
Evidence of Another Anthropic Impact on Iguana delicatissima from the Lesser Antilles: The Presence of Antibiotic Resistant Enterobacteria

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
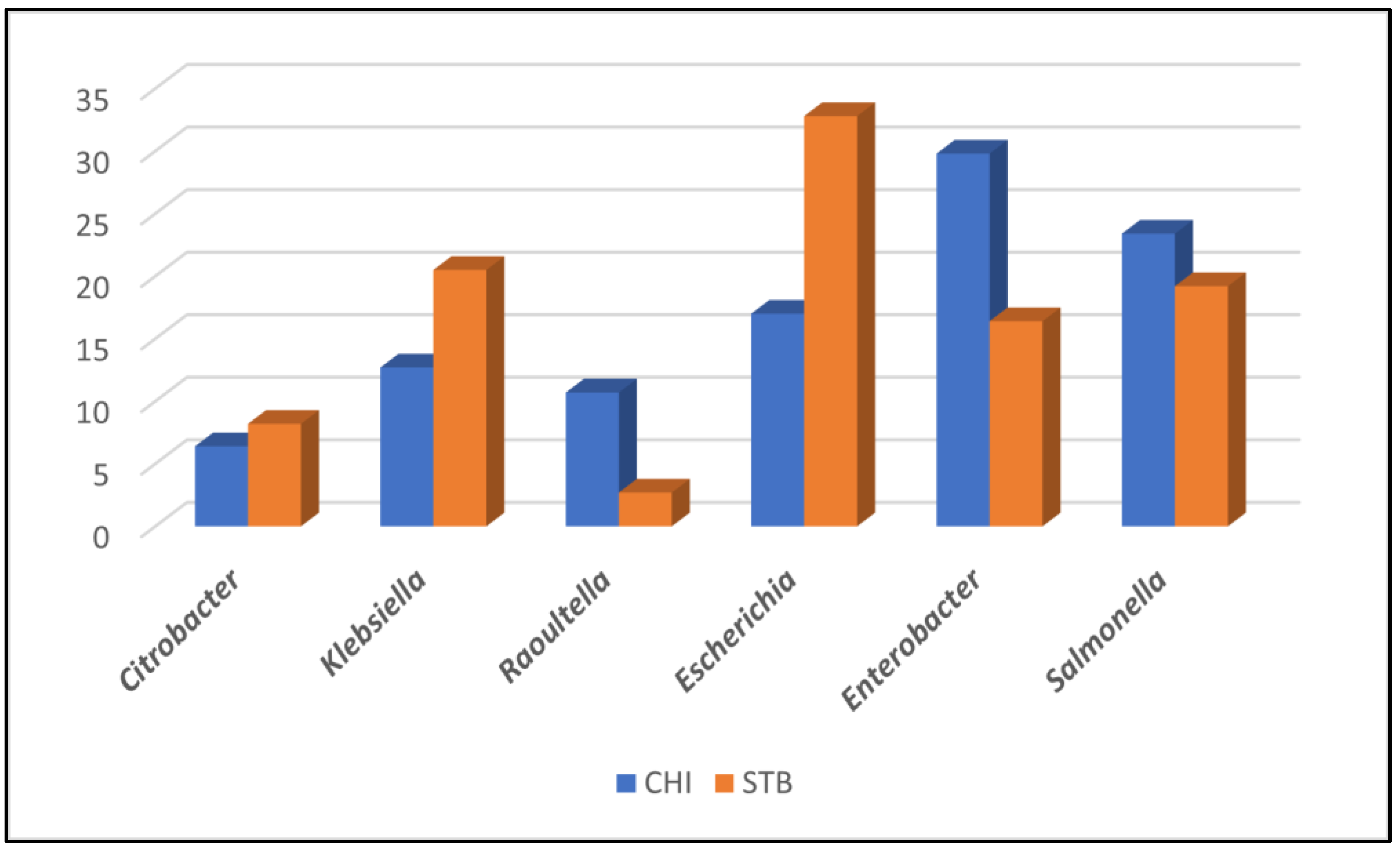
2.1. Bacterial Isolates Identification
2.2. Resistance Traits in DOM Isolates
2.3. Resistance Traits in the Rare Antibiotic-Resistant (RAR) Isolates
3. Materials and Methods
3.1. Sampling
3.2. Bacterial Isolation and Identification
3.3. Antibiotic Susceptibility Profiles
3.4. Similarity between Chancel Island and St. Barth
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethic Statement
References
- WHO (World Health Organization). Antimicrobial Resistance: Global Report on Surveillance; WHO: Geneva, Switzerland, 2014; Available online: https://apps.who.int/iris/handle/10665/112642 (accessed on 3 July 2021).
- United Nations. Political Declaration of the High-Level Meeting of the General Assembly on Antimicrobial Resistance: Draft resolution / submitted by the President of the General Assembly. In Proceedings of the Political Declaration of the High-level Meeting of the General Assembly on Antimicrobial Resistance, A/71/L.2, New York, NY, USA, 22 September 2016. [Google Scholar]
- Gaze, W.H.; Zhang, L.; Abdouslam, N.A.; Hawkey, P.M.; Calvo-Bado, L.; Royle, J.; Brown, H.; Davis, S.; Kay, P.; Boxall, A.B.A.; et al. Impacts of anthropogenic activity on the ecology of class 1 integrons and integron-associated genes in the environment. ISME J. 2011, 5, 1253–1261. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare, A.; Eckert, E.M.; Teruggi, A.; Fontaneto, D.; Bertoni, R.; Callieri, C.; Corno, G. Constitutive presence of antibiotic resistance genes within the bacterial community of a large subalpine lake. Mol. Ecol. 2015, 24, 3888–3900. [Google Scholar] [CrossRef]
- Pallecchi, L.; Bartoloni, A.; Paradisi, F.; Rossolini, G.M. Antibiotic resistance in the absence of antimicrobial use: Mechanisms and implications. Expert. Rev. Anti. Infect. Ther. 2008, 6, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Woodward, D.L.; Khakhria, R.; Johnson, W.M. Human salmonellosis associated with exotic pets. J. Clin. Microbiol. 1997, 35, 2786–2790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylvester, W.R.B.; Amadi, V.; Hegamin-Younger, C.; Pinckney, R.; Macpherson, C.N.L.; McKibben, J.S.; Bruhl-Day, R.; John-Sylvester, K.D.; Harharan, H. Occurrence of Antibiotic Resistant Escherichia Coli in Green Iguanas (Iguana Iguana) in Grenada, West Indies. Int. J. Vet. Med. Res. Rep. 2014, 2014, 260412. [Google Scholar] [CrossRef]
- Bautista-Trujillo, G.U.; Gutiérrez-Miceli, F.A.; Mandujano-García, L.; Oliva-Llaven, M.A.; Ibarra-Martínez, C.; Mendoza-Nazar, P.; Ruiz-Sesma, B.; Tejeda-Cruz, C.; Pérez-Vázquez, L.C.; Pérez-Batrez, J.E.; et al. Captive Green Iguana Carries Diarrheagenic Escherichia coli Pathotypes. Front. Vet. Sci. 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Thaller, M.C.; Migliore, L.; Marquez, C.; Tapia, W.; Cedeño, V.; Rossolini, G.M.; Gentile, G. Tracking acquired antibiotic resistance in commensal bacteria of Galápagos land iguanas: No man, no resistance. PLoS ONE 2010, 5, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, E.; Hong, P.Y.; Bedon, L.C.; Mackie, R.I. Carriage of antibiotic-resistant enteric bacteria varies among sites in Galápagos reptiles. J. Wildl. Dis. 2012, 48, 56–67. [Google Scholar] [CrossRef]
- Breuil, M. Les iguanes (Iguana delicatissima) de l îlet Chancel (Martinique). Report, Office National des Forêts-Direction Régionale de l Environnement Martinique. Association des Amis du Laboratoire des Reptiles et Amphibiens du MNHN. 1994.
- Rodrigues, C.; Laffitte, D. Population survey, Chancel Islet, Martinique: Mars 2013. Report Office National de la Chasse et de la Faune Sauvage, Cellule Technique Antilles Françaises. 2013. [Google Scholar]
- Knapp, C.; Breuil, M.; Rodrigues, C.; Iverson, J. (Eds.) Lesser Antillean Iguana, Iguana delicatissima: Conservation Action Plan, 2014—2016; IUCN SSC Iguana Specialist Group: Gland, Switzerland, 2014. [Google Scholar]
- Van den Burg, M.; Breuil, M.; Knapp, C. Iguana delicatissima. In The IUCN Red List of Threatened Species 2018: E.T10800A122936983; International Union for Conservation of Nature and Natural Resources, 2018; Available online: https://www.iucnredlist.org/species/10800/122936983 (accessed on 3 July 2021).
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef]
- Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.-H.; Stackebrandt, E. (Eds.) The Prokaryotes: Proteobacteria: Gamma Subclass; Springer: Berlin/Heidelberg, Germany, 2006; Volume 6. [Google Scholar]
- Guyomard-Rabenirina, S.; Weill, F.-X.; Le Hello, S.; Bastian, S.; Berger, F.; Ferdinand, S.; Legreneur, P.; Loraux, C.; Malpote, E.; Muanza, B.; et al. Reptiles in Guadeloupe (French West Indies) are a reservoir of major human Salmonella enterica serovars. PLoS ONE 2019, 14, e0220145. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M.; Winstanley, T.G.; Shannon, K.P. Interpretative reading: Recognizing the unusual and inferring resistance mechanisms from resistance phenotypes. J. Antimicrob. Chemother. 2001, 48, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van duijkeren, E.; Johnson, A.P.; Gaastra, W. Editorial: Assessing the antimicrobial susceptibility of bacteria obtained from animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef]
- Bourély, C.; Cazeau, G.; Jarrige, N.; Jouy, E.; Haenni, M.; Lupo, A.; Madec, J.Y.; Leblond, A.; Gay, E. Co-resistance to Amoxicillin and Tetracycline as an Indicator of Multidrug Resistance in Escherichia coli Isolates From Animals. Front. Microbiol. 2019, 10, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.M.; Du, C.; Xu, H.; Miao, Y.H.; Cheng, Y.Y.; Tang, H.; Zhou, J.H.; Wang, X.C. Occurrence of tetracycline-resistant fecal coliforms and their resistance genes in an urban river impacted by municipal wastewater treatment plant discharges. J. Environ. Sci. Heal. Part A 2015, 50, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Bryan, A.; Shapir, N.; Sadowsky, M.J. Frequency and Distribution of Tetracycline Resistance Genes in Genetically Diverse, Nonselected, and Nonclinical Escherichia coli Strains Isolated from Diverse Human and Animal Sources. Appl. Environ. Microbiol. 2004, 70, 2503–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengeløv, G.; Halling-Sørensen, B.; Aarestrup, F.M. Susceptibility of Escherichia coli and Enterococcus faecium isolated from pigs and broiler chickens to tetracycline degradation products and distribution of tetracycline resistance determinants in E. coli from food animals. Vet. Microbiol. 2003, 95, 91–101. [Google Scholar] [CrossRef]
- Guyomard-Rabenirina, S.; Reynaud, Y.; Pot, M.; Albina, E.; Couvin, D.; Ducat, C.; Gruel Gaëlle, F.S.; Legreneur, P.; Le Hello, S.; Malpote, E.; et al. Antimicrobial Resistance in Wildlife in Guadeloupe (French West Indies): Distribution of a Single blaCTX–M–1/IncI1/ST3 Plasmid Among Humans and Wild Animals. Front. Microbiol. 2020, 11, 1524. [Google Scholar] [CrossRef] [PubMed]
- Bartoloni, A.; Benedetti, M.; Pallecchi, L.; Larsson, M.; Mantella, A.; Strohmeyer, M.; Bartalesi, F.; Fernandez, C.; Guzman, E.; Vallejos, Y.; et al. Evaluation of a rapid screening method for detection of antimicrobial resistance in the commensal microbiota of the gut. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 119–125. [Google Scholar] [CrossRef]
- Herlemann, D.P.R.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stock, I.; Grueger, T.; Wiedemann, B. Natural antibiotic susceptibility of strains of Serratia marcescens and the S. liquefaciens complex: S. liquefaciens sensu stricto, S. proteamaculans and S. grimesii. Int. J. Antimicrob. Agents 2003, 22, 35–47. [Google Scholar] [CrossRef]
- Guardabassi, L.; Dijkshoorn, L.; Collard, J.M.; Olsen, J.E.; Dalsgaard, A. Distribution and in-vitro transfer of tetracycline resistance determinants in clinical and aquatic Acinetobacter strains. J. Med. Microbiol. 2000, 49, 929–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 3 July 2021).




{kind=link}
{kind=link}
{kind=link}
| Site | Categorization | Identification | n° of Isolates | AMP | AUG | CTX | FEP | S | K | CN | AK | NA | CIP | C | SXT | TE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CHI | DOM | Citrobacter (C. koseri cluster) | 1 | WT | ||||||||||||
| Citrobacter (C. freundii cluster) | 2 | WT | WT | |||||||||||||
| E. coli | 8 | 1 | ||||||||||||||
| Enterobacter | 14 | WT | WT | 1 | 1 | 1 | ||||||||||
| Klebsiella | 6 | WT | ||||||||||||||
| Raoultella | 5 | WT | ||||||||||||||
| Salmonella | 11 | 1 | ||||||||||||||
| CHI | RAR | Enterobacter | 4 | WT | WT | 1 | 1 | 1 | 2 | |||||||
| E. coli | 8 | 8 | 4 | 1 | 1 | |||||||||||
| Hafnia | 1 | WT | WT | 1 | ||||||||||||
| Proteus mirabilis | 1 | WT | 1 | WT | ||||||||||||
| Klebsiella | 1 | WT | 1 | 1 | ||||||||||||
| STB | DOM | Citrobacter (C. koseri cluster) | 2 | WT | ||||||||||||
| Citrobacter (C. freundii cluster) | 4 | WT | WT | 1 | ||||||||||||
| E. coli | 24 | 1 | ||||||||||||||
| Enterobacter | 12 | WT | WT | 1 | ||||||||||||
| Klebsiella | 15 | WT | ||||||||||||||
| Raoultella | 2 | 2 | ||||||||||||||
| Salmonella | 14 | 1 | 1 | 3 | ||||||||||||
| STB | RAR | E. coli | 19 | 13 | 4 | 9 | 8 | 7 | 5 | 4 | 2 | 3 | 7 | 13 | ||
| Enterobacter | 4 | WT | 4 | 1 | 2 | 1 | 1 | 3 | 1 | 2 | ||||||
| Klebsiella | 5 | 5 | 3 | 2 | 4 | 1 | 3 | 3 | 3 | 1 | 4 | 1 | ||||
| Raoultella | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Site | Isolates (n°) | Acquired Resistance |
|---|---|---|
| CHI | E. coli (1) | AMP/AUG |
| E. coli (4) | AMP | |
| E. coli (1) | AMP/AUG/NAL | |
| E. coli (1) | AMP/AUG/SXT | |
| Enterobacter cloacae complex. (1) | ST | |
| Enterobacter cloacae complex (1) | C/NAL | |
| Enterobacter cloacae complex (2) | SXT | |
| Hafnia alvei | SXT | |
| Klebsiella pneumoniae | NAL/TE | |
| Proteus mirabilis | NA | |
| STB | E. aerogenes (1) | ST/K/AK/NA/SXT/ |
| E. aerogenes (1) | K/CN/NA/SXT/ | |
| E. coli (1) | AMP | |
| E. coli (1) | AMP/AUG/ | |
| E. coli (1) | AMP/AUG/CTX/TE | |
| E. coli (1) | AMP/ST/K/ | |
| E. coli (1) | AMP/TE | |
| E. coli (1) | AMP/AUG/CTX/ST/K/CN/AK/C | |
| E. coli (1) | AMP/AUG/NA/SXT/ | |
| E. coli (1) | AMP/AUG/SXT/TE | |
| E. coli (1) | AMP/NA/TE | |
| E. coli (1) | AMP/SXT/TE | |
| E. coli (1) | AMP/AUG/CTX/NA/C/SXT/ | |
| E. coli (1) | AMP/AUG/ST/K/CN/C/TE | |
| E. coli (1) | AMP/ST/K/CN/NA/C/SXT/TE | |
| E. coli (2) | AMP/AUG/TE | |
| E. coli (2) | AMP/AUG/ST/K/CN/AK/TE | |
| E. coli (2) | AMP/AUG/ST/K/CN/AK/SXT/TE | |
| E. aerogenes (1) | CTX | |
| E. aerogenes (1) | CTX/NA/CIP | |
| K. oxytoca (1) | AUG/SXT/ | |
| K. oxytoca (1) | AUG/CTX/FEP/ST/K/NA/SXT/ | |
| K. oxytoca (1) | AUG/CTX/ST/K/NA/SXT/ | |
| K. oxytoca (1) | AUG/CTX/ST/K/CN/NA/C/SXT/ | |
| K. pneumoniae (1) | AUG/ST/TE | |
| R. ornythinolytica (1) | AUG/CTX/K/NA/SXT/TE |
| RAR Isolates (n°) | MDR Isolates | Ampicillin and Tetracycline Resistance | tetA/tetB |
|---|---|---|---|
| E. aerogenes (4) | 2 | NA | NA |
| E. coli (19) | 10 | 13 | 12 |
| K. oxytoca (4) | 3 | NA | NA |
| K. pneumoniae (1) | NA | NA | 0 |
| R. ornythinolytica (1) | 1 | NA | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Lallo, G.; D’Andrea, M.M.; Sennati, S.; Thaller, M.C.; Migliore, L.; Gentile, G. Evidence of Another Anthropic Impact on Iguana delicatissima from the Lesser Antilles: The Presence of Antibiotic Resistant Enterobacteria. Antibiotics 2021, 10, 885. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080885
Di Lallo G, D’Andrea MM, Sennati S, Thaller MC, Migliore L, Gentile G. Evidence of Another Anthropic Impact on Iguana delicatissima from the Lesser Antilles: The Presence of Antibiotic Resistant Enterobacteria. Antibiotics. 2021; 10(8):885. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080885
Chicago/Turabian StyleDi Lallo, Gustavo, Marco Maria D’Andrea, Samanta Sennati, Maria Cristina Thaller, Luciana Migliore, and Gabriele Gentile. 2021. "Evidence of Another Anthropic Impact on Iguana delicatissima from the Lesser Antilles: The Presence of Antibiotic Resistant Enterobacteria" Antibiotics 10, no. 8: 885. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080885