
Diversity of International High-Risk Clones of Acinetobacter baumannii Revealed in a Russian Multidisciplinary Medical Center during 2017–2019

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Isolate Metadata and Typing
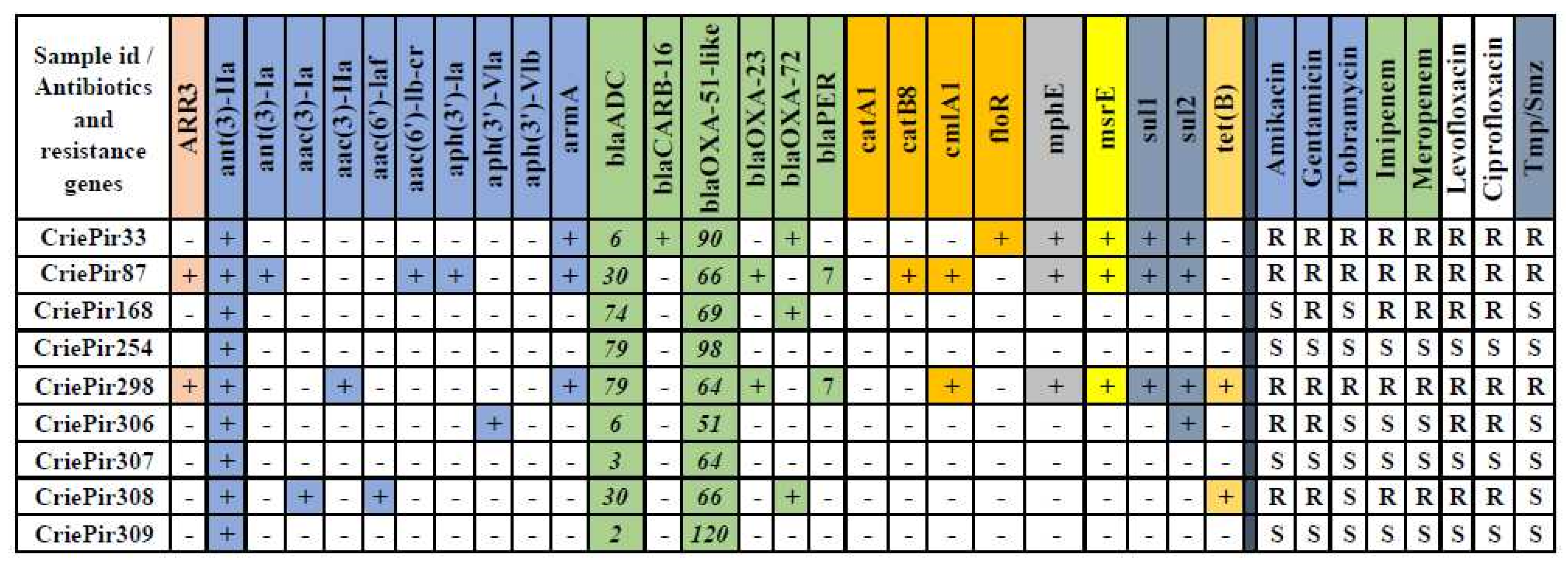
2.2. Antimicrobial Resistance
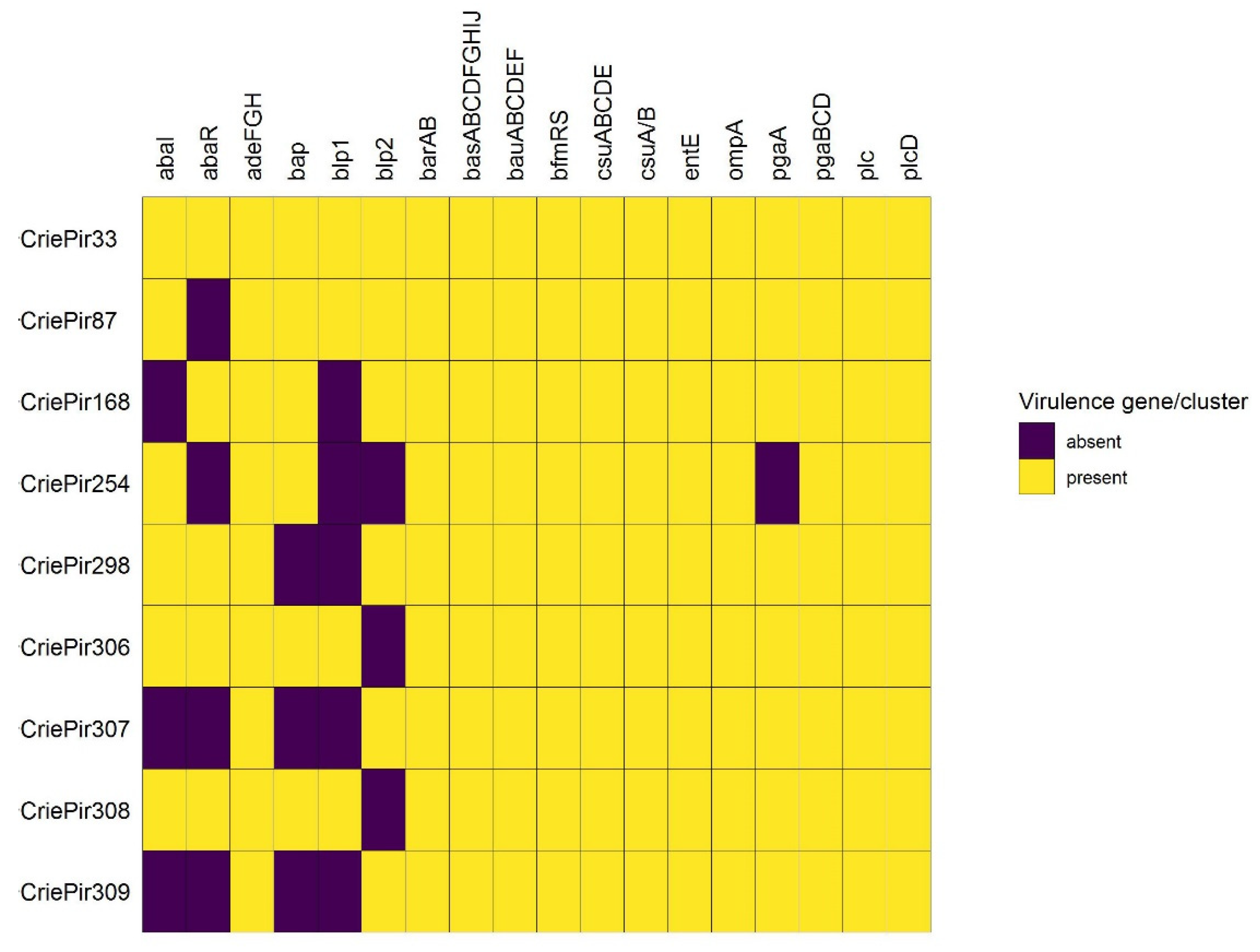
2.3. Virulence Genes
2.4. Plasmids
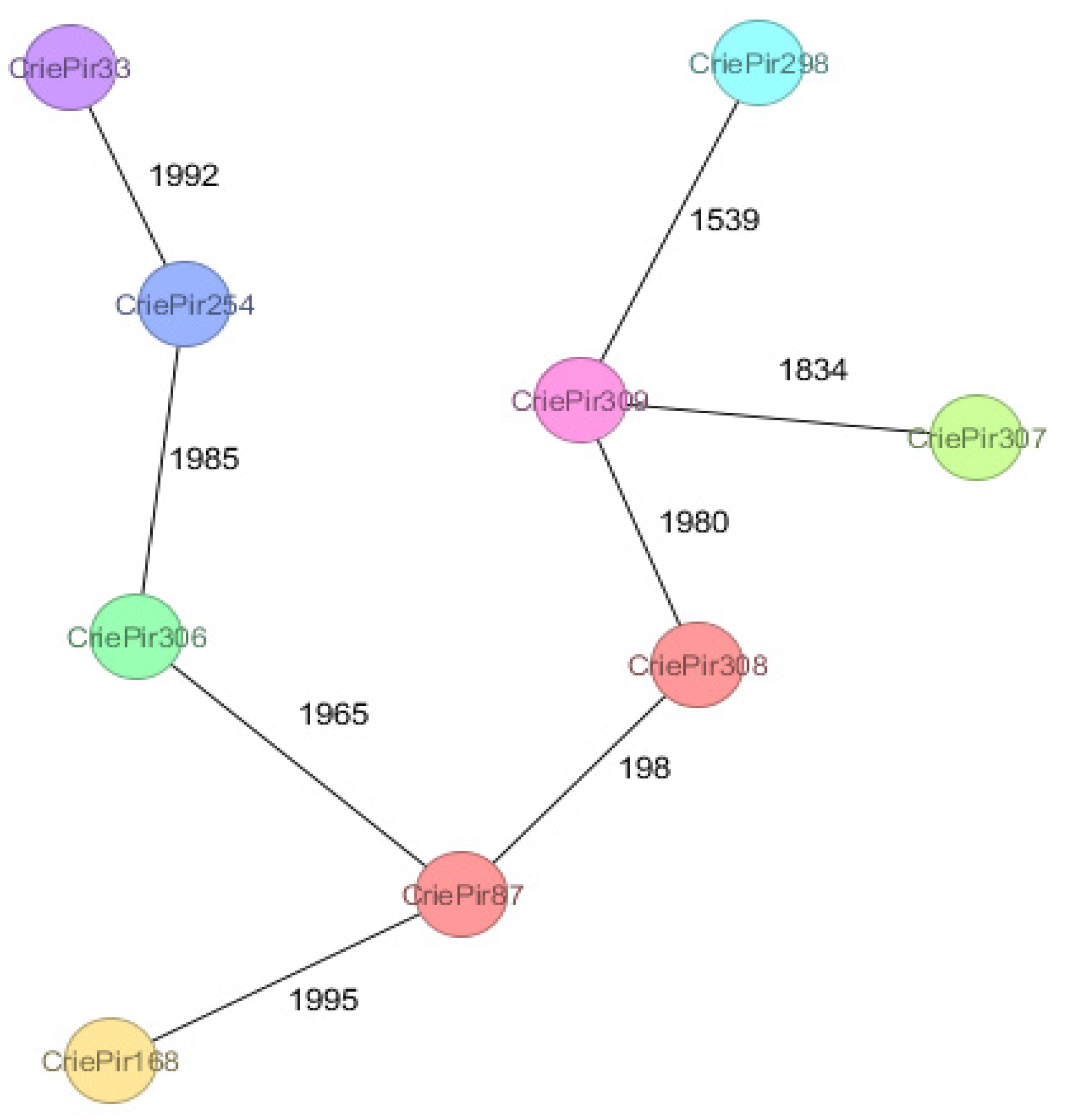
2.5. CRISPR Arrays and CRISPR/Cas Systems
3. Discussion
4. Materials and Methods
4.1. Determination of Antibiotic Susceptibility
4.2. DNA Isolation, Sequencing, and Genome Assembly
4.3. Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Armstrong, G.L.; MacCannell, D.R.; Taylor, J.; Carleton, H.A.; Neuhaus, E.B.; Bradbury, R.S.; Posey, J.E.; Gwinn, M. Pathogen Genomics in Public Health. N. Engl. J. Med. 2019, 381, 2569–2580. [Google Scholar] [CrossRef]
- Gilchrist, C.A.; Turner, S.D.; Riley, M.F.; Petri, W.A., Jr.; Hewlett, E.L. Whole-genome sequencing in outbreak analysis. Clin. Microbiol. Rev. 2015, 28, 541–563. [Google Scholar] [CrossRef] [Green Version]
- Maljkovic Berry, I.; Melendrez, M.C.; Bishop-Lilly, K.A.; Rutvisuttinunt, W.; Pollett, S.; Talundzic, E.; Morton, L.; Jarman, R.G. Next Generation Sequencing and Bioinformatics Methodologies for Infectious Disease Research and Public Health: Approaches, Applications, and Considerations for Development of Laboratory Capacity. J. Infect. Dis. 2020, 221, S292–S307. [Google Scholar] [CrossRef] [Green Version]
- Hawken, S.E.; Snitkin, E.S. Genomic epidemiology of multidrug-resistant Gram-negative organisms. Ann. N. Y. Acad. Sci. 2019, 1435, 39–56. [Google Scholar] [CrossRef]
- Palmieri, M.; D’Andrea, M.M.; Pelegrin, A.C.; Mirande, C.; Brkic, S.; Cirkovic, I.; Goossens, H.; Rossolini, G.M.; van Belkum, A. Genomic Epidemiology of Carbapenem- and Colistin-Resistant Klebsiella pneumoniae Isolates From Serbia: Predominance of ST101 Strains Carrying a Novel OXA-48 Plasmid. Front. Microbiol. 2020, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Osei Sekyere, J.; Reta, M.A. Genomic and Resistance Epidemiology of Gram-Negative Bacteria in Africa: A Systematic Review and Phylogenomic Analyses from a One Health Perspective. mSystems 2020, 5, e00897-20. [Google Scholar] [CrossRef] [PubMed]
- Talebi Bezmin Abadi, A.; Rizvanov, A.A.; Haertle, T.; Blatt, N.L. World Health Organization Report: Current Crisis of Antibiotic Resistance. BioNanoScience 2019, 9, 778–788. [Google Scholar] [CrossRef]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Medicin. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmenkov, A.Y.; Trushin, I.V.; Vinogradova, A.G.; Avramenko, A.A.; Sukhorukova, M.V.; Malhotra-Kumar, S.; Dekhnich, A.V.; Edelstein, M.V.; Kozlov, R.S. AMRmap: An Interactive Web Platform for Analysis of Antimicrobial Resistance Surveillance Data in Russia. Front. Microbiol. 2021, 12, 620002. [Google Scholar] [CrossRef]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate point-prevalence survey of health care-associated infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef] [Green Version]
- Lob, S.H.; Hoban, D.J.; Sahm, D.F.; Badal, R.E. Regional differences and trends in antimicrobial susceptibility of Acinetobacter baumannii. Int. J. Antimicrob. Agents 2016, 47, 317–323. [Google Scholar] [CrossRef]
- Zhen, X.; Lundborg, C.S.; Sun, X.; Hu, X.; Dong, H. Economic burden of antibiotic resistance in ESKAPE organisms: A systematic review. Antimicrob. Resist. Infect. Control. 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Hamidian, M.; Nigro, S.J. Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii. Microb. Genom. 2019, 5, e000306. [Google Scholar] [CrossRef]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Su, P.W.; Moi, S.H.; Chuang, L.Y. Biofilm Formation in Acinetobacter Baumannii: Genotype-Phenotype Correlation. Molecules 2019, 24, 1849. [Google Scholar] [CrossRef] [Green Version]
- Holt, K.; Kenyon, J.J.; Hamidian, M.; Schultz, M.B.; Pickard, D.J.; Dougan, G.; Hall, R. Five decades of genome evolution in the globally distributed, extensively antibiotic-resistant Acinetobacter baumannii global clone 1. Microb. Genom. 2016, 2, e000052. [Google Scholar] [CrossRef]
- Zarrilli, R.; Pournaras, S.; Giannouli, M.; Tsakris, A. Global evolution of multidrug-resistant Acinetobacter baumannii clonal lineages. Int. J. Antimicrob. Agents 2013, 41, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Al-Hassan, L.; Elbadawi, H.; Osman, E.; Ali, S.; Elhag, K.; Cantillon, D.; Wille, J.; Seifert, H.; Higgins, P.G. Molecular Epidemiology of Carbapenem-Resistant Acinetobacter baumannii From Khartoum State, Sudan. Front. Microbiol. 2021, 12, 628736. [Google Scholar] [CrossRef] [PubMed]
- Shek, E.A.; Sukhorukova, M.V.; Edelstein, M.V.; Skleenova, E.Y.; Ivanchik, N.V.; Shajdullina, E.R.; Kuzmenkov, A.Y.; Dekhnich, A.V.; Kozlov, R.S.; Semyonova, N.V.; et al. Antimicrobial resistance, carbapenemase production, and genotypes of nosocomial Acinetobacter spp. isolates in Russia: Results of multicenter epidemiological study “MARATHON 2015–2016”. Clin. Microbiol. Antimicrob. Chemother. 2019, 21, 171–180. [Google Scholar] [CrossRef]
- Mayanskiy, N.; Chebotar, I.; Alyabieva, N.; Kryzhanovskaya, O.; Savinova, T.; Turenok, A.; Bocharova, Y.; Lazareva, A.; Polikarpova, S.; Karaseva, O. Emergence of the Uncommon Clone ST944/ST78 Carrying blaOXA-40-like and blaCTX-M-like Genes Among Carbapenem-Nonsusceptible Acinetobacter baumannii in Moscow, Russia. Microb. Drug Resist. 2017, 23, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Nodari, C.S.; Cayo, R.; Streling, A.P.; Lei, F.; Wille, J.; Almeida, M.S.; de Paula, A.I.; Pignatari, A.C.C.; Seifert, H.; Higgins, P.G.; et al. Genomic Analysis of Carbapenem-Resistant Acinetobacter baumannii Isolates Belonging to Major Endemic Clones in South America. Front. Microbiol. 2020, 11, 584603. [Google Scholar] [CrossRef]
- Cerezales, M.; Xanthopoulou, K.; Wille, J.; Bustamante, Z.; Seifert, H.; Gallego, L.; Higgins, P.G. Acinetobacter baumannii analysis by core genome multi-locus sequence typing in two hospitals in Bolivia: Endemicity of international clone 7 isolates (CC25). Int. J. Antimicrob. Agents 2019, 53, 844–849. [Google Scholar] [CrossRef]
- Higgins, P.G.; Hagen, R.M.; Kreikemeyer, B.; Warnke, P.; Podbielski, A.; Frickmann, H.; Loderstadt, U. Molecular Epidemiology of Carbapenem-Resistant Acinetobacter baumannii Isolates from Northern Africa and the Middle East. Antibiotics 2021, 10, 291. [Google Scholar] [CrossRef]
- Shashkov, A.S.; Cahill, S.M.; Arbatsky, N.P.; Westacott, A.C.; Kasimova, A.A.; Shneider, M.M.; Popova, A.V.; Shagin, D.A.; Shelenkov, A.A.; Mikhailova, Y.V.; et al. Acinetobacter baumannii K116 capsular polysaccharide structure is a hybrid of the K14 and revised K37 structures. Carbohydr. Res. 2019, 484, 107774. [Google Scholar] [CrossRef]
- Bonnin, R.A.; Potron, A.; Poirel, L.; Lecuyer, H.; Neri, R.; Nordmann, P. PER-7, an extended-spectrum beta-lactamase with increased activity toward broad-spectrum cephalosporins in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 2424–2427. [Google Scholar] [CrossRef] [Green Version]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-lactamase database (BLDB)—structure and function. J. Enzyme Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef]
- Matthews, L.; Goodrich, J.S.; Weber, D.J.; Bergman, N.H.; Miller, M.B. The Brief Case: A Fatal Case of Necrotizing Fasciitis Due to Multidrug-Resistant Acinetobacter baumannii. J. Clin. Microbiol. 2019, 57, e01751-18. [Google Scholar] [CrossRef] [Green Version]
- Hamidian, M.; Hawkey, J.; Wick, R.; Holt, K.E.; Hall, R.M. Evolution of a clade of Acinetobacter baumannii global clone 1, lineage 1 via acquisition of carbapenem- and aminoglycoside-resistance genes and dispersion of ISAba1. Microb. Genom. 2019, 5, e000242. [Google Scholar] [CrossRef]
- Shelenkov, A.; Korotkov, A.; Korotkov, E. MMsat—a database of potential micro- and minisatellites. Gene 2008, 409, 53–60. [Google Scholar] [CrossRef]
- Yeh, H.Y.; Awad, A. Genotyping of Campylobacter jejuni Isolates from Poultry by Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR). Curr. Microbiol. 2020, 77, 1647–1652. [Google Scholar] [CrossRef]
- Bartual, S.G.; Seifert, H.; Hippler, C.; Luzon, M.A.; Wisplinghoff, H.; Rodriguez-Valera, F. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4382–4390. [Google Scholar] [CrossRef] [Green Version]
- Diancourt, L.; Passet, V.; Nemec, A.; Dijkshoorn, L.; Brisse, S. The population structure of Acinetobacter baumannii: Expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS ONE 2010, 5, e10034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelenkov, A.; Mikhaylova, Y.; Yanushevich, Y.; Samoilov, A.; Petrova, L.; Fomina, V.; Gusarov, V.; Zamyatin, M.; Shagin, D.; Akimkin, V. Molecular Typing, Characterization of Antimicrobial Resistance, Virulence Profiling and Analysis of Whole-Genome Sequence of Clinical Klebsiella pneumoniae Isolates. Antibiotics 2020, 9, 261. [Google Scholar] [CrossRef]
- Higgins, P.G.; Prior, K.; Harmsen, D.; Seifert, H. Development and evaluation of a core genome multilocus typing scheme for whole-genome sequence-based typing of Acinetobacter baumannii. PLoS ONE 2017, 12, e0179228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venditti, C.; Vulcano, A.; D’Arezzo, S.; Gruber, C.E.M.; Selleri, M.; Antonini, M.; Lanini, S.; Marani, A.; Puro, V.; Nisii, C.; et al. Epidemiological investigation of an Acinetobacter baumannii outbreak using core genome multilocus sequence typing. J. Glob. Antimicrob. Resist. 2019, 17, 245–249. [Google Scholar] [CrossRef]
- Evans, B.A.; Amyes, S.G. OXA beta-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [Green Version]
- Gaiarsa, S.; Batisti Biffignandi, G.; Esposito, E.P.; Castelli, M.; Jolley, K.A.; Brisse, S.; Sassera, D.; Zarrilli, R. Comparative Analysis of the Two Acinetobacter baumannii Multilocus Sequence Typing (MLST) Schemes. Front. Microbiol. 2019, 10, 930. [Google Scholar] [CrossRef] [Green Version]
- Cieslinski, J.M.; Arend, L.; Tuon, F.F.; Silva, E.P.; Ekermann, R.G.; Dalla-Costa, L.M.; Higgins, P.G.; Seifert, H.; Pilonetto, M. Molecular epidemiology characterization of OXA-23 carbapenemase-producing Acinetobacter baumannii isolated from 8 Brazilian hospitals using repetitive sequence-based PCR. Diagn. Microbiol. Infect. Dis. 2013, 77, 337–340. [Google Scholar] [CrossRef]
- Wareth, G.; Brandt, C.; Sprague, L.D.; Neubauer, H.; Pletz, M.W. Spatio-Temporal Distribution of Acinetobacter baumannii in Germany-A Comprehensive Systematic Review of Studies on Resistance Development in Humans (2000–2018). Microorganisms 2020, 8, 375. [Google Scholar] [CrossRef] [Green Version]
- Rafei, R.; Dabboussi, F.; Hamze, M.; Eveillard, M.; Lemarie, C.; Gaultier, M.P.; Mallat, H.; Moghnieh, R.; Husni-Samaha, R.; Joly-Guillou, M.L.; et al. Molecular analysis of Acinetobacter baumannii strains isolated in Lebanon using four different typing methods. PLoS ONE 2014, 9, e115969. [Google Scholar] [CrossRef] [Green Version]
- Caldart, R.V.; Fonseca, E.L.; Freitas, F.; Rocha, L.; Vicente, A.C. Acinetobacter baumannii infections in Amazon Region driven by extensively drug resistant international clones, 2016–2018. Mem. Inst. Oswaldo. Cruz. 2019, 114, e190232. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Sun, Q.; Ruan, Z.; Xie, X. Characterization of a small plasmid carrying the carbapenem resistance gene bla OXA-72 from community-acquired Acinetobacter baumannii sequence type 880 in China. Infect. Drug Resist. 2019, 12, 1545–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaras, A.P.; Dorsey, C.W.; Edelmann, R.E.; Actis, L.A. Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: Involvement of a novel chaperone-usher pili assembly system. Microbiology 2003, 149, 3473–3484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, A.H.; Slamti, L.; Avci, F.Y.; Pier, G.B.; Maira-Litran, T. The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-beta-1-6-N-acetylglucosamine, which is critical for biofilm formation. J. Bacteriol. 2009, 191, 5953–5963. [Google Scholar] [CrossRef] [Green Version]
- Sarshar, M.; Behzadi, P.; Scribano, D.; Palamara, A.T.; Ambrosi, C. Acinetobacter baumannii: An Ancient Commensal with Weapons of a Pathogen. Pathogens 2021, 10, 387. [Google Scholar] [CrossRef]
- Liu, C.; Chang, Y.; Xu, Y.; Luo, Y.; Wu, L.; Mei, Z.; Li, S.; Wang, R.; Jia, X. Distribution of virulence-associated genes and antimicrobial susceptibility in clinical Acinetobacter baumannii isolates. Oncotarget 2018, 9, 21663–21673. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Lu, F.; Yuan, F.; Jiang, D.; Zhao, P.; Zhu, J.; Cheng, H.; Cao, J.; Lu, G. Biofilm Formation Caused by Clinical Acinetobacter baumannii Isolates Is Associated with Overexpression of the AdeFGH Efflux Pump. Antimicrob. Agents Chemother. 2015, 59, 4817–4825. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Lopez, R.; Solano-Galvez, S.G.; Juarez Vignon-Whaley, J.J.; Abello Vaamonde, J.A.; Padro Alonzo, L.A.; Rivera Resendiz, A.; Muleiro Alvarez, M.; Vega Lopez, E.N.; Franyuti-Kelly, G.; Alvarez-Hernandez, D.A.; et al. Acinetobacter baumannii Resistance: A Real Challenge for Clinicians. Antibiotics 2020, 9, 205. [Google Scholar] [CrossRef]
- Tang, J.; Chen, Y.; Wang, X.; Ding, Y.; Sun, X.; Ni, Z. Contribution of the AbaI/AbaR Quorum Sensing System to Resistance and Virulence of Acinetobacter baumannii Clinical Strains. Infect. Drug Resist. 2020, 13, 4273–4281. [Google Scholar] [CrossRef]
- Schneidewind, L.; Kranz, J.; Tandogdu, Z. Rising significance of antibiotic stewardship in urology and urinary tract infections—a rapid review. Curr. Opin. Urol. 2021, 31, 285–290. [Google Scholar] [CrossRef]
- Provenzani, A.; Hospodar, A.R.; Meyer, A.L.; Leonardi Vinci, D.; Hwang, E.Y.; Butrus, C.M.; Polidori, P. Multidrug-resistant gram-negative organisms: A review of recently approved antibiotics and novel pipeline agents. Int. J. Clin. Pharm. 2020, 42, 1016–1025. [Google Scholar] [CrossRef]
- Shelenkov, A.; Slavokhotova, A.; Odintsova, T. Predicting Antimicrobial and Other Cysteine-Rich Peptides in 1267 Plant Transcriptomes. Antibiotics 2020, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, C.; Sarkar, P.; Issa, R.; Haldar, J. Alternatives to Conventional Antibiotics in the Era of Antimicrobial Resistance. Trends Microbiol. 2019, 27, 323–338. [Google Scholar] [CrossRef]
- Bebbington, C.; Yarranton, G. Antibodies for the treatment of bacterial infections: Current experience and future prospects. Curr. Opin. Biotechnol. 2008, 19, 613–619. [Google Scholar] [CrossRef]
- Uppu, D.S.; Manjunath, G.B.; Yarlagadda, V.; Kaviyil, J.E.; Ravikumar, R.; Paramanandham, K.; Shome, B.R.; Haldar, J. Membrane-active macromolecules resensitize NDM-1 gram-negative clinical isolates to tetracycline antibiotics. PLoS ONE 2015, 10, e0119422. [Google Scholar] [CrossRef] [Green Version]
- Tyumentseva, M.; Mikhaylova, Y.; Prelovskaya, A.; Tyumentsev, A.; Petrova, L.; Fomina, V.; Zamyatin, M.; Shelenkov, A.; Akimkin, V. Genomic and Phenotypic Analysis of Multidrug-Resistant Acinetobacter baumannii Clinical Isolates Carrying Different Types of CRISPR/Cas Systems. Pathogens 2021, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Arbatsky, N.P.; Shneider, M.M.; Dmitrenok, A.S.; Popova, A.V.; Shagin, D.A.; Shelenkov, A.A.; Mikhailova, Y.V.; Edelstein, M.V.; Knirel, Y.A. Structure and gene cluster of the K125 capsular polysaccharide from Acinetobacter baumannii MAR13-1452. Int. J. Biol. Macromol. 2018, 117, 1195–1199. [Google Scholar] [CrossRef]
- Wyres, K.L.; Cahill, S.M.; Holt, K.E.; Hall, R.M.; Kenyon, J.J. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 2020, 6, e000339. [Google Scholar] [CrossRef]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Neron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample id | Patient Code | Isolation Date | Clinical Department | Locus | MLST | OCL-Type | KL-Type | IC |
|---|---|---|---|---|---|---|---|---|
| CriePir33 | P1 | 03.05.2017 | Traumatology | Wound | ST78 | OCL1 | KL3 | IC6 (CC78) |
| CriePir87 | P2 | 04.07.2017 | Surgery | Soft tissue abscess | ST2 | OCL1 | KL33 | IC2 (CC2) |
| CriePir168 | P3 | 03.12.2017 | ICU | Urine | ST1 | OCL1 | KL17 | IC1 (CC1) |
| CriePir254 | P4 | 24.08.2018 | Surgery | Bile | ST370 | OCL1 | KL25 | CC252 |
| CriePir298 | P5 | 10.09.2019 | CNS Rehabilitation | Urine | ST25 | OCL6 | KL116 | IC7 (CC25) |
| CriePir306 | P6 | 05.08.2019 | ICU | BAL | ST15 | OCL7 | KL9 | IC4 (CC15) |
| CriePir307 | P7 | 29.06.2019 | ICU | BAL | ST1487 * | OCL2 | KL45 | CC152 |
| CriePir308 | P6 | 22.08.2019 | ICU | CVC | ST2 | OCL7 | KL9 | IC2 (CC2) |
| CriePir309 | P8 | 31.08.2019 | CNS Rehabilitation | Urine | ST911 | OCL6 | KL14 | CC132 |
| Sample Id | Number of Plasmids | Plasmid Sizes | Virulence/Resistance Determinants in Plasmids | Related Plasmids from Other Isolates |
|---|---|---|---|---|
| CriePir33 | 1 | 17,765 | blaOXA-72 | pIBAC_oxa58_20C15, KY202458.1 |
| CriePir87 | 1 | 11,194 | - | pUnnamed1, CP035673.1 |
| CriePir168 | 3 | 1217–10,878 | blaOXA-72 | pAB120, CP031446.1 |
| CriePir254 | 2 | 10,427–90,326 | - | pGFJ6, CP016902.1 |
| CriePir298 | 3 | 2343–183,139 | mph(E), msr(E), armA, sul1, sul2, blaPER-7, ARR-3, aph(6)-Id, aph(3′′)-Ib, aac(3)-IIa, tet(B) | pA85-1a, CP021784.1 |
| CriePir306 | 4 | 2845–80,829 | - | pA1296_1, CP018333.1 |
| CriePir307 | 6 | 2278–114,430 | - | pABAY15001_6E, MK386684.1 |
| CriePir308 | 1 | 10,879 | blaOXA-72 | pAB120, CP031446.1 |
| CriePir309 | 2 | 11,195–94,551 | - | pTS134338, CP042210.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shelenkov, A.; Petrova, L.; Zamyatin, M.; Mikhaylova, Y.; Akimkin, V. Diversity of International High-Risk Clones of Acinetobacter baumannii Revealed in a Russian Multidisciplinary Medical Center during 2017–2019. Antibiotics 2021, 10, 1009. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081009
Shelenkov A, Petrova L, Zamyatin M, Mikhaylova Y, Akimkin V. Diversity of International High-Risk Clones of Acinetobacter baumannii Revealed in a Russian Multidisciplinary Medical Center during 2017–2019. Antibiotics. 2021; 10(8):1009. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081009
Chicago/Turabian StyleShelenkov, Andrey, Lyudmila Petrova, Mikhail Zamyatin, Yulia Mikhaylova, and Vasiliy Akimkin. 2021. "Diversity of International High-Risk Clones of Acinetobacter baumannii Revealed in a Russian Multidisciplinary Medical Center during 2017–2019" Antibiotics 10, no. 8: 1009. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081009