
Enzymatic Activity and Microbial Diversity of Sod-Podzolic Soil Microbiota Using 16S rRNA Amplicon Sequencing following Antibiotic Exposure

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Agrochemical Soil Indicators
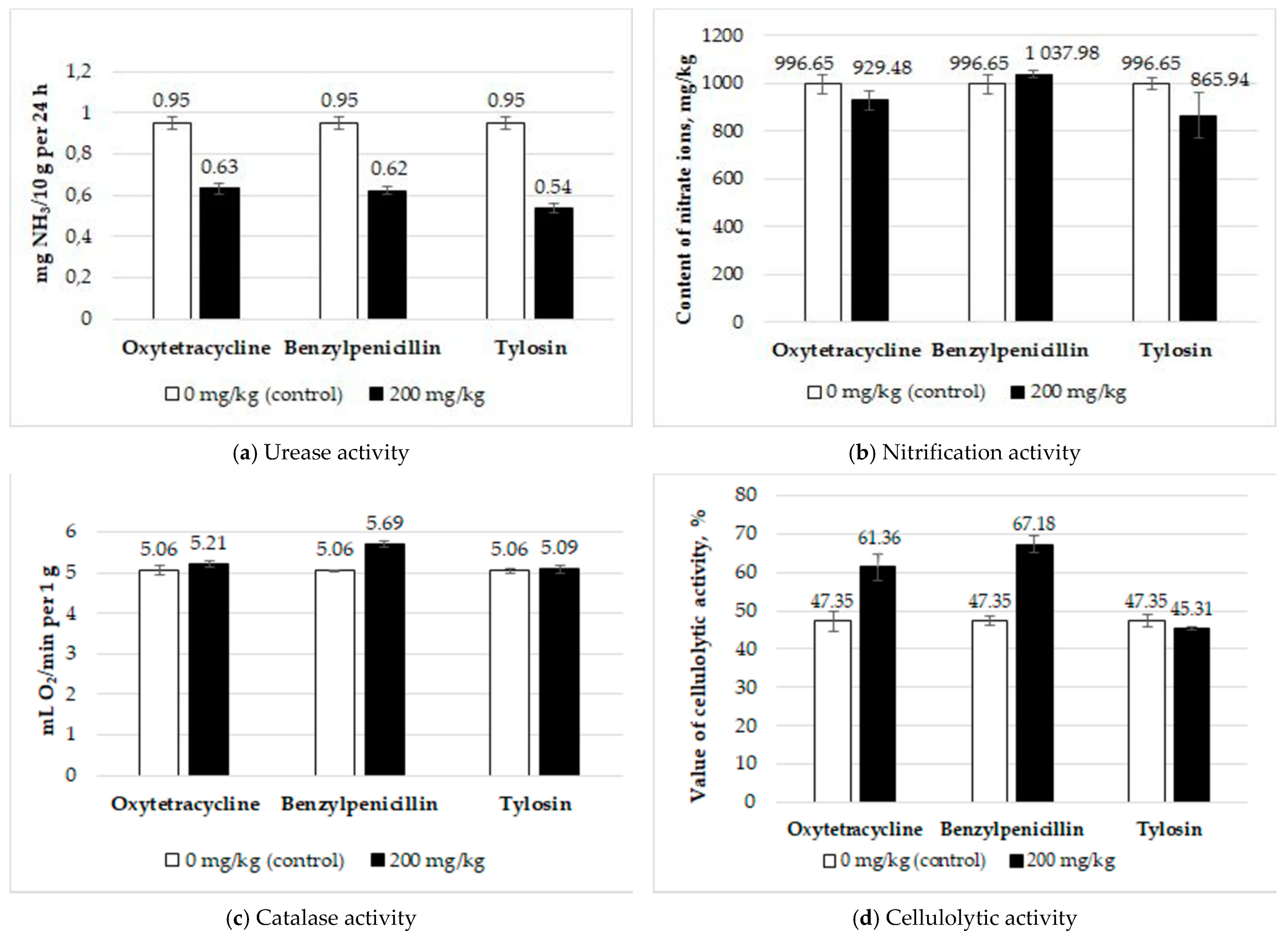
2.2. Determination of the Soil Enzymatic Activity
2.3. Metagenomic Sequencing
3. Discussion
4. Materials and Methods
4.1. Soil
4.2. Determination of the Soil Agrochemical Characteristics
4.3. Antibiotics
4.4. Enzymatic Activity Determination
4.5. Statistics Analysis
4.6. DNA Preparation for Sequencing
4.7. Sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rasschaert, G.; Van Elst, D.; Colson, L.; Herman, L.; Ferreira, H.C.D.C.; Dewulf, J.; Decrop, J.; Meirlaen, J.; Heyndrickx, M.; Daeseleire, A.E. Antibiotic Residues and Antibiotic-Resistant Bacteria in Pig Slurry Used to Fertilize Agricultural Fields. Antibiotics 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Huygens, J.; Daeseleire, E.; Mahillon, J.; Van Elst, D.; Decrop, J.; Meirlaen, J.; Dewulf, J.; Heyndrickx, M.; Rasschaert, G. Presence of Antibiotic Residues and Antibiotic Resistant Bacteria in Cattle Manure Intended for Fertilization of Agricultural Fields: A One Health Perspective. Antibiotics 2021, 10, 410. [Google Scholar] [CrossRef] [PubMed]
- Mdegela, R.H.; Mwakapeje, E.R.; Rubegwa, B.; Gebeyehu, D.T.; Niyigena, S.; Msambichaka, V.; Nonga, H.E.; Antoine-Moussiaux, N.; Fasina, F.O. Antimicrobial Use, Residues, Resistance and Governance in the Food and Agriculture Sectors, Tanzania. Antibiotics 2021, 10, 454. [Google Scholar] [CrossRef]
- Kaczala, F.; Blum, S.E. The Occurrence of Veterinary Pharmaceuticals in the Environment: A Review. Curr. Anal. Chem. 2016, 12, 169–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenni, P.; Ancona, V.; Caracciolo, A.B. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Daniel, R. The Metagenomics of Soil. Nat. Rev. Microbiol. 2005, 3, 470–478. [Google Scholar] [CrossRef]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Antibiotics in the Soil Environment—Degradation and Their Impact on Microbial Activity and Diversity. Front. Microbiol. 2019, 10, 338. [Google Scholar] [CrossRef] [PubMed]
- van de Vijver, L.; Verwer, C.; Smolders, G.; Hospers-Brands, M.; van Eekeren, N. The Cycle of Veterinary Antibiotics in the Ecosystem; Louis Bolk Instituut: Driebergen, The Netherlands, 2016; p. 31. [Google Scholar]
- Chen, W.; Liu, W.; Pan, N.; Jiao, W.; Wang, M. Oxytetracycline on functions and structure of soil microbial community. J. Soil Sci. Plant Nutr. 2013, 13, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Roose-Amsaleg, C.; Laverman, A.M. Do antibiotics have environmental side-effects? Impact of synthetic antibiotics on biogeochemical processes. Environ. Sci. Pollut. Res. 2016, 23, 4000–4012. [Google Scholar] [CrossRef] [PubMed]
- Akimenko, Y.V.; Kazeev, K.S.; Kolesnikov, S.I. Ecological Consequences of Chernozem Contamination with Antibiotics: Monograph; Southern Federal University Press: Rostov-on-Don, Russia, 2013; p. 120. ISBN 978-5-9275-1185-3. (In Russian) [Google Scholar]
- Akimenko, Y.V.; Kazeev, K.S.; Kolesnikov, S.I. The Antibiotic Contamination Influence on the Biological Properties of Ordinary Chernozem: Monograph; Southern Federal University Press: Rostov-on-Don, Russia, 2015; p. 154. ISBN 978-5-9275-1797-8. (In Russian) [Google Scholar]
- Akimenko, Y.V.; Chuvaraeva, O.V.; Kolesnikov, S.I.; Kazeev, K.S.; Minnikova, T.V. Assessment of the Ecological State of the Basic Soils of the Southern Russia under the Contamination Conditions with Antibiotics: Monograph; Southern Federal University Press: Rostov-on-Don, Russia, 2019; p. 114. ISBN 978-5-9275-3247-6. (In Russian) [Google Scholar]
- Schmitt, H. The Effects of Veterinary Antibiotics on Soil Microbial Communities. Ph.D. Thesis, Universiteit Utrecht, Utrecht, The Netherlands, 2005; p. 195. [Google Scholar]
- Wang, X.; Pan, H.; Gu, J.; Qian, X.; Gao, H.; Qin, Q. Effects of oxytetracycline on archaeal community, and tetracycline resistance genes in the anaerobic co-digestion of pig manure and wheat straw. Environ. Technol. 2016, 37, 3177–3185. [Google Scholar] [CrossRef] [PubMed]
- Westergaard, K.; Müller, A.K.; Christensen, S.; Bloem, J.; Sørensen, S.J. Effects of tylosin as a disturbance on the soil microbial community. Soil Biol. Biochem. 2001, 33, 2061–2071. [Google Scholar] [CrossRef]
- Trifonova, T.A.; Chesnokova, S.M.; Kosmacheva, A.G. Evaluation of ampicillin and tylosin antibiotics effect on the enzymatic activity of sod-podzolic soil and their toxicity for cultivated plants. Theor. Appl. Ecol. 2020, 2, 150–156. (In Russian) [Google Scholar] [CrossRef]
- Trifonova, T.A.; Chesnokova, S.M.; Kosmacheva, A.G. Impact study of antibiotics of various groups on the cellulolytic activity of sod-podzolic soil by laboratory modeling. Agrochemistry 2020, 10, 72–78. (In Russian) [Google Scholar] [CrossRef]
- Kosmacheva, A.G. Assessment of the impact of the antibiotics benzylpenicillin, oxytetracycline and tylosin on catalase activity of weakly loamy sod-podzolic and gray forest soils. Grani Pozn. 2019, 6, 79–83. (In Russian) [Google Scholar]
- Kosmacheva, A.G.; Chesnokova, S.M.; Trifonova, T.A. Study of the Effect of Antibiotics of Various Groups on the Nitrification Activity of Sod-podzolic Soil by the Method of Laboratory Modelling. Izv. Saratov Univ. Series Chem. Biol. Ecol. 2020, 3, 352–358. [Google Scholar] [CrossRef]
- Chessa, L.; Jechalke, S.; Ding, G.-C.; Pusino, A.; Mangia, N.P.; Smalla, K. The presence of tetracycline in cow manure changes the impact of repeated manure application on soil bacterial communities. Biol. Fertil. Soils 2016, 52, 1121–1134. [Google Scholar] [CrossRef]
- Reichel, R.; Rosendahl, I.; Peeters, E.T.H.M.; Focks, A.; Groeneweg, J.; Bierl, R.; Schlichting, A.; Amelung, W.; Thiele-Bruhn, S. Effects of slurry from sulfadiazine- (SDZ) and difloxacin- (DIF) medicated pigs on the structural diversity of microorganisms in bulk and rhizosphere soil. Soil Biol. Biochem. 2013, 62, 82–91. [Google Scholar] [CrossRef]
- Reichel, R.; Michelini, L.; Ghisi, R.; Thiele-Bruhn, S. Soil bacterial community response to sulfadiazine in the soil–root zone. J. Plant Nutr. Soil Sci. 2015, 178, 499–506. [Google Scholar] [CrossRef]
- Soil Sci Inst., Moscow. Unified State Register of Soil Resources of Russia. Version 1.0. 2014. (In Russian). Available online: http://egrpr.esoil.ru/ (accessed on 2 May 2021).
- Yang, Y.; Herbold, C.W.; Jung, M.-Y.; Qin, W.; Cai, M.; Du, H.; Lin, J.-G.; Li, X.; Li, M.; Gu, J.-D. Survival strategies of ammonia-oxidizing archaea (AOA) in a full-scale WWTP treating mixed landfill leachate containing copper ions and operating at low-intensity of aeration. Water Res. 2021, 191, 116798. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, Y.; Yu, H. Arthrobacter cupressi sp. nov., an actinomycete isolated from the rhizosphere soil of Cupressus sempervirens. Int. J. Syst. Evol. Microbiol. 2012, 62, 2731–2736. [Google Scholar] [CrossRef]
- Umarov, M.M.; Kurakov, A.V.; Stepanov, A.L. Microbial Transformation of Nitrogen in Soil; GEOS: Moscow, Russia, 2007; p. 138. ISBN 5-89118-315-7. (In Russian) [Google Scholar]
- Anderson, B.; de Peyster, A.; Gad, S.C.; Hakkinen, P.J.B.; Kamrin, M.; Locey, B.; Mehendale, H.M.; Pope, C.; Shugart, L.P.; Wexler, P. Encyclopedia of Toxicology, Four-Volume Set: Encyclopedia of Toxicology, 2nd ed.; Academic Press: Cambridge, MA, USA, 2005; Volume 1, p. 794. [Google Scholar]
- Riviere, J.; Lees, P.; Elliott, J.; Clarke, C.; Anadón, A.; Baggot, D.; Brown, S.A.; Burka, J.F.; Clarke, C.R.; Craigmill, A.; et al. USP Veterinary Pharmaceutical Information Monographs—Antibiotics. J. Vet. Pharmacol. Ther. 2003, 26, 1–271. [Google Scholar] [CrossRef] [PubMed]
- Voznyakovskaya, Y.M. (Ed.) Principle Microbiological and Biochemical Methods of Soil Research: Method, Recommendations; VNIISKHM: Leningrad, Russia, 1987; p. 47. (In Russian) [Google Scholar]
- Khaziev, F.K. Methods of Soil Enzymology; Nauka: Moscow, Russia, 2005; p. 252. ISBN 5-02-033940-7. (In Russian) [Google Scholar]
- Kazeev, K.S.; Kolesnikov, S.I.; Valkov, V.F. Biological Diagnostic and Indication of Soils: The Methodology and Methods of Research; Publishing House of Rostov University: Rostov-on-Don, Russia, 2003; p. 216. (In Russian) [Google Scholar]
- Bates, S.T.; Berg-Lyons, D.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the global distribution of dominant archaeal populations in soil. ISME J. 2011, 5, 908–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator, Unit | Value |
|---|---|
| Acidity, pHKCl | 5.56 ± 0.20 |
| P2O5 mobile, mg/kg | 436.5 ± 87.30 |
| K2O mobile, mg/kg | 275.6 ± 41.34 |
| Organic matter, % | 2.36 ± 0.47 |
| N ammonium, mg/kg | 0 |
| N nitrates, mg/kg | 10.1 ± 1.52 |
| Physical clay, % | 30.2 |
| Sample | OTU | Chao1 Index | Shannon Index | Simpson Index | Feit Phylogenetic Diversity |
|---|---|---|---|---|---|
| 1 | 186 | 189.8 | 4.74 | 0.90 | 7.91 |
| 3 | 191 | 199 | 5.21 | 0.95 | 7.77 |
| 4 | 194 | 202 | 4.90 | 0.92 | 8.01 |
| 5 | 197 | 201.9 | 4.49 | 0.88 | 7.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trifonova, T.; Kosmacheva, A.; Sprygin, A.; Chesnokova, S.; Byadovskaya, O. Enzymatic Activity and Microbial Diversity of Sod-Podzolic Soil Microbiota Using 16S rRNA Amplicon Sequencing following Antibiotic Exposure. Antibiotics 2021, 10, 970. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080970
Trifonova T, Kosmacheva A, Sprygin A, Chesnokova S, Byadovskaya O. Enzymatic Activity and Microbial Diversity of Sod-Podzolic Soil Microbiota Using 16S rRNA Amplicon Sequencing following Antibiotic Exposure. Antibiotics. 2021; 10(8):970. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080970
Chicago/Turabian StyleTrifonova, Tatiana, Anastasia Kosmacheva, Alexander Sprygin, Svetlana Chesnokova, and Olga Byadovskaya. 2021. "Enzymatic Activity and Microbial Diversity of Sod-Podzolic Soil Microbiota Using 16S rRNA Amplicon Sequencing following Antibiotic Exposure" Antibiotics 10, no. 8: 970. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080970