2 MIC–50%; p < 0.05 |
| 1R1 | N.S. 1 | N.S. 1 | 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; p < 0.05 | N.S. 1 | N.S. 1 | N.S. 1 |
| 21sp | N.S. 1 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; 1:64 MIC–1.56% p < 0.05 | N.S. 1 | N.S. 1 | 1:8 MIC–12.5%; p<0.05 | 1:2 MIC–50%; p < 0.05 |
| T9 2 | N.S. 1 | 1:4 MIC–25%; 1:8 MIC–12.5%; p < 0.05 | 1:2 MIC–50%; 1:8 MIC–12.5%; p < 0.05 | N.S. 1 | 1:4 MIC–25%; 1:16 MIC–6.25%; p < 0.05 | 1:2 MIC–50%; 1:8 MIC–12.5%; p < 0.05 |
| ATCC–29212 | 1:8 MIC–12.5% p < 0.05 | 1:4 MIC–25%; 1:8 MIC–12.5%; p < 0.05 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; p < 0.05 | 1:4 MIC–25%; 1:8 MIC–12.5%; 1:16 MIC–6.25%; 1:32 MIC–3.125%; p < 0.05 | N.S. 1 | 1:2 MIC–50%; 1:4 MIC–25%; 1:8 MIC–12.5%; p < 0.05 |
| Isolate | 1:2 MIC | 1:4 MIC | 1:8 MIC |
|---|---|---|---|
| 11sp | Doxycycline a,c,d Tetracycline b,d Fosfomycin a,b,c Vancomycin b,d | N.S. 1 | N.S. 1 |
| RGFR | Doxycycline b,c Amoxicillin a,c,d Fosfomycin a,b,d Penicillin G a,b,d | Fosfomycin a Amoxicillin b Penicillin G b Vancomycin b | N.S. 1 |
| 1R1 | Doxycycline a Fosfomycin b Vancomycin c | Doxycycline a Fosfomycin b Vancomycin c | Fosfomycin a Vancomycin b |
| 21sp | Tetracycline b Fosfomycin a Vancomycinc | N.S. 1 | N.S. 1 |
| ATCC-29212 | Tetracycline b Amoxicillin a | N.S. 1 | N.S. 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardi, S.; Anderson, A.; Macchiarelli, G.; Hellwig, E.; Cieplik, F.; Vach, K.; Al-Ahmad, A. Subinhibitory Antibiotic Concentrations Enhance Biofilm Formation of Clinical Enterococcus faecalis Isolates. Antibiotics 2021, 10, 874. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070874
Bernardi S, Anderson A, Macchiarelli G, Hellwig E, Cieplik F, Vach K, Al-Ahmad A. Subinhibitory Antibiotic Concentrations Enhance Biofilm Formation of Clinical Enterococcus faecalis Isolates. Antibiotics. 2021; 10(7):874. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070874
Chicago/Turabian StyleBernardi, Sara, Annette Anderson, Guido Macchiarelli, Elmar Hellwig, Fabian Cieplik, Kirstin Vach, and Ali Al-Ahmad. 2021. "Subinhibitory Antibiotic Concentrations Enhance Biofilm Formation of Clinical Enterococcus faecalis Isolates" Antibiotics 10, no. 7: 874. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070874