
Thermal Shock and Ciprofloxacin Act Orthogonally on Pseudomonas aeruginosa Biofilms


1
Department of Chemical and Biochemical Engineering, 4133 Seamans Center for the Engineering Arts and Sciences, University of Iowa, Iowa City, IA 52242, USA
2
Department of Chemical Engineering, University of Technology, Baghdad 10066, Iraq
*
Author to whom correspondence should be addressed.
Antibiotics 2021, 10(8), 1017; https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081017
Submission received: 20 June 2021
/
Revised: 12 August 2021
/
Accepted: 16 August 2021
/
Published: 21 August 2021
(This article belongs to the Special Issue New Insights on Biofilm Antimicrobial Strategies, 2nd Volume)
Abstract
:Bacterial biofilm infections are a major liability of medical implants, due to their resistance to both antibiotics and host immune response. Thermal shock can kill established biofilms, and some evidence suggests antibiotics may enhance this efficacy, despite having an insufficient effect themselves. The nature of this interaction is unclear, however, complicating efforts to integrate thermal shock into implant infection treatment. This study aimed to determine whether these treatments were truly synergistic or simply orthogonal (i.e., independent). Pseudomonas aeruginosa biofilms of different architectures and stationary-phase population density were subjected to various thermal shocks, antibiotic exposures, or combinations thereof, and examined either immediately after treatment or after subsequent reincubation. Population decreases from the combination treatment matched the product of the decreases of individual treatments, indicating their orthogonality. However, reincubation showed binary behavior, where biofilms with an immediate population decrease beyond a critical factor (~104) died off completely during reincubation, while biofilms with a smaller immediate decrease regrew. This critical factor was independent of the initial population density and the combination of treatments that achieved the immediate decrease. While antibiotics do not appear to enhance thermal shock directly, their contribution to achieving a critical population decrease for biofilm elimination can make the treatments appear strongly synergistic, strongly decreasing the intensity of thermal shock needed.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
More than 750,000 knee replacement and 500,000 hip replacement surgeries are performed each year in the United States [1], and these numbers are expected to increase exponentially in the next decade [2]. In total, 1% to 4% of the knee replacement and 1% to 2% of the hip replacement procedures are followed by incidences of periprosthetic joint infection [3,4,5]. The pathogens in these infections typically organize themselves in a densely populated thin layer of polysaccharides, proteins, and DNA called a biofilm, in which they exhibit a phenotype 20–100 times more resistant to antibiotics and host immune response than their planktonic phenotype [6,7,8,9,10]. Thus, the current standard of care is high doses of antibiotics and surgical explantation of the implant with its surrounding infected tissue, followed eventually by implantation of a replacement device [11,12,13]. Though this is successful in over 90% of cases [14,15], the new implant has a higher risk of infection than the original one [16]. These multiple invasive procedures expose the patient to physical risk and low quality of life, in addition to significant financial costs [11,17]. The incidence of infection has persisted despite decades of effort to create surfaces that prevent biofilm formation [18,19,20,21,22,23,24] and to develop methods to eradicate established biofilms [25,26,27,28,29,30,31,32], none of which have progressed to clinical implementation.
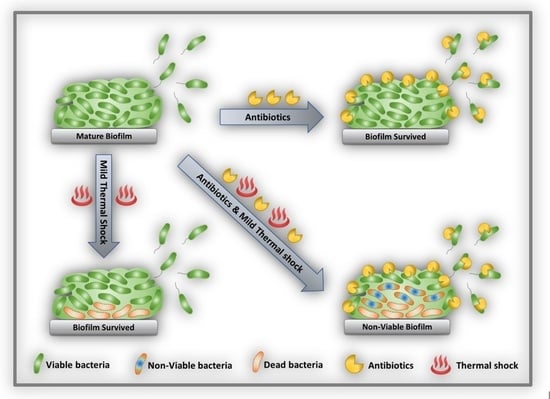
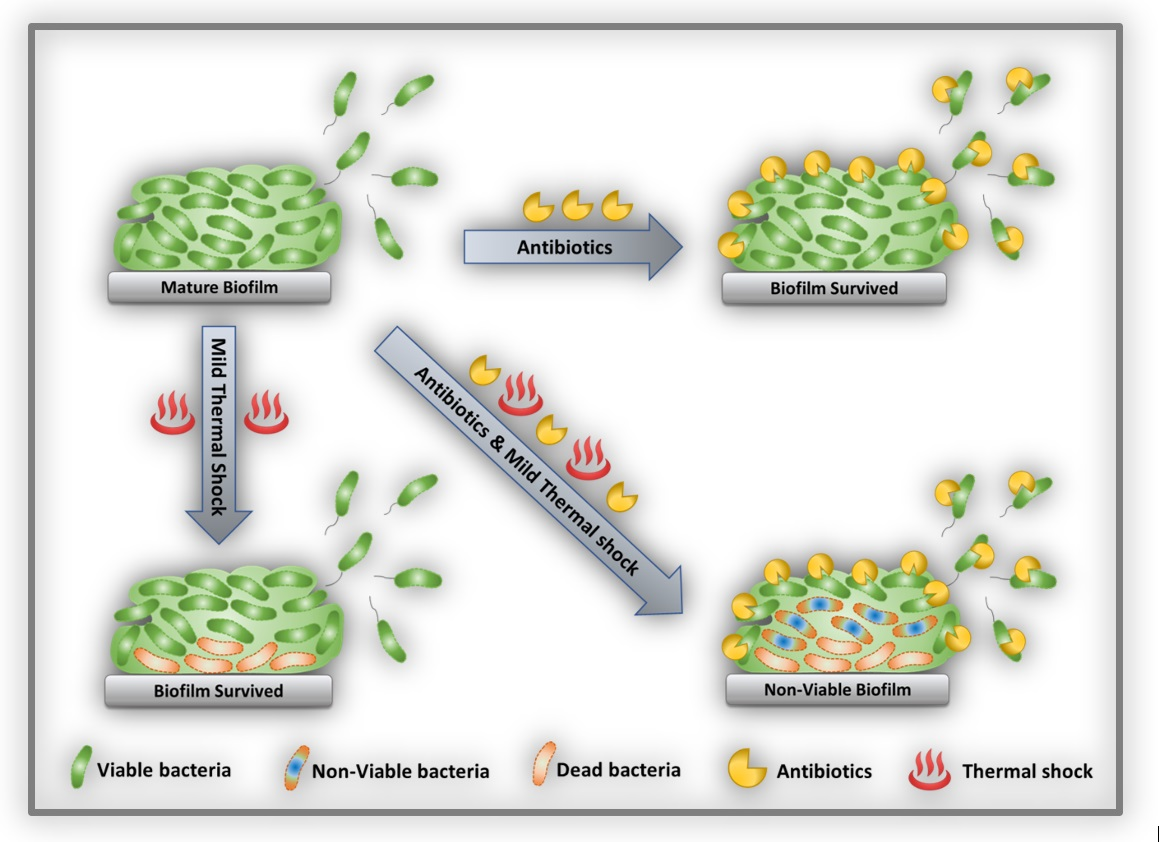
Thermal shock has been demonstrated as a means of deactivating established biofilms but may also damage adjacent tissue [33,34]. Recent studies have suggested a synergism between antibiotics and thermal shock, with the combined treatment decreasing biofilm population density more than either treatment alone, or even by the product of their individual effects [35,36,37,38]. The nature of this interaction is poorly understood. One hypothesis is that biofilms may have a critical population density below which they become non-viable, and that any combination of approaches that drops the population below that level will result in complete elimination of the biofilm, even if the individual approaches are not nearly so effective. This study investigated this hypothesis in Pseudomonas aeruginosa biofilms. Using biofilms of significantly different architecture and initial population density, it demonstrates not a critical population level but rather a critical population decrease, beyond which the population proceeds to zero.
Two different protocols were used to culture P. aeruginosa biofilms with significantly different population density and architecture: Shaker table (ST) and drip flow reactor (DFR). Biofilms of each type were subjected to thermal shocks ranging from 50 °C for 1 min to 70 °C for 30 min, with or without 4 h of prior exposure to ciprofloxacin (CP) concentrations ranging from 0.25 to 64 μg/mL. Shocked biofilms were either immediately enumerated or reincubated for 1 or 2 days before enumeration. Biofilms of each type were similarly exposed to antibiotics and re-incubation without thermal shock.
2. Results
2.1. Population Density, Architecture, and Thermal Susceptibility
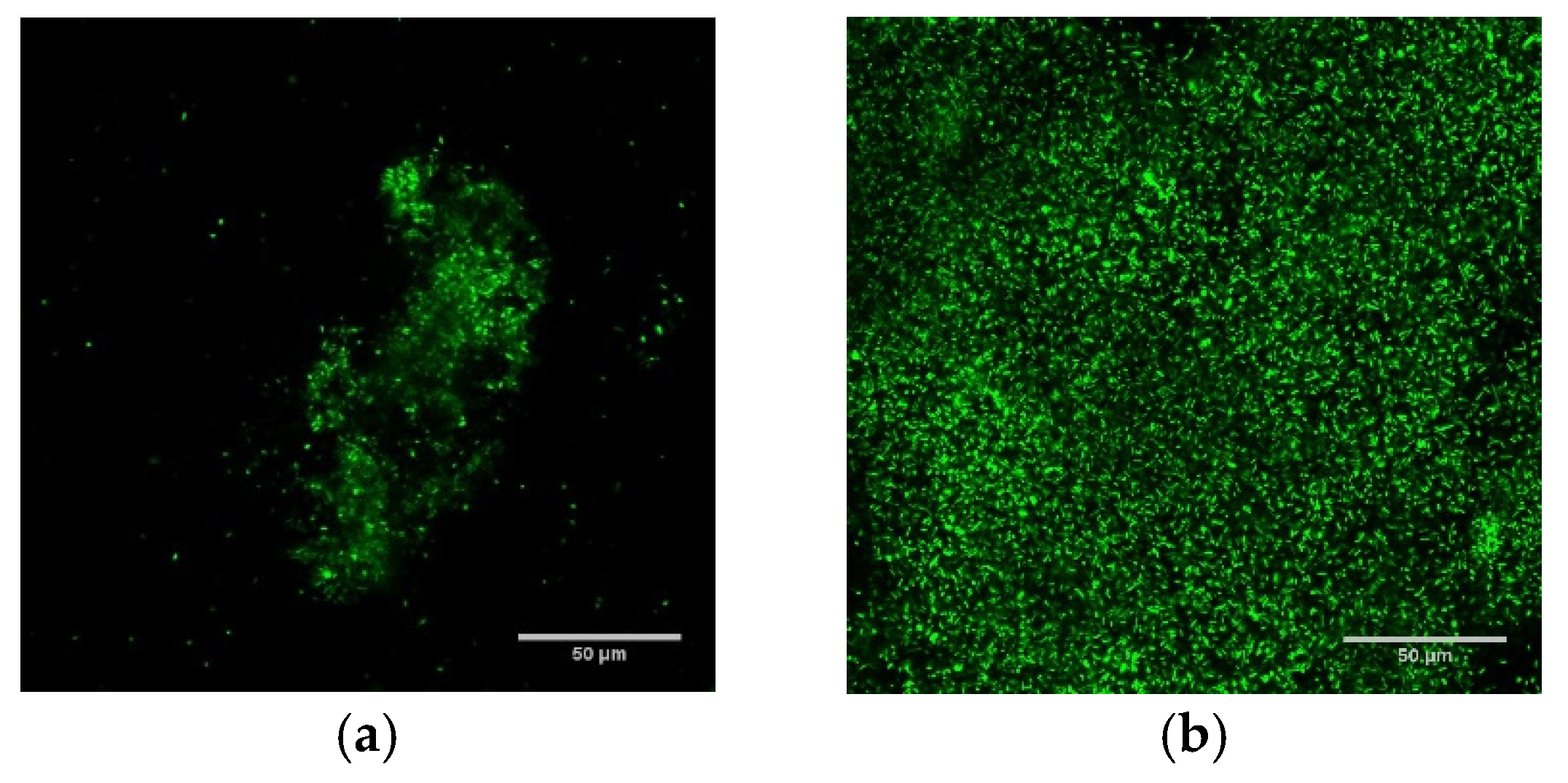
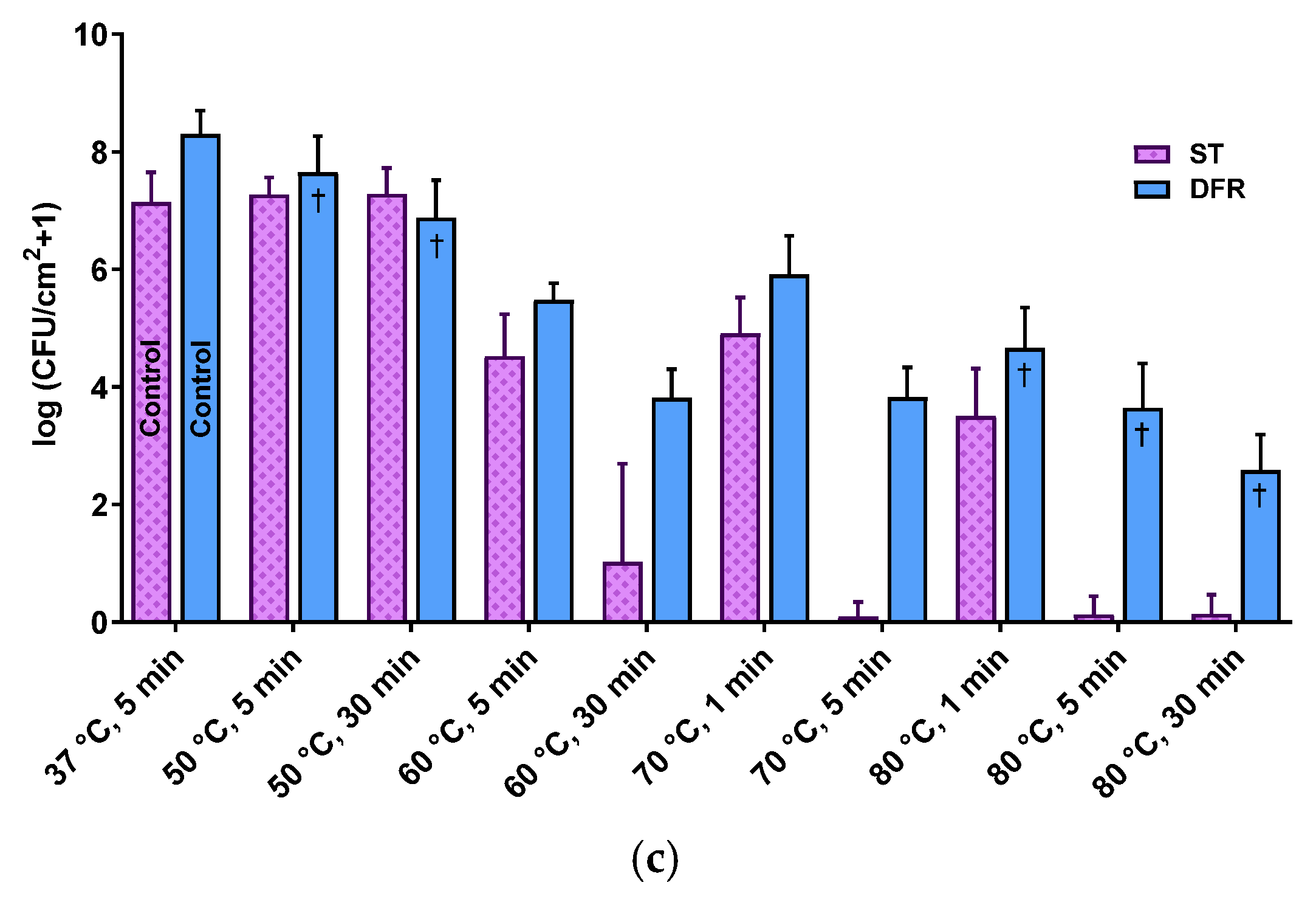
Biofilms grown using DFR and ST protocols had markedly different population densities (as measured in colony forming units (CFU) per cm2), spanning from sparsely populated (107.13±0.58 CFU/cm2) biofilms with individual micron-scale features ~50 μm in height (ST) to densely populated (108.3±0.4 CFU/cm2) carpets over 100 μm thick (DFR). Figure 1 demonstrates these architectural differences with confocal fluorescent microscopy images of ST (panel a) and DFR (panel b) biofilms, as well as comparing their population densities (panel c).
The thermal susceptibility of the biofilms was similarly different at modest temperatures (50 °C), with DFR biofilms decreasing by up to 1.5 orders of magnitude while ST biofilms showed no effect. At high temperatures (80 °C), however, the thermal susceptibility of the different biofilms converges, with both biofilm types decreasing by 3.5 orders of magnitude after 1 min of exposure. For longer exposures, ST biofilm populations appear to drop off completely.
2.2. Re-Incubation
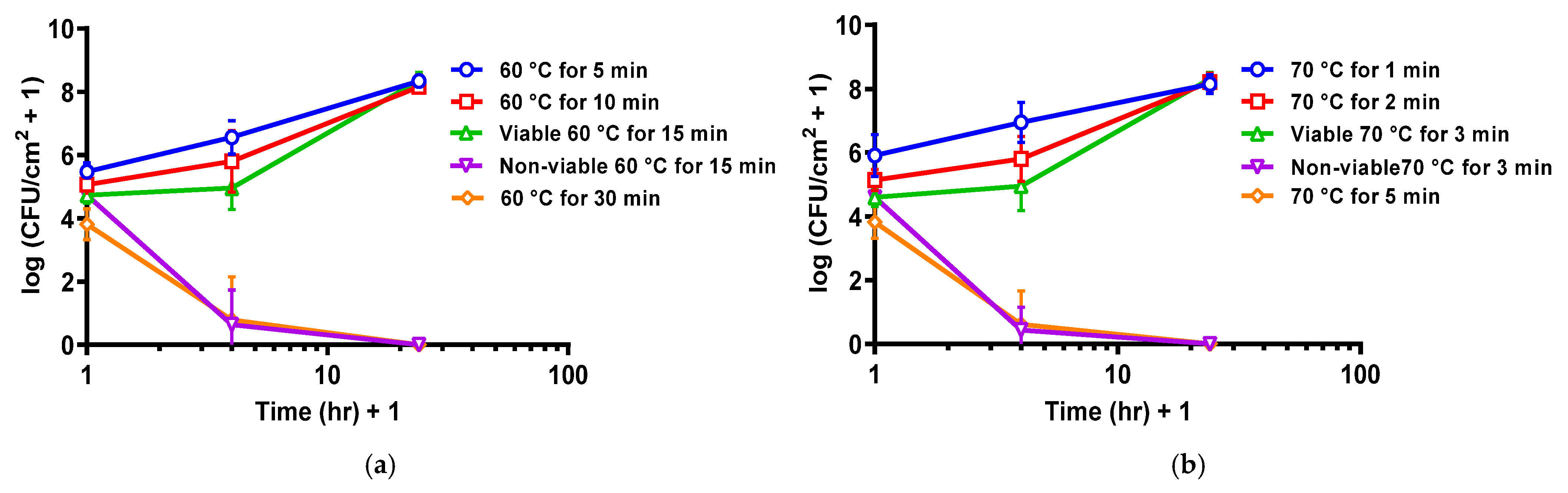
Re-incubation of thermally shocked DFR biofilms showed two opposing trends. After shocks sufficient to drop the population density below 104.5 CFU/cm2, the population density continued to decrease during re-incubation, dying off completely within a few hours. After milder thermal shocks, however, the biofilms regrew during reincubation, achieving their previous stationary-phase population density within a day. This is shown graphically in Figure 2 for eight thermal shock conditions, two of which typically dropped DFR biofilms to ~104.5 CFU/cm2. At those two conditions, both trends can be observed, with 60% (6 of 10) of the biofilms shocked at 60 °C for 15 min dying off while the remainder grew back (Figure 2a), and 66% (8 of 12) of the biofilms shocked at 70 °C for 3 min dying off while the remainder grew back (Figure 2b).
2.3. Antibiotic Exposure
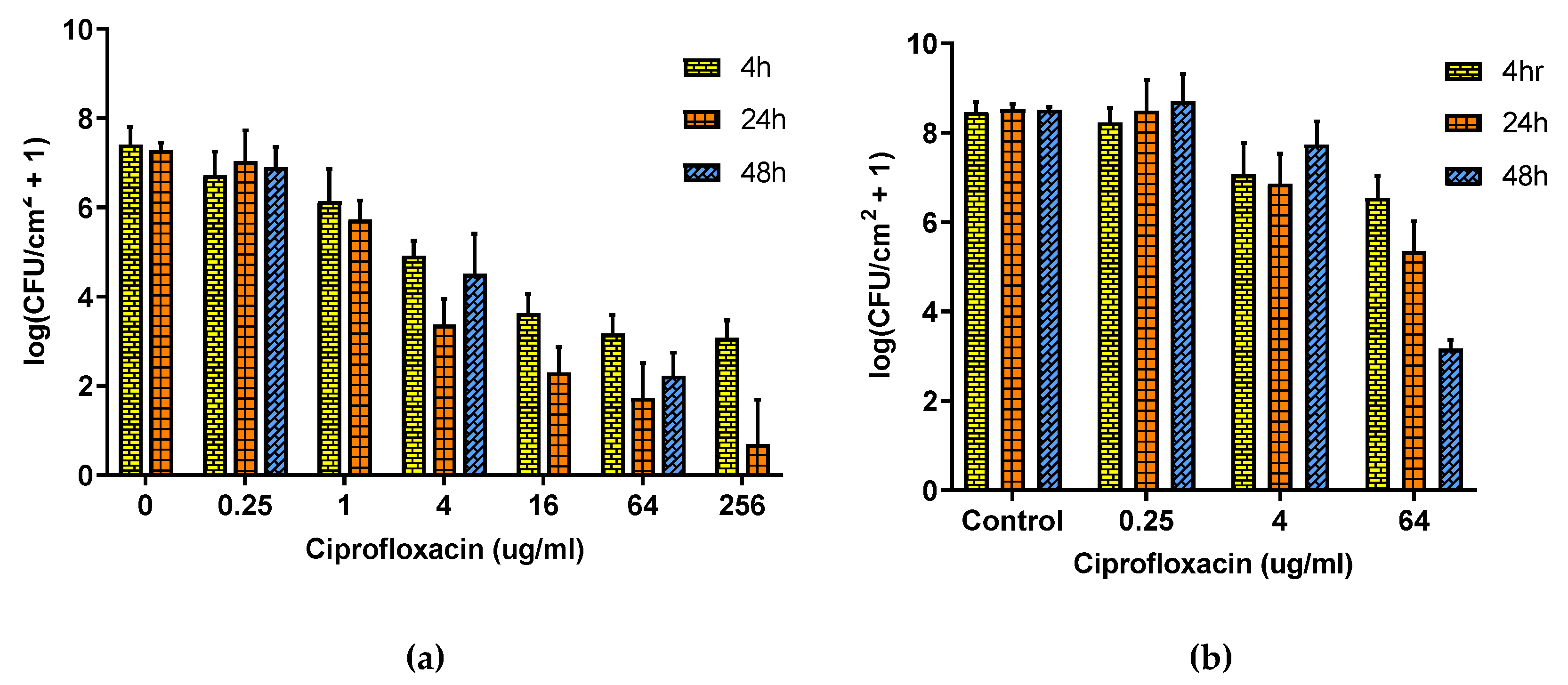
Figure 3 shows that the relationship between the P. aeruginosa population density and CP concentration follows a power law for both DFR and ST biofilms, albeit shifted, with the same dosage having less efficacy on DFR biofilms than on ST ones. Panel (a) shows that for ST biofilms, four hours of exposure failed to decrease the population density by four orders of magnitude even at concentrations over 60 times higher than physiological dosing 4 μg/mL [39]. Increasing the exposure time to 24 h increased efficacy but still did not eliminate the biofilm, and with further exposure (48 h) the population density actually recovered rather than further decreased. The effect of CP is much smaller on DFR biofilms (Panel b) with only modest decreases in population density even at grossly toxic concentration (64 μg/mL) and long exposure times, though under these circumstances, the population density is at least still trending downward with exposure time.
2.4. Combined Antibiotic and Thermal Exposure of Shaker Table (ST) Biofilms
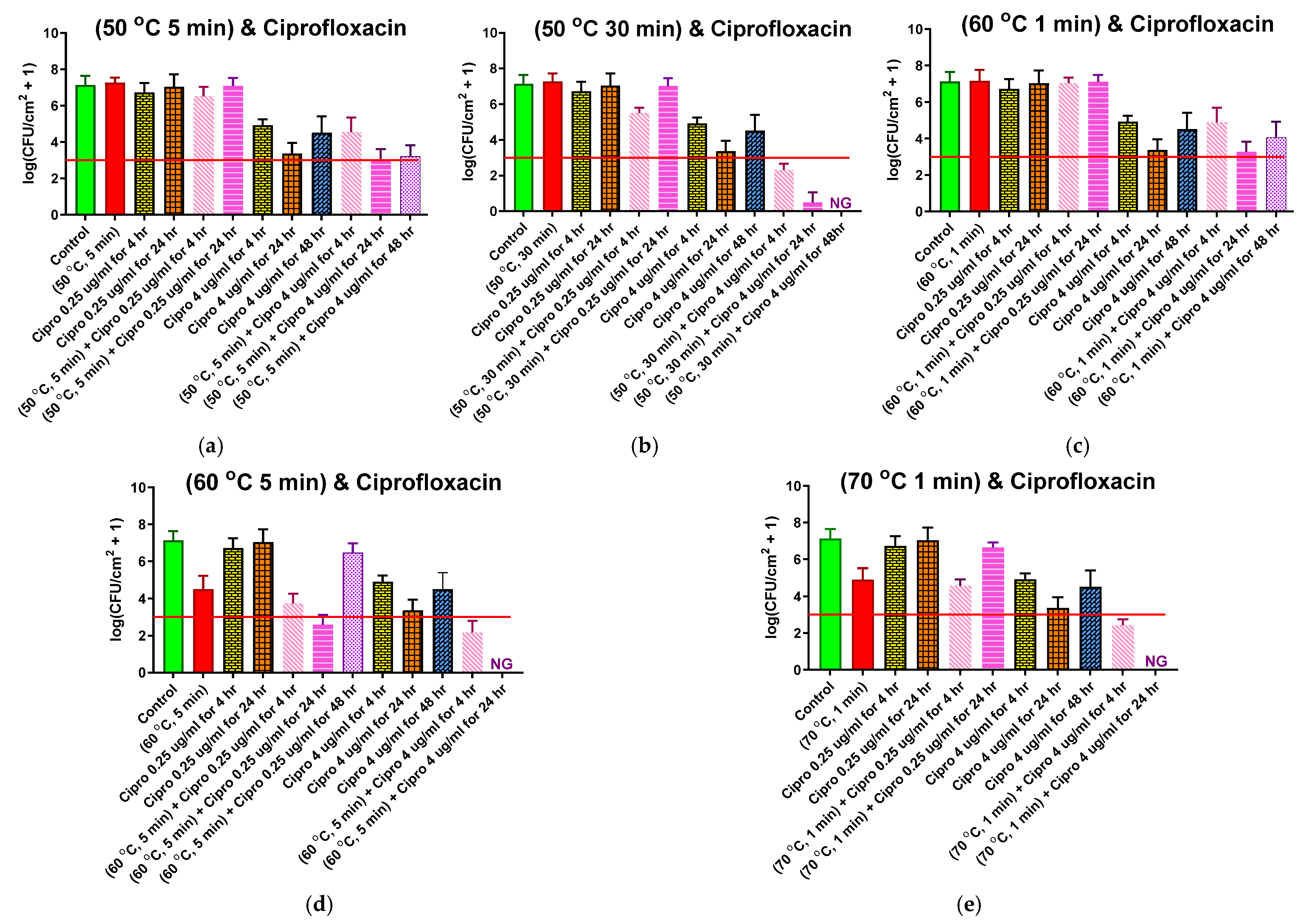
Five different thermal shock protocols (50 °C for 5 or 30 min, 60 °C for 1 or 5 min, 70 °C for 1 min) were applied to ST biofilms after 4 h of CP (0.25 or 4.0 μg/mL) exposure at 37 °C. For each protocol and CP concentration, three reincubation conditions were investigated: 0, 24, or 48 h in fresh media at the designated CP concentration. The results for each protocol are shown in their own panel of Figure 4, alongside controls with no thermal shock. These controls include: No treatment (first, or leftmost, bar of each panel), with an average pre-treatment population density of 107.13 CFU/cm2; 0.25 μg/mL CP exposure for 4 or 24 h (3rd and 4th bars, respectively), showing that this exposure by itself had no significant effect on population density; and 4 μg/mL CP exposure for 4, 24, or 48 h (7th, 8th, and 9th bars, respectively), showing that, by itself, this maximum physiological dose of ciprofloxacin only reduced the population density to 104.92 ±0.33 CFU/cm2 after 4 h of exposure and 103.37±0.58 after 24 h of exposure, recovering back to 104.51±0.9 after 48 h of exposure.
The second bar of each chart shows the effect of the thermal shock by itself (i.e., no antibiotic exposure), with only the shocks at 60 °C for 5 min (Figure 4d) or 70 °C for 1 min (Figure 4e) showing any significant effect. Combining thermal shock with exposure to 0.25 μg/mL CP, thermal shocks that had no effect by themselves also had no effect when added with 0.25 μg/mL antibiotic. There is a significant effect for the 50 °C for 30 min (Figure 4b) shock after 4 h of antibiotic exposure, but this disappeared with 24 h of exposure and may be an anomaly. For thermal shocks that did have a significant effect by themselves, 4 h of exposure to 0.25 μg/mL CP may have increased the effect, albeit not significantly. Further exposure for 24 h has conflicting results, with the enhancement becoming significant with a 60 °C shock for 5 min (Figure 4d), while allowing regrowth after 70 °C shocks for 1 min (Figure 4e). No consistent synergism (or antagonism) is seen.
Combining thermal shock with exposure to 4.0 μg/mL CP, shocks at 50 °C for 5 min (Figure 4a) and 60 °C for 1 min (Figure 4c) appear to have no effect; the results are essentially the same with or without the shock. Since these shocks also had no effect by themselves, one can state that the effects of the two treatments are additive, with the log reduction of the combined treatment matching the sum of the log reductions for each treatment by itself. Similarly, for more aggressive shocks (60 °C for 5 min, or 70 °C for 1 min), both the thermal shock and the 4 h exposure to 4 μg/mL CP have an effect by themselves, and the log reduction in population density for the combined treatment roughly matches the sum of the log reductions for each individual treatment. Only for the intermediate shock at 50 °C for 30 min (Figure 5b) does the combined treatment prompt a log reduction larger than the sum of the individual treatments, suggesting a synergism between the treatments.
Additional exposure to 4 μg/mL CP after these more aggressive thermal shocks has a dramatically different result, however. After 24 h of exposure, the biofilms shocked at 60 °C for 5 min or 70 °C for 1 min have no detectable CFU, see Figure 4d,e respectively. After 48 h, the biofilm shocked at 50 °C for 30 min (Figure 4b) also has no detectable CFU. These treatments appear highly synergistic.
2.5. Combined Antibiotic and Thermal Exposure of Drip Flow Reactor (DFR) Biofilms
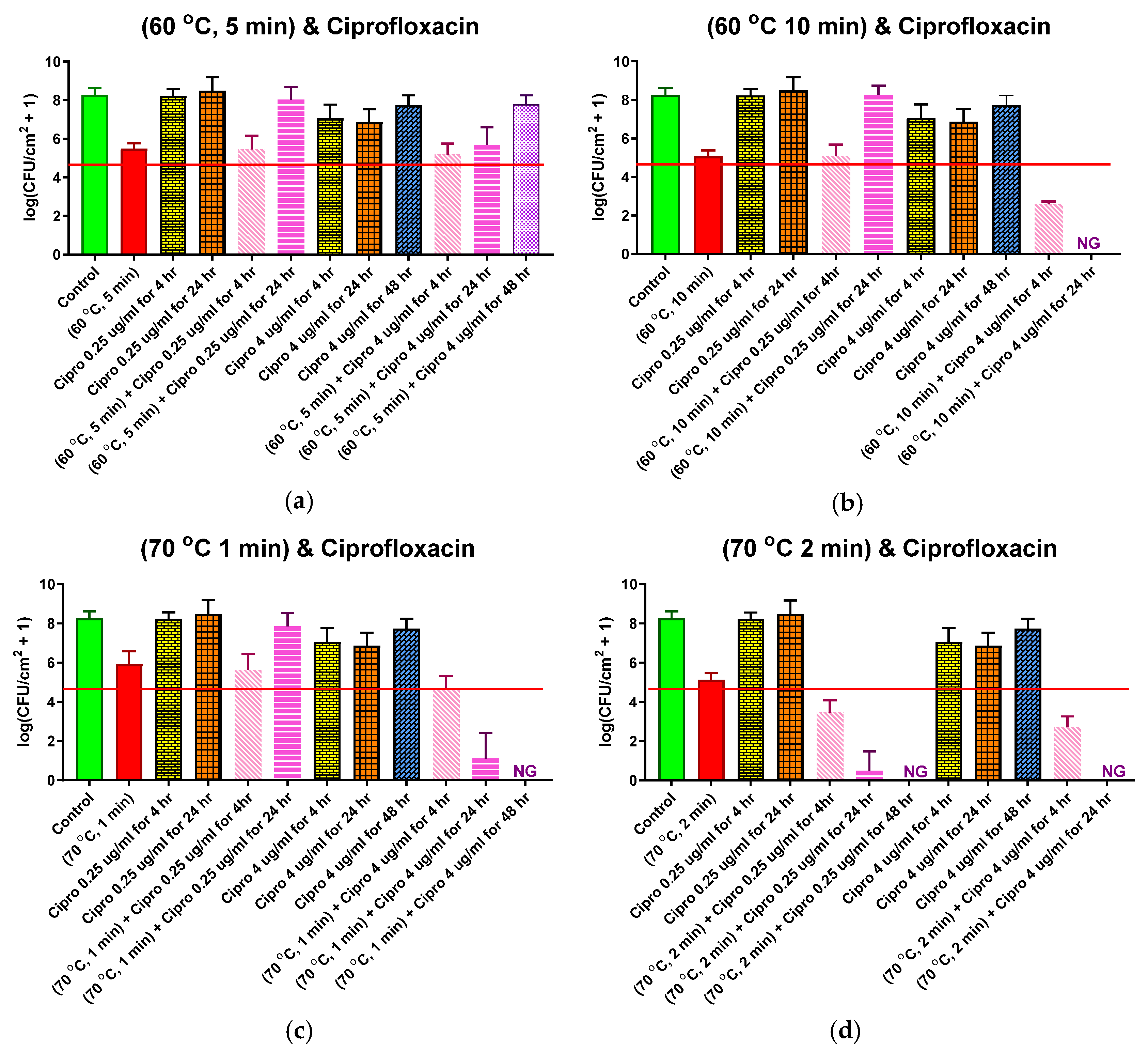
Four different thermal shock protocols (60 °C for 5 or 10 min, 70 °C for 1 or 2 min) were applied to DFR biofilms in combination with subsequent CP (0.25 or 4.0 μg/mL) exposure at 37 °C for 4, 24, or 48 h. These protocols were chosen because they each cause a large population density reduction in DFR biofilms but not so large that the biofilm eventually dies off, as seen in Figure 2. The results for each protocol are shown in their own panel of Figure 5, alongside controls with no thermal shock. These controls include: No treatment (first, or leftmost, bar of each panel), with an average pre-treatment population density of 108.3 CFU/cm2; 0.25 μg/mL CP exposure for 4 or 24 h (3rd and 4th bars, respectively), showing that this exposure by itself had no significant effect on population density; and 4 μg/mL CP exposure for 4, 24, or 48 h (7th, 8th, and 9th bars, respectively), showing that, by itself, this maximum physiological dose of CP only reduced the population density by an average log reduction of about 1.5 after 4 or 24 h of exposure, and that biofilms actually recovered to approximately their pre-treatment population density within 48 h, despite continuous antibiotics exposure.
The second bar of each chart shows the effect of the thermal shock by itself (i.e., no antibiotic exposure), with log reductions of 2.3–3.4. Adding four hours of exposure to 0.25 μg/mL CP, the thermal shock had virtually no additional effect except at the highest temperature for its longest shock (70 °C for 2 min), where an additional log reduction of 1.7, to 103.5 CFU/mL, was observed (Figure 5d). Increasing the antibiotic exposure to 24 h simply allowed the biofilm to grow back to its pre-treatment population density, except in the latter case, where the bacteria continued to die off, with no detectable CFU after 48 h.
Adding four hours of exposure to 4.0 μg/mL CP after the thermal shock, however, reduced the population density beyond the thermal shock alone in each case. For the shorter shocks at each temperature (Figure 5a,c), the log reductions of the combined treatments were slightly less than the sum of the log reductions of each treatment alone, see Figure 5b,d. The longer shocks, which had larger log reductions by themselves, also had larger log reductions from the combined treatment, even larger than the sum of the individual treatments. With longer antibiotic exposure, the biofilm population plummeted for most cases, with no detectable CFU after 24 h for the longer shocks at each temperature, and no detectable CFU after 48 h for the 1 min shock at 70 °C. Only for the 5 min shock at 60 C°, which had the highest population density after 4 h of 4 μg/mL CP exposure, did the biofilm regrow with continued antibiotic exposure, eventually (48 h) approaching its pre-treatment population density. Like the ST biofilms, these results suggest significant synergism between thermal shock and antibiotic exposure.
3. Discussion
Biofilm infections are particularly problematic because they cannot be eliminated by our primary in vivo treatment against bacteria, antibiotics. Ciprofloxacin’s ability to eliminate this strain of bacteria in the planktonic phenotype has been previously reported [35]. In the biofilm phenotype, however, dramatically higher drug concentrations are needed for comparable population reduction. While this reduction follows a power-law relationship with concentration, it levels out without elimination of the biofilm, as reported previously for MBEC assay biofilms [35] and indicated here in Figure 3 for ST biofilms, where at grossly toxic antibiotic concentrations, the population density appears to asymptote at ~103 CFU/cm2 after 4 h. The population density decreases further over the next 20 h but recovers again over the following day. For DFR biofilms, the decrease is irrelevant even at grossly toxic concentrations. Other approaches are clearly needed for biofilm infection mitigation.
Thermal shock is known to kill biofilms, but implementation in vivo requires minimizing the severity of the shock in order to minimize the accompanying damage to the adjacent tissue. Previous reports have indicated that antibiotics increase the efficacy of thermal shock, mitigating biofilms with less aggressive thermal treatment [35]. In some cases, the two approaches appear to be synergistic, with the reduction from the combined treatment exceeding the sum of the reductions from the individual treatments. The nature of this synergism was unknown, so this project aimed to investigate it, specifically scrutinizing the effect of re-incubation, with or without antibiotics, for biofilms spanning a wide range of architecture and population density.
As discussed earlier, the ST and DFR growth protocols involve starkly different growing conditions, which provide biofilms with dramatically different population density, structure, and maturity, as illustrated in Figure 1. These differences also prompt different thermal susceptibilities, with DFR biofilms strongly impacted by 50 °C exposure while ST biofilms are unaffected by it. At higher temperatures, however, their susceptibility (as measured by immediate population decrease) converges.
Immediate population decrease does not appear to completely quantify thermal susceptibility, though. After the thermal shock is removed and incubation conditions are restored, some DFR biofilms will recover to their pre-treatment population density, while others will continue to die off until no CFU are detectable. Figure 2 shows that this behavior can be predicted from the population density immediately after the thermal shock. DFR biofilms with a population density above ~104.5 CFU/cm2 grow back, while biofilms with a population density below that value die off, regardless of the temperature of the shock. Similar behavior was previously reported for ST biofilms, but in that case, the critical population density was ~103 CFU/cm2 [40]. Notably, this difference in critical population density (by a factor of ~101.5) is the same as the difference in the initial population densities of the two biofilms. In both cases, when the population density dropped by over four orders of magnitude, the biofilm died off, while smaller population density reductions resulted in the biofilm growing back completely, no matter what temperature and exposure time were used to achieve the reduction. These results suggest that mitigating biofilms is not based on driving its population density below a particular critical quorum, but rather achieving a particular population density reduction. This is encouraging since, at higher temperatures, this reduction appears to be the same regardless of the type of biofilm or its initial population density, as shown in Figure 1.
Figure 4 and Figure 5 suggest that this re-incubation behavior is responsible for the perceived synergism between thermal shock and antibiotics. Each panel in Figure 5 includes a horizontal line at the critical population density identified in Figure 2. Similarly, each panel in Figure 4 includes a horizontal line at the critical population density previously reported for ST biofilms [40]. Looking at combination treatments that resulted in populations above the critical value after 4 h, the decrease in population density for the combined thermal shock + antibiotics treatment is not significantly different than the sum of the decreases by thermal shock alone and antibiotics alone. The only exception to this in either figure is for ST biofilms exposed to 0.25 μg/mL ciprofloxacin and shocked at 50 °C for 30 min. In every other case, the treatments appear to be simply additive, indicating that their mechanisms are orthogonal and non-overlapping but not synergistic. Even for the 50 °C/30 min/0.25 μg/mL case, the enhanced decrease disappears within 24 h.
On the other hand, when the population reduction of the combined thermal shock + antibiotic treatment brings the population density below the critical value, this reduction is significantly larger than the sum of the reductions of its individual components, and this difference becomes pronounced at longer times as the population of CFU drops to zero. This is demonstrated five times in Figure 4 (50 °C for 30 min with 4 μg/mL; 60 °C for 5 min with 0.25 μg/mL) and Figure 5 (60 °C for 10 min with 4 μg/mL; 70 °C for 1 min with 4 μg/mL; 70 °C for 2 min with 0.25 μg/mL) with a wide range of temperature/time/antibiotic combinations, showing that this is independent of the original architecture of the biofilm and the manner in which the critical decrease is achieved. There are three instances where the reduction at 4 h is beyond the critical reduction but not significantly different than the sum of component reductions (ST biofilms shocked at 60 °C for 5 min or 70 °C for 1 min and DFR biofilms shocked at 70 °C for 2 min, all exposed to 4 μg/mL ciprofloxacin), but in all three cases, the biofilms proceed to die off within 24 h, resulting in a population reduction that is again much larger than the sum of the heat-shock only and antibiotics-only reductions. While neither treatment by itself achieved the critical population drop, the combination of the treatments did, prompting further population decrease, which makes the treatment appear synergistic, even though there is no indication that the treatments actually interact in any way.
In summary, previous studies have suggested a synergistic link between antibiotic exposure and thermal shock in the eradication of bacterial biofilms, i.e., that one treatment enhances the efficacy of the other in some way, resulting in a population reduction larger than predicted from simply adding the effects of the treatments by themselves. This study investigated that link, growing P aeruginosa biofilms of two disparate population densities and architectures and combining a variety of different thermal shocks with different concentrations of ciprofloxacin. When the sum of the log population decrease for thermal shock alone and for antibiotics alone was less than four orders of magnitude, the population decrease when combining the two treatments was not significantly different than the sum from the individual treatments, indicating no synergism, just orthogonal mechanisms. However, when the sum from the individual treatments was more than four orders of magnitude, the decrease from the combined treatment was even larger, eventually eliminating the biofilm altogether. While this gives the appearance of synergistic interaction in just those instances, the same eventual elimination is observed even without antibiotic exposure when the initial population decrease exceeds four orders of magnitude from thermal shock alone. It appears likely that thermal shock and antibiotics act with strictly orthogonal mechanisms on P aeruginosa biofilms, but both contribute to a separate, general, critical decrease phenomenon.
4. Materials and Methods
4.1. Streak and Inoculum
Pseudomonas aeruginosa PAO1 (15692, American Type Culture Collection, Manassas, VA, USA) was streaked on an agar plate (Difco Nutrient Agar, Sparks, MD, USA) and incubated inverted for 24 h at 37 °C. Two colonies from the streaked plate were harvested and moved into 5 mL (30 g/L) Tryptic Soy Broth (TSB, Becton, Dickinson and Company, Franklin Lakes, NJ, USA) using a sterile inoculating loop. Inoculated TSB was incubated for 24 h at 37 °C to form an inoculum with an average of ~109 colony forming units (CFU) per mL.
4.2. Biofilm Culture
4.2.1. Shaker Table Biofilm
In total, 0.333 mL of inoculum and 5 mL of 30 g/L TSB were added to each well of 4-well dishes (Thermo Fisher Scientific, Waltham, MA, USA). In each well, a microscope slide fully frosted on one side, 75 mm × 25 mm × 1 mm (Leica Biosystems, Buffalo Grove, IL, USA) was immersed, and then the dish was sealed with parafilm. Dishes were placed on an orbital shaker table (VWR 1000, 15 mm orbit, Radnor, PA, USA) set at 160 rpm, and placed inside an incubator at 37 °C for 96 h to culture mature ST biofilms.
4.2.2. Drip Flow Reactor (DFR) Biofilm
DFR biofilm culture includes two distinct modes, batch and continuous. A frosted microscope slide was immersed in 15 mL (30 g/L) TSB in each well of a 4-well reactor (Biosurface Technologies Corporation, Bozeman, MT). In total, 1 mL of inoculum was added to each well, then the wells were sealed with their lids. During batch mode, the reactor was at rest horizontally inside a 37 °C incubator for 4 h. To begin continuous mode, the reactor was tilted by 10 degrees and a steady drip of TSB (3 g/L) was applied to the upper end of the nascent biofilm, flowing down the slide by gravity to a drain hose at the lower end. The continuous mode ran for 20 h inside the incubator at 37 °C, with a drip flowrate of 1.25 L/day per well.
4.3. Thermal Shock
Mature ST or DFR biofilms were removed from their culture wells and rinsed in 5 mL of 3 g/L TSB for 1 min to remove planktonic bacteria, and then transferred to preheated 4-well dishes with 5 mL of 3 g/L TSB in each well. Biofilms were thermally shocked at (50, 60, or 70 °C) for (1, 2, 5, 10, or 30 min). Temperature was controlled by keeping the dishes in a water bath at the target temperature for 30 min prior to thermal shock and throughout the shock itself. Biofilms were transferred immediately after the thermal shock to new 4-well dishes with 5 mL of 3 g/L TSB per well at ambient temperature.
4.4. Re-Incubation
To investigate the viability of thermally shocked biofilms, they were re-incubated under conditions identical to their initial culturing. Thermally shocked DFR biofilms were placed in a fresh DFR inclined at 10°, in which the 3 g/L TSB drip. Biofilms were re-incubated for 4 or 24 h at 37 °C and the same flowrate of 1.25 L/day per well.
4.5. Antibiotic Exposure
In total, 5 mg/mL ciprofloxacin (CP) stock was prepared by dissolving ciprofloxacin hydrochloride (MP Biomedicals, Santa Ana, CA, USA) in de-ionized water. The stock was filtered with a 0.22 μm PES membrane sterile filter (Millex®GP filter unit) and stored at 2 °C.
4.5.1. Shaker Table Antibiotic Exposure
Mature ST biofilms were rinsed in 5 mL of TSB (3 g/L) for 1 min to remove planktonic bacteria, and then placed in a fresh 4-well dish, where each well contained 5 mL TSB (30 g/L) and CP (0.25, 1, 4, 16, 64, or 256 μg/mL). These concentrations were selected to observe antibiotic effects below, at, or above intravenous administration concentrations (4 μg/mL) on biofilm [39]. Biofilms were kept in these antibiotics challenge plates for 4, 24, or 48 h at 37 °C.
4.5.2. Drip Flow Reactor Antibiotic Exposure
Mature DFR biofilms were placed in a fresh DFR inclined at 10°, in which the 3 g/L TSB drip also contained CP at 0.25, 4, or 64 μg/mL. The flowrate remained at 1.25 L/day per well for 4, 24, or 48 h at 37 °C.
4.6. Antibiotic and Thermal Exposure
To investigate the interaction of antibiotics and thermal shock on the reduction of biofilm population density, mature biofilms were exposed to antibiotics for four hours as described above. Following this exposure, biofilms were immediately transferred to preheated 4-well dishes with 5 mL of 3 g/L TSB and the same antibiotic concentration for thermal shock as described above. After the thermal shock, biofilms were either immediately enumerated, or reincubated in a new 4-well dish (ST) or new reactor (DFR) at 37 °C with the same antibiotics concentration for the remainder of a 24 or 48 h antibiotics exposure.
4.7. Enumeration
Biofilms’ population density was determined via suspension and plating. Biofilms were transferred to a fresh 4-well dish containing 5 mL of 3 g/L TSB and mechanically disrupted and homogenized with a sonicator bath for 10 min at 45kHz (VWR Symphony, 9.5 L). The sonicated homogenous solutions were then serial diluted by 10 fold, and spot plated with 10 μL samples on nutrient agar plates. After 20 h of incubation, grown colonies were counted and the population density of colony forming units per square centimeter was calculated.
4.8. Confocal Microscopy
Confocal laser scanning microscopy was performed to evaluate ST and DFR biofilms’ architectural characteristics. Biofilms were exposed to fluorescent dyes from a LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen, Eugene, OR) for 20 min in low-light conditions to stain the biofilms prior to imaging with a non-inverted confocal microscope (Zeiss LSM 710, Oberkochen, Germany).
4.9. Statistical Analysis
Enumeration results were calculated on a log scale for statistical analysis. Significance was determined using two-tailed student-t tests with a 95% confidence interval. Variance for each trial arm is assumed to be uncorrelated, and differences in variance were reconciled per Cochran [41].
5. Conclusions
Reincubation studies on treated P aeruginosa biofilms indicate a critical population decrease beyond which the population will continue decreasing rather than recover. The factor by which thermal shock or ciprofloxacin reduce biofilm population density appear to be the same regardless of whether the other treatment factor is also applied, indicating that the reduction mechanisms are orthogonal (i.e., independent of each other) rather than synergistic. However, beyond the critical overall population decrease factor, the biofilm population continues to crash to zero, resulting in a post-treatment reduction far beyond the product of the individual treatments, making them appear synergistic even if their mechanisms do not actually overlap.
Author Contributions
Conceptualization, H.A. and E.N.; Data curation, H.A.; Formal analysis, H.A. and E.N.; Funding acquisition, H.A. and E.N.; Investigation, H.A., Y.G. and H.C.; Methodology, H.A. and E.N.; Project administration, E.N.; Resources, H.A. and E.N.; Supervision, E.N.; Validation, H.A.; Visualization, H.A.; Writing—original draft, H.A. and E.N.; Writing—review and editing, E.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the American Heart Association, grant number 18IPA34170108 and the National Science Foundation, grant number CBET-1133297.
Data Availability Statement
The raw data collated and reported here is available in the NuxollResearchGroup DataVerse (URL and journal-specific dataset name to be added after manuscript acceptance).
Acknowledgments
Aljaafari, Haydar is a PhD student supported by the Higher Committee of Education Development in Iraq (HCED). The Zeiss LSM 710 confocal microscope was also obtained with support from the National Institutes of Health (1 S10 RR025439–01).
Conflicts of Interest
The authors declare no conflict of interest, and the funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- McDermott, K.W.; Freeman, W.J.; Elixhauser, A. Overview of Operating Room Procedures during Inpatient Stays in U.S. Hospitals, 2014; HCUP Statistical brief #233; Agency for Healthcare Research and Quality: Rockville, MD, USA, 2017. [Google Scholar]
- O’Toole, P.; Maltenfort, M.G.; Chen, A.F.; Parvizi, J. Projected increase in periprosthetic joint infections secondary to rise in diabetes and obesity. J. Arthroplast. 2016, 31, 7–10. [Google Scholar] [CrossRef]
- Parvizi, J.; Ghanem, E.; Azzam, K.; Davis, E.; Jaberi, F.; Hozack, W. Periprosthetic infection: Are current treatment strategies adequate? Acta Orthop. Belg. 2008, 74, 793–800. [Google Scholar]
- Phillips, J.E.; Crane, T.P.; Noy, M.; Elliott, T.S.J.; Grimer, R.J. The incidence of deep prosthetic infections in a specialist orthopaedic hospital. J. Bone Jt. Surg. Br. Vol. 2006, 88, 943–948. [Google Scholar] [CrossRef]
- Cui, Q.; Mihalko, W.M.; Shields, J.S.; Ries, M.; Saleh, K.J. Antibiotic-impregnated cement spacers for the treatment of infection associated with total hip or knee arthroplasty. J. Bone Jt. Surg. Am. Vol. 2007, 89, 871–882. [Google Scholar] [CrossRef]
- Anderl, J.N.; Zahller, J.; Roe, F.; Stewart, P. Role of nutrient limitation and stationary-phase existence in klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob. Agents Chemother. 2003, 47, 1251–1256. [Google Scholar] [CrossRef] [Green Version]
- Piddock, L.J.V. Multidrug-resistance efflux pumps? Not just for resistance. Nat. Rev. Genet. 2006, 4, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kamesh, A.C.; Xiao, Y.; Sun, V.; Hayes, M.; Daniell, H.; Koo, H. Topical delivery of low-cost protein drug candidates made in chloroplasts for biofilm disruption and uptake by oral epithelial cells. Biomaterials 2016, 105, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Pérez-Díaz, M.; Alvarado-Gomez, E.; Magaña-Aquino, M.; Sánchez-Sánchez, R.; Velasquillo, C.; Gonzalez, C.; Ganem-Rondero, A.; Martinez-Castanon, G.-A.; Zavala-Alonso, N.; Martinez-Gutierrez, F. Anti-biofilm activity of chitosan gels formulated with silver nanoparticles and their cytotoxic effect on human fibroblasts. Mater. Sci. Eng. C 2016, 60, 317–323. [Google Scholar] [CrossRef]
- Darouiche, R.O. Treatment of infections associated with surgical implants. N. Engl. J. Med. 2004, 350, 1422–1429. [Google Scholar] [CrossRef]
- Tran, N.; Tran, P. Nanomaterial-based treatments for medical device-associated infections. ChemPhysChem 2012, 13, 2481–2494. [Google Scholar] [CrossRef] [PubMed]
- Osmon, D.R.; Berbari, E.F.; Berendt, A.R.; Lew, D.; Zimmerli, W.; Steckelberg, J.M.; Rao, N.; Hanssen, A.; Wilson, W.R. Executive summary: Diagnosis and management of prosthetic joint infection: Clinical practice guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2013, 56, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berend, K.R.; Lombardi, A.V.; Morris, M.J.; Bergeson, A.G.; Adams, J.; Sneller, M.A. Two-stage treatment of hip periprosthetic joint infection is associated with a high rate of infection control but high mortality. Clin. Orthop. Relat. Res. 2013, 471, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, H.; Della Valle, C.J. The two-stage standard in revision total hip replacement. Bone Jt. J. 2013, 95-B, 84–87. [Google Scholar] [CrossRef]
- Rohde, H.; Burandt, E.C.; Siemssen, N.; Frommelt, L.; Burdelski, C.; Wurster, S.; Scherpe, S.; Davies, A.; Harris, L.; Horstkotte, M.A.; et al. Polysaccharide intercellular adhesin or protein factors in biofilm accumulation of Staphylococcus epidermidis and Staphylococcus aureus isolated from prosthetic hip and knee joint infections. Biomaterials 2007, 28, 1711–1720. [Google Scholar] [CrossRef]
- Wilkins, M.; Hall-Stoodley, L.; Allan, R.; Faust, S.N. New approaches to the treatment of biofilm-related infections. J. Infect. 2014, 69, S47–S52. [Google Scholar] [CrossRef]
- Tulloch, A.W.; Chun, Y.; Kealey, C.; Mohanchandra, K.P.; Chang, J.; Milisavljevic, V.; Levi, D.S.; Lawrence, P.F.; Rigberg, D.A. PS236. Hydrophilic surface treatment of thin film nickel titanium reduces bacterial biofilm production compared to commercially available endograft materials. J. Vasc. Surg. 2010, 51, 79S–80S. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Ravn, D.B.; Gram, L.; Kingshott, P. Stainless steel modified with poly(ethylene glycol) can prevent protein adsorption but not bacterial adhesion. Colloids Surf. B Biointerfaces 2003, 32, 275–291. [Google Scholar] [CrossRef]
- Tunney, M.; Gorman, S. Evaluation of a poly(vinyl pyrollidone)-coated biomaterial for urological use. Biomaterials 2002, 23, 4601–4608. [Google Scholar] [CrossRef]
- Francolini, I.; Donelli, G. Prevention and control of biofilm-based medical-device-related infections. FEMS Immunol. Med. Microbiol. 2010, 59, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Koopaie, M.; Bordbar-Khiabani, A.; Kolahdooz, S.; Darbandsari, A.K.; Mozafari, M. Advanced surface treatment techniques counteract biofilm-associated infections on dental implants. Mater. Res. Express 2020, 7, 015417. [Google Scholar] [CrossRef]
- Chen, J.; Howell, C.; Haller, C.A.; Patel, M.S.; Ayala, P.; Moravec, K.A.; Dai, E.; Liu, L.; Sotiri, I.; Aizenberg, M.; et al. An immobilized liquid interface prevents device associated bacterial infection in vivo. Biomaterials 2017, 113, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Khare, M.D.; Bukhari, S.S.; Swann, A.; Spiers, P.; McLaren, I.; Myers, J. Reduction of catheter-related colonisation by the use of a silver zeolite-impregnated central vascular catheter in adult critical care. J. Infect. 2007, 54, 146–150. [Google Scholar] [CrossRef]
- Tan, L.; Li, J.; Liu, X.; Cui, Z.; Yang, X.; Zhu, S.; Li, Z.; Yuan, X.; Zheng, Y.; Yeung, K.; et al. Rapid biofilm eradication on bone implants using red phosphorus and near-infrared light. Adv. Mater. 2018, 30, e1801808. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ren, D. Controlling Streptococcus mutans and Staphylococcus aureus biofilms with direct current and chlorhexidine. AMB Express 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Coffel, J.; Nuxoll, E. Magnetic nanoparticle/polymer composites for medical implant infection control. J. Mater. Chem. B 2015, 3, 7538–7545. [Google Scholar] [CrossRef] [PubMed]
- Van der Borden, A.; van der Mei, H.; Busscher, H. Electric block current induced detachment from surgical stainless steel and decreased viability of Staphylococcus epidermidis. Biomaterials 2005, 26, 6731–6735. [Google Scholar] [CrossRef]
- Carmen, J.C.; Roeder, B.L.; Nelson, J.L.; Ogilvie, R.L.R.; Robison, R.; Schaalje, G.B.; Pitt, W.G. Treatment of biofilm infections on implants with low-frequency ultrasound and antibiotics. Am. J. Infect. Control. 2005, 33, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Zhang, X.; Yang, Y.; Zhang, H.; Shi, J.; Yao, X. Near-infrared light-triggered therapy to combat bacterial biofilm infections by MoSe2 /TiO2 nanorod arrays on bone implants. Adv. Mater. Interfaces 2019, 7. [Google Scholar] [CrossRef]
- Richardson, I.P.; Sturtevant, R.; Heung, M.; Solomon, M.; Younger, J.G.; Vanepps, J.S. Hemodialysis catheter heat transfer for biofilm prevention and treatment. ASAIO J. 2016, 62, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Bandara, H.; Nguyen, D.; Mogarala, S.; Osiñski, M.; Smyth, H. Magnetic fields suppress Pseudomonas aeruginosa biofilms and enhance ciprofloxacin activity. Biofouling 2015, 31, 443–457. [Google Scholar] [CrossRef]
- O’Toole, A.; Ricker, E.B.; Nuxoll, E. Thermal mitigation of Pseudomonas aeruginosabiofilms. Biofouling 2015, 31, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Chopra, R.; Shaikh, S.; Chatzinoff, Y.; Munaweera, I.; Cheng, B.; Daly, S.M.; Xi, Y.; Bing, C.; Burns, D.; Greenberg, D.E. Employing high-frequency alternating magnetic fields for the non-invasive treatment of prosthetic joint infections. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricker, E.B.; Nuxoll, E. Synergistic effects of heat and antibiotics on Pseudomonas aeruginosa biofilms. Biofouling 2017, 33, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Hajdu, S.; Holinka, J.; Reichmann, S.; Hirschl, A.M.; Graninger, W.; Presterl, E. Increased temperature enhances the antimicrobial effects of daptomycin, vancomycin, tigecycline, fosfomycin, and cefamandole on staphylococcal biofilms. Antimicrob. Agents Chemother. 2010, 54, 4078–4084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Wang, L.; Gong, M.; Lin, Y.; Xu, Y.; Ye, L.; Yu, X.; Liu, J.; Liu, J.; He, S.; et al. Synergistic effects of nanoparticle heating and amoxicillin on H. pylori inhibition. J. Magn. Magn. Mater. 2019, 485, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Pijls, B.G.; Sanders, I.M.J.G.; Kuijper, E.J.; Nelissen, R.G.H.H. Synergy between induction heating, antibiotics, and N-acetylcysteine eradicates Staphylococcus aureus from biofilm. Int. J. Hyperth. 2020, 37, 130–136. [Google Scholar] [CrossRef] [Green Version]
- McEvoy, G.K. (Ed.) AHFS Drug Information 2008; American Society of Health-System Pharmacists: Bethesda, MD, USA, 2008. [Google Scholar]
- Ricker, E.B.; Aljaafari, H.A.S.; Bader, T.M.; Hundley, B.S.; Nuxoll, E. Thermal shock susceptibility and regrowth of Pseudomonas aeruginosabiofilms. Int. J. Hyperth. 2018, 34, 168–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochran, W.J. Approximate significance levels of the behrens-fischer test. Biometrics 1964, 20, 191–195. [Google Scholar] [CrossRef]
Figure 1.
Culture protocols produce biofilm with starkly different population density, architecture, and thermal susceptibility. (a,b) are overhead views of confocal fluorescent images of shaker table (a) and drip flow reactor (b) biofilms. (c) Effect of thermal shock on population density of P. aeruginosa ST and DFR biofilms. † from [35].
Figure 1.
Culture protocols produce biofilm with starkly different population density, architecture, and thermal susceptibility. (a,b) are overhead views of confocal fluorescent images of shaker table (a) and drip flow reactor (b) biofilms. (c) Effect of thermal shock on population density of P. aeruginosa ST and DFR biofilms. † from [35].


Figure 2.
Critical population density for DFR biofilm reincubation. (a) Population density for DFR biofilm reincubation after thermal shock at 60 °C. (b) Population density for DFR biofilm reincubation after thermal shock at 70 °C. Error bars indicate standard deviation for at least six slides from three different dishes.
Figure 2.
Critical population density for DFR biofilm reincubation. (a) Population density for DFR biofilm reincubation after thermal shock at 60 °C. (b) Population density for DFR biofilm reincubation after thermal shock at 70 °C. Error bars indicate standard deviation for at least six slides from three different dishes.

Figure 3.
Effect of Ciprofloxacin on ST and DFR biofilms. (a) Effect of Ciprofloxacin on ST biofilm. (b) Effect of Ciprofloxacin on DFR biofilm. Error bars indicate standard deviation for at least six slides from three different dishes.
Figure 3.
Effect of Ciprofloxacin on ST and DFR biofilms. (a) Effect of Ciprofloxacin on ST biofilm. (b) Effect of Ciprofloxacin on DFR biofilm. Error bars indicate standard deviation for at least six slides from three different dishes.

Figure 4.
Combined ciprofloxacin and thermal shock effect on the P. aeruginosa shaker table biofilm population. Each panel shows results for the thermal shock and antibiotic exposure indicated. Red horizontal lines show the critical population density below which thermal shocked bacterial biofilms are not viable. Error bars indicate standard deviation for at least six slides from three different dishes.
Figure 4.
Combined ciprofloxacin and thermal shock effect on the P. aeruginosa shaker table biofilm population. Each panel shows results for the thermal shock and antibiotic exposure indicated. Red horizontal lines show the critical population density below which thermal shocked bacterial biofilms are not viable. Error bars indicate standard deviation for at least six slides from three different dishes.

Figure 5.
Combined ciprofloxacin and thermal shock effect on the P. aeruginosa drip flow reactor biofilm population. Each panel shows results for the thermal shock and antibiotic exposure indicated. Red horizontal lines show the critical population density below which thermal shocked bacterial biofilms are not viable. Error bars indicate standard deviation for at least six slides from three different dishes.
Figure 5.
Combined ciprofloxacin and thermal shock effect on the P. aeruginosa drip flow reactor biofilm population. Each panel shows results for the thermal shock and antibiotic exposure indicated. Red horizontal lines show the critical population density below which thermal shocked bacterial biofilms are not viable. Error bars indicate standard deviation for at least six slides from three different dishes.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aljaafari, H.; Gu, Y.; Chicchelly, H.; Nuxoll, E. Thermal Shock and Ciprofloxacin Act Orthogonally on Pseudomonas aeruginosa Biofilms. Antibiotics 2021, 10, 1017. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081017
AMA Style
Aljaafari H, Gu Y, Chicchelly H, Nuxoll E. Thermal Shock and Ciprofloxacin Act Orthogonally on Pseudomonas aeruginosa Biofilms. Antibiotics. 2021; 10(8):1017. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081017
Chicago/Turabian StyleAljaafari, Haydar, Yuejia Gu, Hannah Chicchelly, and Eric Nuxoll. 2021. "Thermal Shock and Ciprofloxacin Act Orthogonally on Pseudomonas aeruginosa Biofilms" Antibiotics 10, no. 8: 1017. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081017
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.