
In Vitro Synergy of Colistin in Combination with Meropenem or Tigecycline against Carbapenem-Resistant Acinetobacter baumannii

 ,
,  ,
,
Abstract
:1. Introduction
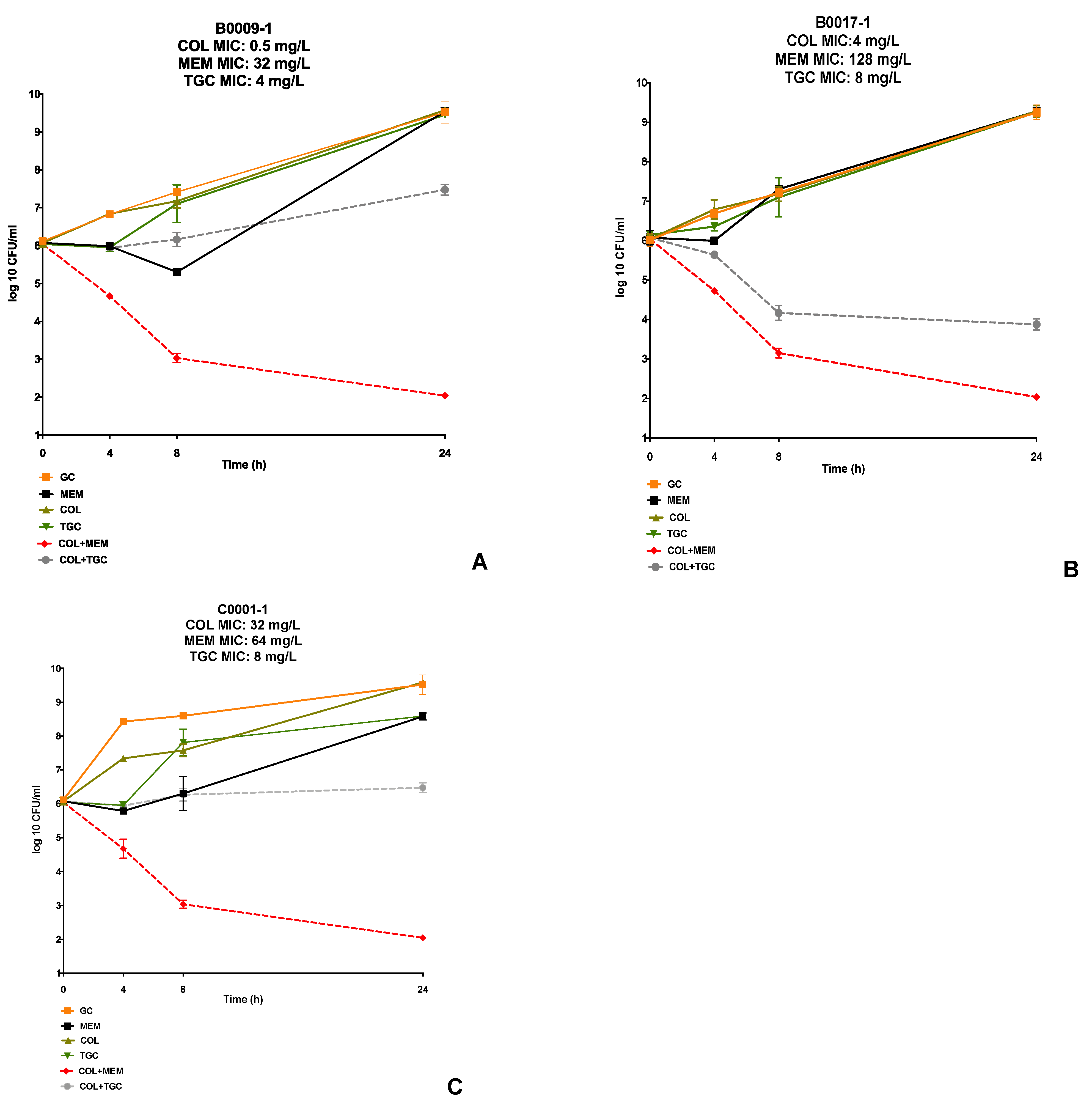
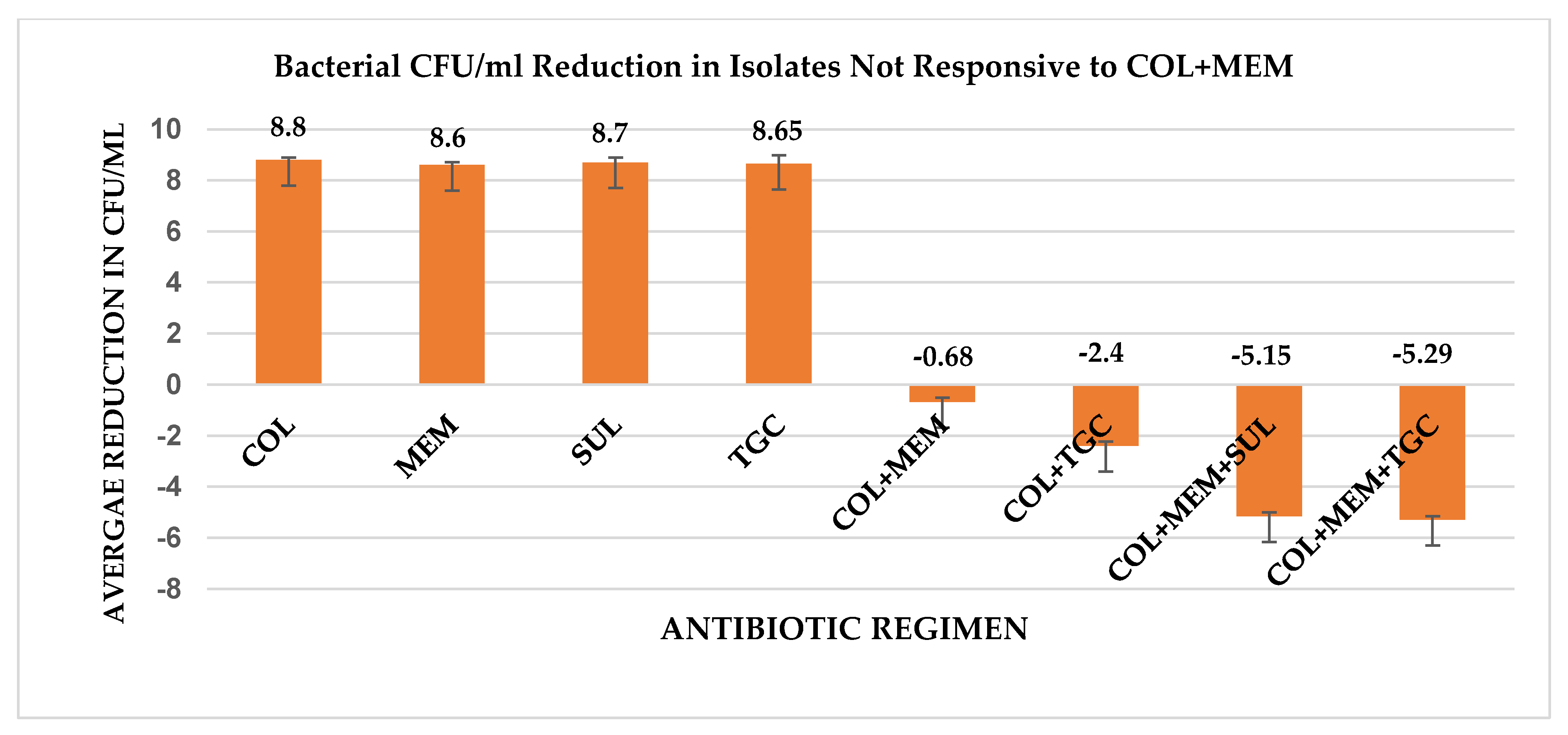
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
Antimicrobials
4.2. Susceptibility Testing
4.3. In Vitro Time-Kill Analysis
4.4. Whole Genome Sequencing Analysis of Antimicrobial Resistance Genes
4.5. Whole Transcriptome Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sievert, D.M.; Ricks, P.; Edwards, J.R.; Schneider, A.; Patel, J.; Srinivasan, A.; Kallen, A.; Limbago, B.; Fridkin, S.; National Healthcare Safety Network (NHSN) Team and Participating NHSN Facilities. Antimicrobial-resistant pathogens associated with healthcare-associated infections: Summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect. Control. Hosp. Epidemiol. 2013, 34, 975. [Google Scholar] [CrossRef]
- Garnacho-Montero, J.; Amaya-Villar, R. Multiresistant Acinetobacter baumannii infections: Epidemiology and management. Curr. Opin. Infect. Dis. 2010, 23, 332–339. [Google Scholar] [CrossRef]
- Pogue, J.M.; Mann, T.; Barber, K.E.; Kaye, K.S. Carbapenem-resistant Acinetobacter baumannii: Epidemiology, surveillance and management. Expert Rev. Anti Infect. Ther. 2013, 11, 383–393. [Google Scholar] [CrossRef]
- Gopalakrishnan, R.; Sureshkumar, D. Changing trends in antimicrobial susceptibility and hospital acquired infections over an 8 year period in a tertiary care hospital in relation to introduction of an infection control programme. J. Assoc. Physicians India 2010, 58, 25–31. [Google Scholar] [PubMed]
- Evans, B.A.; Hamouda, A.; Amyes, S.G. The rise of carbapenem-resistant Acinetobacter baumannii. Curr. Pharm. Des. 2013, 19, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Fishbain, J.; Peleg, A.Y. Treatment of Acinetobacter infections. Clin. Infect. Dis. 2010, 51, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention: Washington, DC, USA, 2019.
- Zilberberg, M.D.; Kollef, M.H.; Shorr, A.F. Secular trends in Acinetobacter baumannii resistance in respiratory and blood stream specimens in the United States, 2003 to 2012: A survey study. J. Hosp. Med. 2016, 11, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, present, and future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Chen, Y.; Fang, Y.; Wang, X.; Chen, Y.; Qi, Q.; Huang, F.; Xiao, X. Meta-analysis of colistin for the treatment of Acinetobacter baumannii infection. Sci. Rep. 2015, 5, 17091. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, Z.A.; Hittle, L.E.; O’Hara, J.A.; Rivera, J.I.; Syed, A.; Shields, R.K.; Pasculle, A.W.; Ernst, R.K.; Doi, Y. Colistin-resistant Acinetobacter baumannii: Beyond carbapenem resistance. Clin. Infect. Dis. 2015, 60, 1295–1303. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, B.T.; Pogue, J.M.; Zavascki, A.P.; Paul, M.; Daikos, G.L.; Forrest, A.; Giacobbe, D.R.; Viscoli, C.; Giamarellou, H.; Karaiskos, I.; et al. International Consensus Guidelines for the Optimal Use of the Polymyxins: Endorsed by the American College of Clinical Pharmacy (ACCP), European Society of Clinical Microbiology and Infectious Diseases (ESCMID), Infectious Diseases Society of America (IDSA), International Society for Anti-infective Pharmacology (ISAP), Society of Critical Care Medicine (SCCM), and Society of Infectious Diseases Pharmacists (SIDP). Pharmacotherapy 2019, 39, 10–39. [Google Scholar] [CrossRef]
- Viehman, J.A.; Nguyen, M.H.; Doi, Y. Treatment options for carbapenem-resistant and extensively drug-resistant Acinetobacter baumannii infections. Drugs 2014, 74, 1315–1333. [Google Scholar] [CrossRef] [Green Version]
- Bergen, P.J.; Tsuji, B.T.; Bulitta, J.B.; Forrest, A.; Jacob, J.; Sidjabat, H.E.; Paterson, D.L.; Nation, R.L.; Li, J. Synergistic killing of multidrug-resistant Pseudomonas aeruginosa at multiple inocula by colistin combined with doripenem in an in vitro pharmacokinetic/pharmacodynamic model. Antimicrob. Agents Chemother. 2011, 55, 5685–5695. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Liu, X.F.; Huang, J.; Zhu, D.M.; Li, J.; Zhang, J. Activities of colistin- and minocycline-based combinations against extensive drug resistant Acinetobacter baumannii isolates from intensive care unit patients. BMC Infect. Dis. 2011, 11, 109. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Chen, C.L.; Wu, S.R.; Huang, C.W.; Chiu, C.H. Risk factors and outcome analysis of acinetobacter baumannii complex bacteremia in critical patients. Crit. Care Med. 2014, 42, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Batirel, A.; Balkan, I.I.; Karabay, O.; Agalar, C.; Akalin, S.; Alici, O.; Alp, E.; Altay, F.A.; Altin, N.; Arslan, F.; et al. Comparison of colistin-carbapenem, colistin-sulbactam, and colistin plus other antibacterial agents for the treatment of extremely drug-resistant Acinetobacter baumannii bloodstream infections. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1311–1322. [Google Scholar] [CrossRef]
- Marchaim, D.; Pogue, J.M.; Tzuman, O.; Hayakawa, K.; Lephart, P.R.; Salimnia, H.; Painter, T.; Zervos, M.J.; Johnson, L.E.; Perri, M.B.; et al. Major variation in MICs of tigecycline in Gram-negative bacilli as a function of testing method. J. Clin. Microbiol. 2014, 52, 1617–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moland, E.S.; Craft, D.W.; Hong, S.G.; Kim, S.Y.; Hachmeister, L.; Sayed, S.D.; Thomson, K.S. In vitro activity of tigecycline against multidrug-resistant Acinetobacter baumannii and selection of tigecycline-amikacin synergy. Antimicrob. Agents Chemother. 2008, 52, 2940–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smani, Y.; Fabrega, A.; Roca, I.; Sanchez-Encinales, V.; Vila, J.; Pachon, J. Role of OmpA in the multidrug resistance phenotype of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2014, 58, 1806–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussi, M.A.; Relling, V.M.; Limansky, A.S.; Viale, A.M. CarO, an Acinetobacter baumannii outer membrane protein involved in carbapenem resistance, is essential for L-ornithine uptake. FEBS Lett. 2007, 581, 5573–5578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. M100-S25 Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Eigth Informational Supplement. 2018. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 1 June 2021).
- Sader, H.S.; Jones, R.N.; Dowzicky, M.J.; Fritsche, T.R. Antimicrobial activity of tigecycline tested against nosocomial bacterial pathogens from patients hospitalized in the intensive care unit. Diagn. Microbiol. Infect. Dis. 2005, 52, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T.; Chiang, M.C.; Kuo, S.C.; Wang, Y.C.; Lee, I.H.; Chen, T.L.; Yang, Y.S. Carbapenem Breakpoints for Acinetobacter baumannii Group: Supporting Clinical Outcome Data from Patients with Bacteremia. PLoS ONE 2016, 11, e0163271. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, M.; Barbosa, P.I.; Vespero, E.C.; Tanita, M.T.; Carrilho, C.M.; Perugini, M.; Saridakis, H.O.; Possuelo, L.G.; Renner, J.D.; Valim, A.R.; et al. Carbapenem-resistant OXA-23-producing Acinetobacter baumannii isolates causing ventilator-associated pneumonia. Am. J. Infect. Control. 2010, 38, 667–669. [Google Scholar] [CrossRef] [PubMed]
- Espinal, P.; Macia, M.D.; Roca, I.; Gato, E.; Ruiz, E.; Fernandez-Cuenca, F.; Oliver, A.; Rodriguez-Bano, J.; Bou, G.; Tomas, M.; et al. First report of an OXA-23 carbapenemase-producing Acinetobacter baumannii clinical isolate related to Tn2006 in Spain. Antimicrob. Agents Chemother. 2013, 57, 589–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams-Haduch, J.M.; Paterson, D.L.; Sidjabat, H.E.; Pasculle, A.W.; Potoski, B.A.; Muto, C.A.; Harrison, L.H.; Doi, Y. Genetic basis of multidrug resistance in Acinetobacter baumannii clinical isolates at a tertiary medical center in Pennsylvania. Antimicrob. Agents Chemother. 2008, 52, 3837–3843. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.D.; Nickel, G.C.; Bajaksouzian, S.; Lavender, H.; Murthy, A.R.; Jacobs, M.R.; Bonomo, R.A. Resistance to colistin in Acinetobacter baumannii associated with mutations in the PmrAB two-component system. Antimicrob. Agents Chemother. 2009, 53, 3628–3634. [Google Scholar] [CrossRef] [Green Version]
- Oleksiuk, L.M.; Nguyen, M.H.; Press, E.G.; Updike, C.L.; O’Hara, J.A.; Doi, Y.; Clancy, C.J.; Shields, R.K. In vitro responses of Acinetobacter baumannii to two- and three-drug combinations following exposure to colistin and doripenem. Antimicrob. Agents Chemother. 2014, 58, 1195–1199. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Cuenca, F.; Martinez-Martinez, L.; Conejo, M.C.; Ayala, J.A.; Perea, E.J.; Pascual, A. Relationship between beta-lactamase production, outer membrane protein and penicillin-binding protein profiles on the activity of carbapenems against clinical isolates of Acinetobacter baumannii. J. Antimicrob. Chemother. 2003, 51, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Fu, Y.; Zhang, J.; Wang, Y.; Zhao, Y.; Fan, X.; Yu, L.; Wang, Y.; Zhang, X.; Li, C. Efficacy of tigecycline monotherapy versus combination therapy with other antimicrobials against carbapenem-resistant Acinetobacter baumannii sequence type 2 in Heilongjiang Province. Ann. Palliat. Med. 2019, 8, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Lertsrisatit, Y.; Santimaleeworagun, W.; Thunyaharn, S.; Traipattanakul, J. In vitro activity of colistin mono- and combination therapy against colistin-resistant Acinetobacter baumannii, mechanism of resistance, and clinical outcomes of patients infected with colistin-resistant A. baumannii at a Thai university hospital. Infect. Drug Resist. 2017, 10, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.; Daikos, G.L.; Durante-Mangoni, E.; Yahav, D.; Carmeli, Y.; Benattar, Y.D.; Skiada, A.; Andini, R.; Eliakim-Raz, N.; Nutman, A.; et al. Colistin alone versus colistin plus meropenem for treatment of severe infections caused by carbapenem-resistant Gram-negative bacteria: An open-label, randomised controlled trial. Lancet Infect. Dis. 2018, 18, 391–400. [Google Scholar] [CrossRef]
- Kaye, K. Trial for the Treatment of Extensively Drug-Resistant Gram-Negative Bacilli.October 2012–September 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT01597973 (accessed on 1 June 2021).
- Dizbay, M.; Tozlu, D.K.; Cirak, M.Y.; Isik, Y.; Ozdemir, K.; Arman, D. In vitro synergistic activity of tigecycline and colistin against XDR-Acinetobacter baumannii. J. Antibiot. 2010, 63, 51–53. [Google Scholar] [CrossRef]
- Satlin, M.J. The Search for a Practical Method for Colistin Susceptibility Testing: Have We Found It by Going Back to the Future? J. Clin. Microbiol. 2019, 57, e01163-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, P.A.; Petersen, P.J.; Young, M.; Jones, C.H.; Tischler, M.; O’Connell, J. Tigecycline MIC testing by broth dilution requires use of fresh medium or addition of the biocatalytic oxygen-reducing reagent oxyrase to standardize the test method. Antimicrob. Agents Chemother. 2005, 49, 3903–3909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markou, N.; Markantonis, S.L.; Dimitrakis, E.; Panidis, D.; Boutzouka, E.; Karatzas, S.; Rafailidis, P.; Apostolakos, H.; Baltopoulos, G. Colistin serum concentrations after intravenous administration in critically ill patients with serious multidrug-resistant, gram-negative bacilli infections: A prospective, open-label, uncontrolled study. Clin. Ther. 2008, 30, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Michalopoulos, A.S.; Falagas, M.E. Colistin: Recent data on pharmacodynamics properties and clinical efficacy in critically ill patients. Ann. Intensive Care 2011, 1, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouton, J.W.; van den Anker, J.N. Meropenem clinical pharmacokinetics. Clin. Pharmacokinet 1995, 28, 275–286. [Google Scholar] [CrossRef]
- Greer, N.D. Tigecycline (Tygacil): The first in the glycylcycline class of antibiotics. Proc. Bayl. Univ. Med. Cent. 2006, 19, 155–161. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antimicrobial | MIC Range (mg/L) | MIC50 (mg/L) | MIC90 (mg/L) | Median Fold-Reduction |
|---|---|---|---|---|
| COL | 0.5–256 | 4 | 8 | |
| MEM | 8–128 | 32 | 128 | |
| TGC | 0.25–8 | 4 | 4 | |
| MEM+COL | 1–64 | 16 | 32 | 2 |
| TGC+COL | 0.25–8 | 2 | 4 | 2 |
| Isolate Number | MLST | COL MIC (mg/L) | MEM MIC (mg/L) | Beta-Lactamase Genes | Sequencing Annotation | Differential Expression Analysis (Fold Increase/Decrease in Expression Compared to A. baumannii ATCC 17978) a | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PmrA | PmrB | ADC | OXA (Intrinsic) | OprD | CarO (porin) | OmpA Family | |||||
| R9314 | 3 | 1 | 16 | ADC-79, OXA-23, OXA-71 | S119T | P360Q, N440H | 12.13 | 3.65 | 0.459 | 0.025 | 0.154 |
| R9751 | 3 | 2 | 16 | ADC-79, OXA-23, OXA-71 | A14T, S119T | P360Q, N440H | 12.46 | 5.20 | 0.201 | 0.020 | 0.188 |
| R9761 | 2 | 0.5 | 16 | ADC-73, OXA-23, OXA-66, TEM-1 | WT | A138T, N440H, A444V | 4.35 | 2.35 | 0.283 | 0.594 | 0.090 |
| R11542 | 2 | 4 | 64 | ADC-33, OXA-23, OXA-82 | WT | N440H, A444V | 3.29 | 54.08 | 0.045 | 0.731 | 0.085 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul-Mutakabbir, J.C.; Yim, J.; Nguyen, L.; Maassen, P.T.; Stamper, K.; Shiekh, Z.; Kebriaei, R.; Shields, R.K.; Castanheira, M.; Kaye, K.S.; et al. In Vitro Synergy of Colistin in Combination with Meropenem or Tigecycline against Carbapenem-Resistant Acinetobacter baumannii. Antibiotics 2021, 10, 880. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070880
Abdul-Mutakabbir JC, Yim J, Nguyen L, Maassen PT, Stamper K, Shiekh Z, Kebriaei R, Shields RK, Castanheira M, Kaye KS, et al. In Vitro Synergy of Colistin in Combination with Meropenem or Tigecycline against Carbapenem-Resistant Acinetobacter baumannii. Antibiotics. 2021; 10(7):880. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070880
Chicago/Turabian StyleAbdul-Mutakabbir, Jacinda C., Juwon Yim, Logan Nguyen, Philip T. Maassen, Kyle Stamper, Zain Shiekh, Razieh Kebriaei, Ryan K. Shields, Mariana Castanheira, Keith S. Kaye, and et al. 2021. "In Vitro Synergy of Colistin in Combination with Meropenem or Tigecycline against Carbapenem-Resistant Acinetobacter baumannii" Antibiotics 10, no. 7: 880. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070880